
What Is in Store for EPS Microalgae in the Next Decade?

 ,
,  ,
, 


Abstract
:1. Introduction
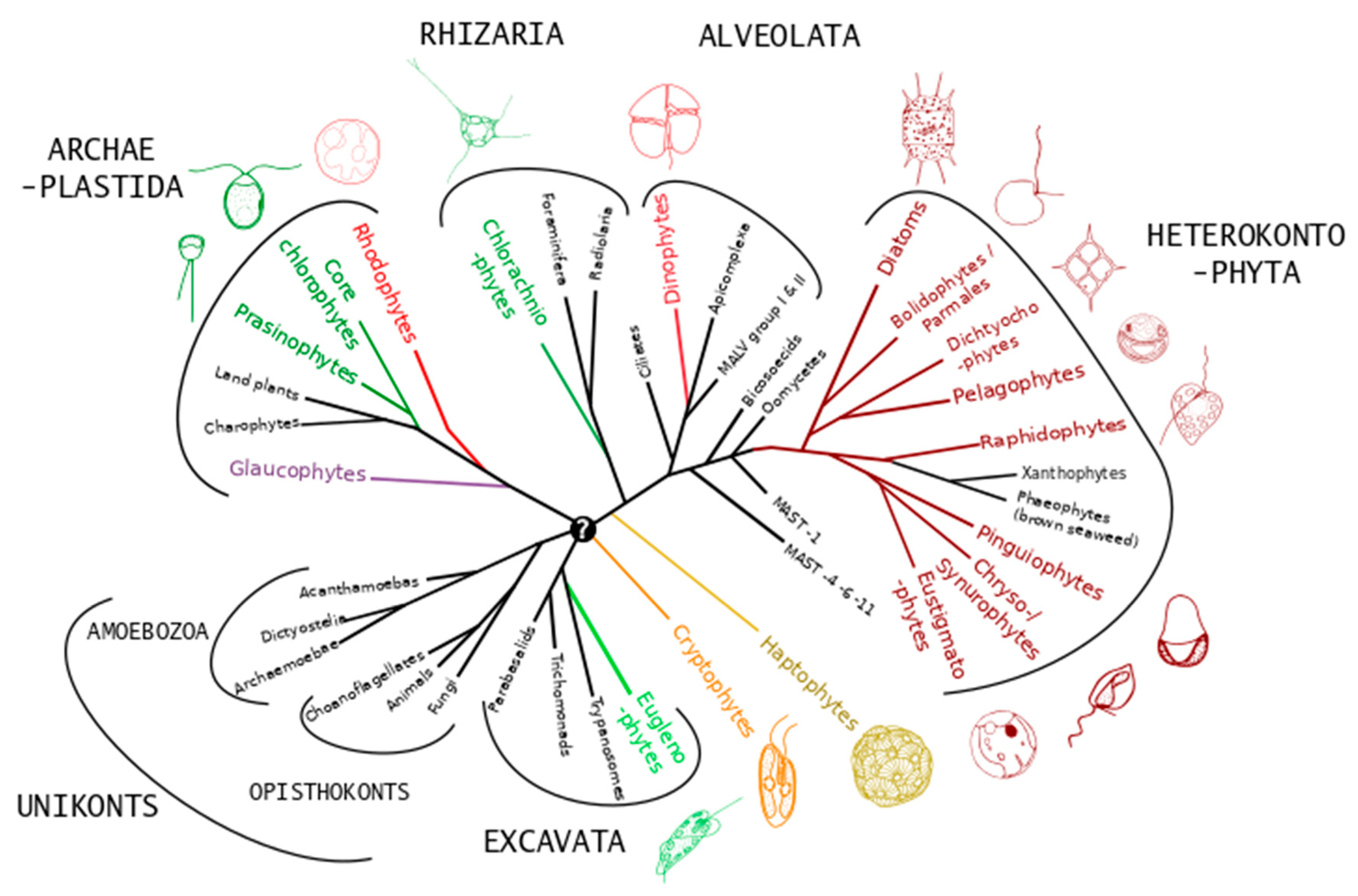
2. Ecology of Microalgae and Diversity
2.1. Morphological Diversity
2.2. Ecological Diversity
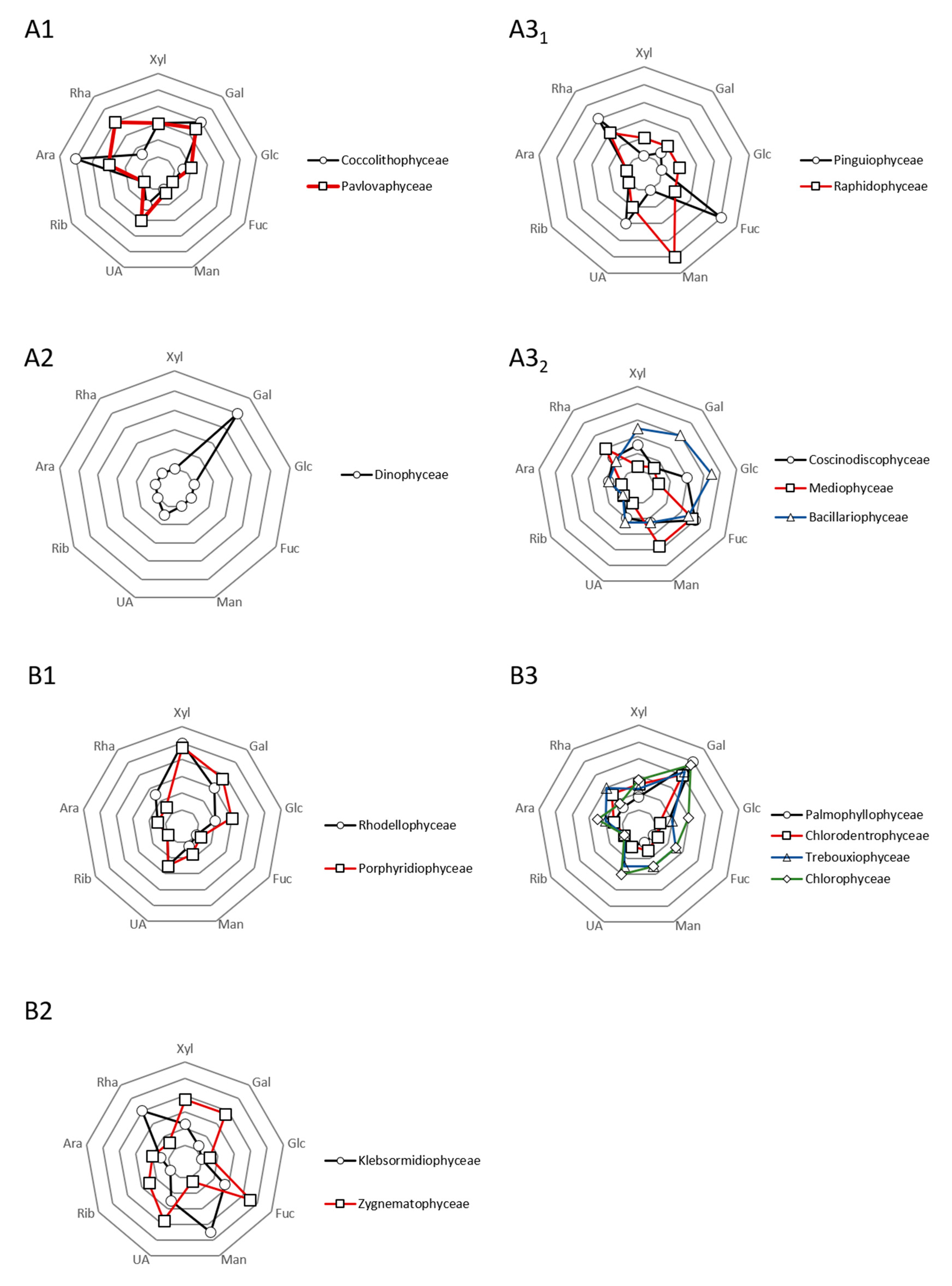
2.3. Biochemical Diversity
3. Biomass and Exopolysaccharides from Microalgae
3.1. Production of Biomass
3.2. Exopolysaccharides (EPS) from Microalgae
4. Current Valorization and Markets for EPS Microalgae
4.1. Food and Nutraceutical Applications
4.2. Cosmetic Applications
4.3. Pharmaceutical Applications
5. Perspectives on Issues Affecting the Future of EPS Microalgae
5.1. Differences Between Bacterial and Microalgal EPS Production
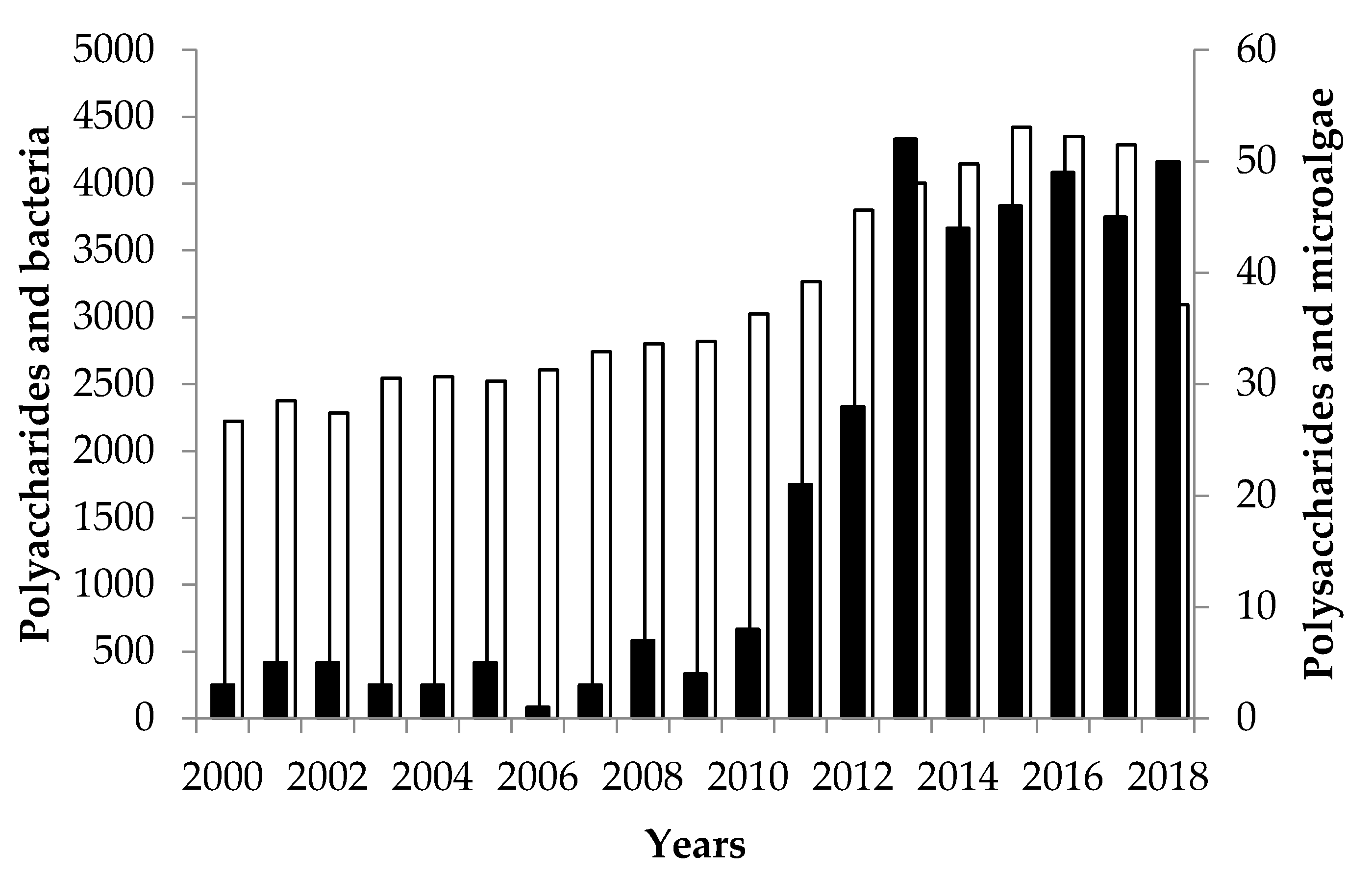
5.1.1. EPS Productivity and EPS Concentration
5.1.2. Production Systems
5.1.3. Culture Medium
5.1.4. Downstream Processing
5.1.5. Factors Limiting the Development of Microalgal EPS
5.2. Suggestions to Reduce Microalgae EPS Production Cost
5.2.1. Strain Selections
5.2.2. EPS Production and Concentration Improvement
5.2.3. Culture Costs Reduction
5.2.4. Biorefinery Approach
5.2.5. Omic Approach
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bazaka, K.; Crawford, R.J.; Nazarenko, E.L.; Ivanova, E.P. Bacterial extracellular polysaccharides. Adv. Exp. Med. Biol. 2011, 715, 213–226. [Google Scholar] [CrossRef]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysacharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef]
- Nvodo, U.U.; Green, E.; Okoh, A.I. Bacterial Exopolysaccharides: Functionality and Prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A. Handbook of Microalgal Culture: Biotechnology and Applied Phycology, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2004; p. 588. ISBN 978-0-632-05953-9. [Google Scholar]
- Norton, T.A.; Melkonian, M.; Andersen, R.A. Algal biodiversity. Phycologia 1996, 35, 308–326. [Google Scholar] [CrossRef]
- De Philippis, R.; Sili, C.; Paperi, R.; Vincenzini, M. Exopolysaccharide-producing cyanobacteria and their possible exploitation: A review. J. Appl. Phycol. 2001, 13, 293–299. [Google Scholar] [CrossRef]
- Gaignard, C.; Gargouch, N.; Dubessay, P.; Delattre, C.; Pierre, G.; Laroche, C.; Fendri, I.; Abdelkafi, S.; Michaud, P. New Horizons in Culture and Valorization of Red Microalgae. Biotechnol. Adv. 2019, 37, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Yeh, K.L.; Aisyah, R.; Lee, D.J.; Chang, J.S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef]
- Johnson, T.J.; Katuwal, S.; Anderson, G.A.; Gu, L.; Zhou, R.; Gibbons, W.R. Photobioreactor cultivation strategies for microalgae and cyanobacteria. Biotechnol. Prog. 2018, 34, 811–827. [Google Scholar] [CrossRef]
- Perez-Garcia, O.; Bashan, Y. Microalgal heterotrophic and mixotrophic culturing for Bio-refining: From matabolic routes to techno-economics. In Algal Biorefineries, 1st ed.; Prokop, A., Bajpai, R., Zappi, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 61–131. [Google Scholar] [CrossRef]
- Pignolet, O.; Jubeau, S.; Vacca-Garcia, C.; Michaud, P. Highly valuable microalgae: Biochemical and topological aspects. J. Ind. Microbiol. Biotechnol. 2013, 40, 781–796. [Google Scholar] [CrossRef]
- Whitfield, C.; Valvano, M.A. Biosynthesis and expression of cell-surface polysaccharide in gram negative bacteria. Adv. Microb. Physiol. 1993, 35, 136–246. [Google Scholar] [CrossRef]
- Kamerling, J.P.; Gerwig, G.J. Strategies for the structural analysis of carbohydrates. In Comprehensive Glycoscience—From Chemistry to Systems Biology, 1st ed.; Kamerling, J.P., Boons, G.-J., Lee, Y.C., Suzuki, A., Taniguchi, N., Voragen, A.G.J., Eds.; Elsevier, B.V.: Oxford, UK, 2007; Volume 2, pp. 1–68. [Google Scholar] [CrossRef]
