
Volatile Compounds in Fruit Peels as Novel Biomarkers for the Identification of Four Citrus Species




Abstract
:1. Introduction
2. Results
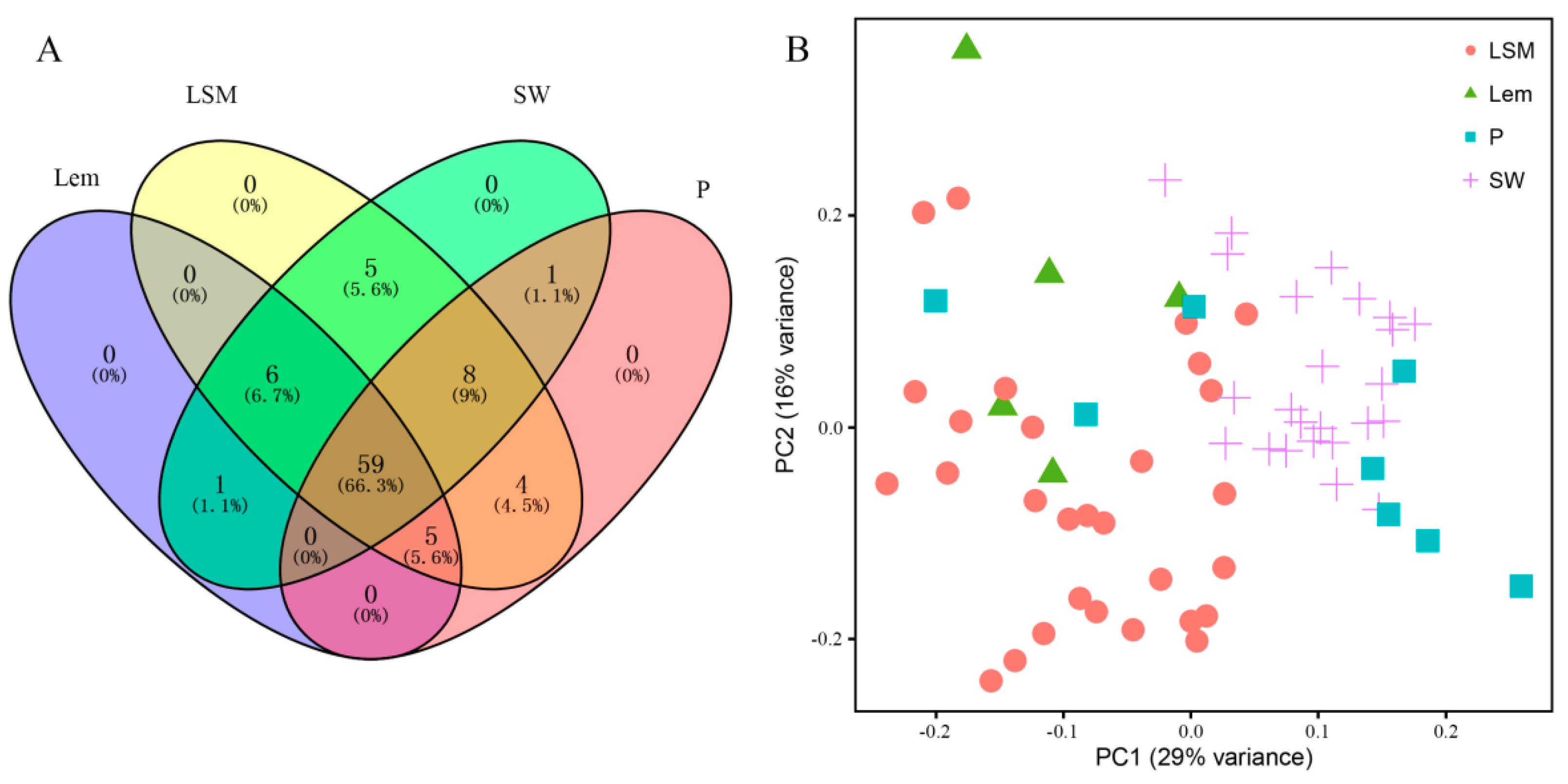
2.1. Volatile Compounds in Fruit Peels
2.2. Accumulation Pattern of Volatile Compounds is Citrus Species Dependent
2.2.1. Discrimination of LSM from the Other Three Citrus Species
2.2.2. Discrimination of Sweet Orange from the