- Arad, S.M.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotech. 2010, 21, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Prieto, A.; Weber, A.P.M.; Bhattacharya, D. The origin and establishment of the plastid in algae and plants. Annu. Rev. Genet. 2007, 41, 147–168. [Google Scholar] [CrossRef] [PubMed]
- Not, F.; Siano, R.; Kooistra, W.; Simon, N.; Vaulot, D.; Probert, I. Chapter one—Diversity and ecology of eukaryotic marine phytoplankton. Adv. Bot. Res. 2012, 64, 1–53. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: Cell size and elemental stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef]
- Becker, B.; Marin, B.; Melkonian, M. Structure, composition, and biogenesis of prasinophyte cell coverings. Protoplasma 1994, 181, 233–244. [Google Scholar] [CrossRef]
- Becker, B.; Becker, D.; Kamerling, J.P.; Melkonian, M. 2-Keto-sugar acids in green flagellates: A chemical marker for prasinophycean scales. J. Phycol. 1991, 27, 498–504. [Google Scholar] [CrossRef]
- Blokker, P.; Schouten, S.; van den Ende, H.; de Leeuw, J.W.; Hatcher, P.G.; Sinninghe Damsté, J.S. Chemical structure of algaenan from the fresh water Tetraedron minimum, Scenedesmus communis and Pediastrum boryanum. Organ. Geochem. 1998, 29, 1453–1468. [Google Scholar] [CrossRef]
- Cordeiro, L.M.; Carbonero, E.R.; Sassaki, G.L.; Reis, R.A.; Stocker-Worgotter, E.; Gorin, P.A.; Iacomini, M. A fungus-type beta-galactofuranan in the cultivated Trebouxia photobiont of the lichen Ramalina gracilis. Fems Microbiol. Lett. 2005, 244, 193–198. [Google Scholar] [CrossRef]
- Adair, W.S.; Appel, H. Identification of a highly conserved hydroxyproline-rich glycoprotein in the cell walls of Chlamydomonas reinhardtii and two other Volvocales. Planta 1989, 179, 381–386. [Google Scholar] [CrossRef]
- Leadbeater, B.S.C. Silicification of “cell walls” of certain protistan flagellates. Phil. Trans. R. Soc. Lond. B. 1984, 304, 529–536. [Google Scholar] [CrossRef]
- Ichinomiya, M.; Lopes dos Santos, A.; Gourvil, P.; Yoshikawa, S.; Kamiya, M.; Ohki, K.; Audic, S.; de Vargas, C.; Noël, M.-H.; Vaulot, D.; et al. Diversity and oceanic distribution of the Parmales (Bolidophyceae) a picoplanktonic group closely related to diatoms. ISME J. 2016, 10, 2419–2434. [Google Scholar] [CrossRef] [PubMed]
- Brett, S.J.; Perasso, L.; Wetherbee, R. Structure and development of the cryptomonad periplast: A review. Protoplasma 1994, 181, 106–122. [Google Scholar] [CrossRef]
- Williamson, C.J.; Cameron, K.A.; Cook, J.M.; Zarsky, J.D.; Stibal, M.; Edwards, A. Glacier algae: A dark past and a darker future. Front. Microbiol. 2019, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cardon, Z.G.; Gray, D.W.; Lewis, L.A. The green algal underground: Evolutionary secrets of desert cells. Bioscience 2008, 58, 114–122. [Google Scholar] [CrossRef]
- Tesson, S.V.M.; Ambelas Skjoth, C.; Santi-Temkiv, T.; Londahl, J. Airborne microalgae: Insights, opportunities, and challenges. Appl. Environ. Microbiol. 2016, 82, 1978–1991. [Google Scholar] [CrossRef]
- MacIntyre, H.L.; Geider, R.J.; Miller, D.C. Microphytobenthos: The ecological role of the “secret garden” of unvegetated, shallow-water marine habitats. I. Distribution, abundance and primary production. Estuaries 1996, 19, 186–201. [Google Scholar] [CrossRef]
- Pinckney, J.L. A mini-review of the contribution of benthic microalgae to the ecology of the continental shelf in the South Atlantic Bight. Estaur. Coast. 2018, 41, 2070–2078. [Google Scholar] [CrossRef]
- Not, F.; Latasa, M.; Scharek, R.; Viprey, M.; Karleskind, P.; Balagué, V.; Ontoria-Oviedo, I.; Cumino, A.; Goetze, E.; Vaulot, D.; et al. Phytoplankton diversity across the Indian Ocean: A focus on the picoplanktonic size fraction. Deep Sea Res. Pt. I 2008, 55, 1456–1473. [Google Scholar] [CrossRef] [Green Version]
- Burkholder, J.M.; Glibert, P.M.; Skelton, H.M. Mixotrophy, a major mode of nutrition for harmful algal species in eutrophic waters. Harmful Algae 2008, 8, 77–93. [Google Scholar] [CrossRef]
- Ball, S.; Colleoni, C.; Cenci, U.; Raj, J.N.; Tirtiaux, C. The evolution of glycogen and starch metabolism in eukaryotes gives molecular clues to understand the establishment of plastid endosymbiosis. J. Exp. Bot. 2011, 62, 1775–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. Central and storage carbon metabolism of the brown alga Ectocarpus siliculosus: Insights into the origin and evolution of storage carbohydrates in Eukaryotes. New Phytol. 2010, 188, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfield, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant. J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Mourelle, M.; Gómez, C.; Legido, J.J.C. The potential use of marine microalgae and cyanobacteria in cosmetics and thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Ejike, C.E.; Collins, S.A.; Balasuriya, N.; Swanson, A.K.; Mason, B.; Udenigwe, C.C. Prospects of microalgae proteins in producing peptide-based functional foods for promoting cardiovascular health. Trends Food Sci. Tech. 2017, 59, 30–36. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Ras, M.; Steyer, J.P.; Bernard, O. Temperature effect on microalgae: A crucial factor for outdoor production. Rev. Environ. Sci. Biotechnol. 2013, 12, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Villay, A.; Laroche, C.; Roriz, D.; El-Alaoui, H.; Delbac, F.; Michaud, P. Optimisation of culture parameters for exopolysaccharides production by the microalga Rhodella violacea. Bioresour. Technol. 2013, 146, 732–735. [Google Scholar] [CrossRef]
- Soanen, N.; Silva, E.D.; Gardarin, C.; Michaud, P.; Laroche, C. Improvement of exopolysaccharide production by Porphyridium marinum. Bioresour. Technol. 2016, 213, 231–238. [Google Scholar] [CrossRef]
- Gardian, Z.; Bumber, L.; Schrofel, A.; Herbstova, M.; Nebesarova, J.; Vacha, F. Organisation of photosystem I and photosystem II in red alga Cyanidium caldarium: Encounter of cyanobacterial and higher plant concept. Biochim. Biophys. Acta 2007, 1767, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, T.; Aoki, M.; Jun, X.; Uedan, T.; Nakamura, Y.; Fujiwara, S.; Umemura, T.; Tsuzuki, M.; Minoda, A. Profiling of lipid and glycogen accumulations under different growth conditions in the sulfothermophilic red alga Galdieria sulphuraria. Bioresour. Technol. 2016, 200, 861–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaoka, A.; Ozawa, Y.; Ueno, Y.; Endo, T.; Morimoto, Y.; Urushiyama, A.; Ohmori, D.; Imai, T. Cyanidioschyzon merolae ferredoxin: A high-resolution crystal structure analysis and its thermal stability. Febs Lett. 2011, 585, 1299–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.K.; Tan, H.M. Effect of temperature, light intensity and dilution rate on the cellular composition red alga Porphyridium cruentum in light-limited chemostat cultures. MIRCEN J. Appl. Microbiol. Biotechnol. 1988, 4, 231–237. [Google Scholar] [CrossRef]
- Nicolaus, B.; Panico, A.; Lama, L.; Romano, I.; Manca, M.C.; De Giulio, A.; Gambacorta, A. Chemical composition and production of exopolysaccharides from representative members of heterocystous and non-heterocystous cyanobacteria. Phytochemistry 1999, 52, 639–647. [Google Scholar] [CrossRef]
- Otero, A.; Vincenzini, M. Nostoc (Cyanophyceae) goes nude: Extracellular polysaccharides serve as a sink for reducing power under unbalanced C/N metabolism. J. Phycol. 2004, 40, 74–81. [Google Scholar] [CrossRef]
- Lupi, F.M.; Fernandes, H.M.L.; Si-Correia, I.; Novais, J.M. Temperature profiles of cellular growth and exopolysaccharide synthesis by Botryococus braunii Kfitz. UC 58. J. Appl. Phycol. 1991, 3, 35–42. [Google Scholar] [CrossRef]
- Moreno, J.; Vargas, M.A.; Olivares, H.; Rivas, J.; Guerrero, M.G. Exopolysaccharide production by the cyanobacterium Anabaena sp. ATCC 33047 in batch and continuous culture. J. Biotechnol. 1998, 60, 175–182. [Google Scholar] [CrossRef]
- Iqbal, M.; Zafar, S.I. Effects of Photon Flux Density, CO2, Aeration Rate, and Inoculum Density on Growth and Extracellular Polysaccharide Production by Porphyridium cruentum. Folia Microbiol. 1993, 38, 509–514. [Google Scholar] [CrossRef]
- Trabelsi, L.; Ouada, H.B.; Bacha, H.; Ghoul, M. Combined effect of temperature and light intensity on growth and extracellular polymeric substance production by the cyanobacterium Arthrospira platensis. J. Appl. Phycol. 2009, 21, 405–412. [Google Scholar] [CrossRef]
- de Philippis, R.; Sili, C.; Vincenzini, M. Response of an exopolysaccharide-producing heterocystous cyanobacterium to changes in metabolic carbon flux. J. Appl. Phycol. 1996, 8, 275–281. [Google Scholar] [CrossRef]
- Merchuk, J.C.; Ronen, M.; Giris, S.; Arad, S.M. Light/Dark Cycles in the Growth of the Red Microalga Porphyridium sp. Biotechnol. Bioeng. 1998, 59, 705–713. [Google Scholar] [CrossRef]
- Philips, E.J.; Zeman, C.; Hansen, P. Growth, photosynthesis, nitrogen fixation and carbohydrate production by a unicellular cyanobacterium, Synechococcus sp. (Cyanophyta). J. Appl. Phycol. 1989, 1, 137–145. [Google Scholar] [CrossRef]
- You, T.; Barnett, S.M. Effect of light quality on production of extracellular polysaccharides and growth rate of Porphyridium cruentum. Biochem. Eng. J. 2004, 19, 251–258. [Google Scholar] [CrossRef]
- Su, C.; Zhenming, C.; Weidong, L. Optimization of medium and cultivation conditions for enhanced exopolysaccharide yield by marine Cyanothece sp. 113. Chin. J. Oceanol. Limnol. 2007, 25, 411–417. [Google Scholar] [CrossRef]
- Barsanti, L.; Gualtieri, P. Is exploitation of microalgae economically and energetically sustainable? Algal Res. 2018, 31, 107–115. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Won, B.Y.; Cho, T.O. Two new encrusting species from the genus Phymatolithon (Hapalidiales, Corallinophycidae, Rhodophyta) from Korea. Phycologia 2019, 58, 1–13. [Google Scholar] [CrossRef]
- Rossi, F.; De Philippis, R. Exocellular Polysaccharides in Microalgae and Cyanobacteria: Chemical Features, Role and Enzymes and Genes Involved in Their Biosynthesis. In The Physiology of Microalgae, 1st ed.; Borowitzka, M., Beardall, J., Raven, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 6, pp. 565–590. [Google Scholar] [CrossRef]
- Karemore, A.; Sen, R. Downstream processing of microalgal feedstock for lipid and carbohydrate in a biorefinery concept: A holistic approach for biofuel applications. RSC Adv. 2016, 6, 29486–29496. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- de Jesus Raposo, M.F.; De Morais, A.M.B.; De Morais, R.M.S.C. Marine Polysaccharides from Algae with Potential Biomedical Applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef]
- Prybylski, N.; Toucheteau, C.; El Alaoui, H.; Bridiau, N.; Maugard, T.; Abdelkafi, S.; Fendri, I.; Delattre, C.; Dubessay, P.; Pierre, G.; et al. Bioactive polysaccharides from microalgae. In Handbook of Microalgae-based Processes and Products, 1st ed.; Jacob-Lopes, E., Ed.; Elsevier-Academic Press: Cambridge, MA, USA, 2019; in press. [Google Scholar]
- Gaignard, C.; Laroche, C.; Pierre, G.; Dubessay, P.; Delattre, C.; Gardarin, C.; Gourvil, P.; Probert, I.; Dubuffet, A.; Michaud, P. Screening of microalgae: Investigation of new exopolysaccharide producers. Algal Res. 2019, in press. [Google Scholar] [CrossRef]
- Robert, S.S.; Petrie, J.R.; Zhou, X.R.; Mansour, M.P.; Blackburn, S.I.; Green, A.G.; Singh, S.P.; Nichols, P.D. Isolation and characterization of a Δ5-fatty acid elongase from the marine microalga Pavlova salina. Mar. Biotechnol. 2009, 11, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Pohnert, G.; Wei, D. Extracellular Metabolites from Industrial Microalgae and Their Biotechnological Potential. Mar. Drugs 2016, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Leyland, B.; Leu, S.; Boussiba, S. Are Thraustochytrids algae? Fungal Biol. 2017, 121, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Lommerse, J.P.M.; Melkonian, M.; Kamerling, J.P.; Vliegenthart, J.F.G. The structure of an acidic trisaccharide component from a cell wall polysaccharide preparation of the green alga Tetraselmis striata Butcher. Carbohydr. Res. 1995, 267, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.; Melkonian, M.; Kamerling, J.P. The cell wall (theca) of Tetraselmis striata (chlorophyta): Macromolecular composition and structural elements of the complex polysaccharides. J. Phycol. 1998, 34, 779–787. [Google Scholar] [CrossRef]
- Chen, B.; Li, F.; Liu, N.; Ge, F.; Xiao, H.; Yang, Y. Role of extracellular polymeric substances from Chlorella vulgaris in the removal of ammonium and orthophosphate under the stress of cadmium. Bioresour. Technol. 2015, 190, 299–306. [Google Scholar] [CrossRef]
- Hayashi, K.; Hayashi, T.; Kojima, I. A Natural Sulfated Polysaccharide, Calcium Spirulan, Isolated from Spirulina platensis: In Vitro and ex Vivo Evaluation of Anti-Herpes Simplex Virus and Anti-Human Immunodeficiency Virus Activities. Aids Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-B.; Hayashi, T.; Hayashi, K.; Sankawa, U.; Maeda, M.; Nemoto, T.; Nakanishi, H. Further Purification and Structural Analysis of Calcium Spirulan from Spirulina platensis. J. Nat. Prod. 1998, 61, 1101–1104. [Google Scholar] [CrossRef]
- Arad, S.; Adda, M.; Cohen, E. The potential of production of sulfated polysaccharides from Porphyridium. Plant Soil 1985, 89, 117–127. [Google Scholar] [CrossRef]
- Thepenier, C.; Gudin, C. Studies on optimal conditions for polysaccharide production by Porphyridium cruentum. J. Appl. Microbiol. Biotechnol. 1985, 1, 257–268. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, S.; Wang, C.; Shi, L. Effects of Renewal Regime on Consumption of Nitrogen and Phosphorus, Biomass and Polysaccharide Production by Porphyridium cruentum in Semicontinuous Culture. In Proceedings of the 3rd International Conference on Bioinformatics and Biomedical Engineering, Beijing, China, 11–13 June 2009; No. 10777697. pp. 1–5. [Google Scholar] [CrossRef]
- Razaghi, A.; Godhe, A.; Albers, E. Effects of nitrogen on growth and carbohydrate formation in Porphyridium cruentum. Cent. Eur. J. Biol. 2014, 9, 156. [Google Scholar] [CrossRef]
- Geresh, S.; Mamontov, A.; Weinstein, J. Sulfation of extracellular polysaccharides of red microalgae: Preparation, characterization and properties. J. Biochem. Biophys. Methods 2002, 50, 179–187. [Google Scholar] [CrossRef]
- de Jesus Raposo, M.F.; de Morais, R.M.S.C.; de Morais, A.M.M.B. Bioactivity and Applications of Sulphated Polysaccharides from Marine Microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Influence of sulphate on the composition and antibacterial and antiviral properties of the exopolysaccharide from Porphyridium cruentum. Life Sci. 2014, 101, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium Spirulan, an Inhibitor of Enveloped Virus Replication, from a Blue-Green Alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef]
- Talyshinsky, M.M.; Souprun, Y.Y.; Huleihel, M.M. Anti-viral activity of red microalgal polysaccharides against retroviruses. Cancer Cell Int. 2002, 7. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-B.; Hou, X.; Hayashi, K.; Hayashi, T. Effect of partial desulfation and oversulfation of sodium spirulan on the potency of anti-herpetic activities. Carbohydr. Polym. 2007, 69, 651–658. [Google Scholar] [CrossRef]
- Mader, J.; Gallo, A.; Schommartz, T.; Handke, W.; Nagel, C.-H.; Günther, P.; Brune, W.; Reich, K. Calcium spirulan derived from Spirulina platensis inhibits herpes simplex virus 1 attachment to human keratinocytes and protects against herpes labialis. J. Allergy Clin. Immunol. 2016, 137, 197–203.e3. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, L.; Zhou, Y. Immunomodulation and antitumor activities of different-molecular-weight polysaccharides from Porphyridium cruentum. Carbohydr. Polym. 2012, 87, 1206–1210. [Google Scholar] [CrossRef]
- Tannin-Spitz, T.; Bergman, M.; van-Moppes, D.; Grossman, S.; Arad, S. Antioxidant activity of the polysaccharide of the red microalga Porphyridium sp. J. Appl. Phycol. 2005, 17, 215–222. [Google Scholar] [CrossRef]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Andhare, P.; Chauhan, K.; Dave, M.; Pathak, H. Microbial Exopolysaccharides: Advances in Applications and Future Prospects. In Biotechnology Volume 3: Microbial Biotechnology, 1st ed.; Tewari, R., Ed.; Studium Press LLC: Delhi, India, 2014; Volume 3, p. 25. [Google Scholar] [CrossRef]
- Paniagua-Michel, J.d.J.; Olmos-Soto, J.; Morales-Guerrero, E.R. Chapter Eleven—Algal and Microbial Exopolysaccharides. Adv. Food Nutr. Res. 2014, 73, 221–257. [Google Scholar] [CrossRef]
- Singha, T.K. Microbial Extracellular Polymeric Substances: Production, Isolation and Applications. Iosr J. Pharm. 2012, 2, 276–281. [Google Scholar] [CrossRef]
- Poli, A.; Di Donata, P.; Abbamondi, G.R.; Nicolaus, B. Synthesis, Production, and Biotechnological Applications of Exopolysaccharides and Polyhydroxyalkanoates by Archaea. Archaea 2011, 2011, 13. [Google Scholar] [CrossRef] [Green Version]
- Markou, G.; Nerantzis, E. Microalgae for high-value compounds and biofuels production: A review with focus on cultivation under stress conditions. Biotechnol. Adv. 2013, 31, 1532–1542. [Google Scholar] [CrossRef]
- Kim, J.-H.; Yoo, S.-J.; Oh, D.-K.; Kweon, Y.-G.; Park, D.-W.; Lee, C.-H.; Gil, G.-H. Selection of a Streptococcus equi mutant and optimization of culture conditions for the production of high molecular weight hyaluronic acid. Enzyme Microb. Technol. 1996, 19, 440–445. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.; Li, J.; Du, G.; Chen, J. Microbial production of hyaluronic acid: Current state, challenges, and perspectives. Microb. Cell Fact. 2011, 10, 99. [Google Scholar] [CrossRef] [Green Version]
- Borges, C.D.; de Paula, R.C.M.; Feitosa, J.P.A.; Vendruscolo, C.T. The influence of thermal treatment and operational conditions on xanthan produced by X. arboricola pv pruni strain 106. Carbohydr. Polym. 2009, 75, 262–268. [Google Scholar] [CrossRef]
- García-Ochoa, F.; Santos, V.E.; Casas, J.-A.; Gómez, E. Xanthan gum: Production, recovery, and properties. Biotechnol. Adv. 2000, 18, 549–579. [Google Scholar] [CrossRef]
- Krishna Leela, J.; Sharma, G. Studies on xanthan production from Xanthomonas Campestris. Bioprocess Engin. 2000, 23, 687–689. [Google Scholar] [CrossRef]
- Rottava, I.; Batesini, G.; Silva, M.F.; Lerin, L.; de Oliveira, D.; Padilha, F.F.; Toniazzo, G.; Mossi, A.; Cansian, R.L.; Di Luccio, M.; et al. Xanthan gum production and rheological behavior using different strains of Xanthomonas sp. Carbohydr. Polym. 2009, 77, 65–71. [Google Scholar] [CrossRef]
- Du, R.; Xing, H.; Jiang, H.; Zhou, Z.; Han, Y. Optimization, purification and structural characterization of a dextran produced by L. mesenteroides isolated from Chinese sauerkraut. Carbohydr. Polym. 2017, 174, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.; Teixeira, J.; Rodrigues, A.R. Production of dextransucrase, dextran and fructose from sucrose using Leuconostoc mesenteroides NRRL B512(f). Biochem. Eng. J. 2000, 4, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Doble, M.; Kruthiventi, K.; Gaikar, V.G. Biotransformations and bioprocesses. J. Chem. Technol. Biotechnol. 2005, 80, 1330. [Google Scholar] [CrossRef]
- Rehm, H.J.; Reed, G. Biotechnology: Bioprocessing, Volume 3, 2nd ed.; VCH Wiley-Blackwell: Hoboken, NJ, USA, 1993; p. 816. [Google Scholar] [CrossRef]
- Vinci, V.A.; Parekh, S.R. Handbook of Industrial Cell Culture—Mammalian, Microbial, and Plant Cells, 1st ed.; Humana Press: Towata, Japan, 2003; p. 536. [Google Scholar] [CrossRef]
- Harada, Y.; Sakata, K.; Sato, S.; Takayama, S. Chapter 1—Fermentation Pilot Plant. In Fermentation and Biochemical Engineering Handbook, 3rd ed.; Vogel, H.C., Todaro, C.M., Eds.; William Andrew Publishing: Boston, MA, USA, 2014; pp. 3–15. [Google Scholar] [CrossRef]
- Khan, M.; Karmakar, R.; Das, B.; Diba, F.; Razu, M.H. Heterotrophic Growth of Micro Algae. In Recent Advances in Microalgal Biotechnology, 1st ed.; Liu, J., Sun, Z., Gerken, H., Eds.; OMICS Group eBooks: Foster City, CA, USA, 2016; pp. 1–18. [Google Scholar]
- Pires, J.C.M. Chapter 5—Mass Production of Microalgae. In Handbook of Marine Microalgae, 1st ed.; Kim, S.-K., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 55–68. [Google Scholar] [CrossRef]
- Vuppaladadiyam, A.K.; Prinsen, P.; Raheem, A.; Luque, R.; Zhao, M. Sustainability Analysis of Microalgae Production Systems: A Review on Resource with Unexploited High-Value Reserves. Environ. Sci. Technol. 2018, 52, 14031–14049. [Google Scholar] [CrossRef] [PubMed]
- Pruvost, J.; Cornet, J.-F.; Le Borgne, F.; Jenck, J. Production industrielle de microalgues et cyanobactéries. Tech. De L’ingénieur 2011, 1–22. [Google Scholar]
- Shen, Y.; Yuan, W.; Pei, Z.J.; Wu, Q.; Mao, E. Microalgae Mass Production Methods. Trans. Asabe 2009, 52, 1275–1287. [Google Scholar] [CrossRef]
- Van Wagenen, J.; De Francisci, D.; Angelidaki, I. Comparison of mixotrophic to cyclic autotrophic/heterotrophic growth strategies to optimize productivity of Chlorella Sorokiniana. J. Appl. Phycol. 2015, 27, 1775–1782. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.C.; Todaro, C.L. Fermentation and Biochemical Engineering Handbook—Principles, Process Design and Equipment, 3rd ed.; William Andrew Publishing: Boston, MA, USA, 2014; p. 454. [Google Scholar] [CrossRef]
- Barker, P.E.; Alsop, R.M.; Vlachogiannis, G.J. Fractionation, purification and concentration of dextran solutions by ultrafiltration. J. Membr. Sci. 1984, 20, 79–91. [Google Scholar] [CrossRef]
- Palaniraj, A.; Jayaraman, V. Production, recovery and applications of xanthan gum by Xanthomonas campestris. J. Food Eng. 2011, 106, 1–12. [Google Scholar] [CrossRef]
- Molina Grima, E.; Belarbi, E.H.; Acién-Fernández, F.G.; Robles Medina, A.; Chisti, Y. Recovery of microalgal biomass and metabolites: Process options and economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef]
- Uduman, N.; Qi, Y.; Danquah, M.K.; Forde, G.M.; Hoadley, A. Dewatering of microalgal cultures: A major bottleneck to algae-based fuels. J. Renew. Sustain. Energy 2010, 2, 012701. [Google Scholar] [CrossRef]
- Pruvost, J.; Le Borgne, F.; Artu, A.; Legrand, J. Development of a thin-film solar photobioreactor with high biomass volumetric productivity (AlgoFilm©) based on process intensification principles. Algal Res. 2017, 21, 120–137. [Google Scholar] [CrossRef]
- Souliès, A.; Pruvost, J.; Castelain, C.; Burghelea, T. Microscopic flows of suspensions of the green non-motile Chlorella micro-alga at various volume fractions: Applications to intensified photobioreactors. J. Nonnewton. Fluid Mech. 2016, 231, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Dayananda, C.; Sarada, R.; Rani, M.U.; Shamala, T.R.; Ravishankar, G.A. Autotrophic cultivation of Botryococcus braunii for the production of hydrocarbons and exopolysaccharides in various media. Biomass Bioenergy 2007, 31, 87–93. [Google Scholar] [CrossRef]
- Pruvost, J.; Jaouen, P.; Marchal, L.; Jubeau, S.; Fleurence, J. Degumming method. U.S. 2017/0233499A1, 17 August 2017. [Google Scholar]
- Vanthoor-Koopmans, M.; Wijffrls, R.H.; Barbosa, M.J.; Eppink, M.H.M. Biorefinery of microalgae for food and fuel. Bioresour. Technol. 2013, 135, 142–149. [Google Scholar] [CrossRef]
- Yen, H.-W.; Hu, I.C.; Chen, C.Y.; Ho, S.H.; Lee, D.J.; Chang, J.S. Microalgae-based biorefinery—From biofuels to natural products. Bioresour. Technol. 2013, 135, 166–174. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae | Year of Approval |
|---|---|
| Arthrospira platensis (spirulina) | Used prior to May 1997 |
| Chlorella luteoviridis | Used prior to May 1997 |
| Chlorella pyrenoidosa | Used prior to May 1997 |
| Chlorella vulgaris | Used prior to May 1997 |
| Aphanizomenon flos-aquae | Used prior to May 1997 |
| Odontella aurita | 2005 |
| Ulkenia sp. | 2009 |
| Tetraselmis chuii | 2014 |
| Haematococcus pluvialis | 2014 |
| Schizochytrium sp. | 2015 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pierre, G.; Delattre, C.; Dubessay, P.; Jubeau, S.; Vialleix, C.; Cadoret, J.-P.; Probert, I.; Michaud, P. What Is in Store for EPS Microalgae in the Next Decade? Molecules 2019, 24, 4296. https://doi.org/10.3390/molecules24234296
Pierre G, Delattre C, Dubessay P, Jubeau S, Vialleix C, Cadoret J-P, Probert I, Michaud P. What Is in Store for EPS Microalgae in the Next Decade? Molecules. 2019; 24(23):4296. https://doi.org/10.3390/molecules24234296
Chicago/Turabian StylePierre, Guillaume, Cédric Delattre, Pascal Dubessay, Sébastien Jubeau, Carole Vialleix, Jean-Paul Cadoret, Ian Probert, and Philippe Michaud. 2019. "What Is in Store for EPS Microalgae in the Next Decade?" Molecules 24, no. 23: 4296. https://doi.org/10.3390/molecules24234296
APA StylePierre, G., Delattre, C., Dubessay, P., Jubeau, S., Vialleix, C., Cadoret, J. -P., Probert, I., & Michaud, P. (2019). What Is in Store for EPS Microalgae in the Next Decade? Molecules, 24(23), 4296. https://doi.org/10.3390/molecules24234296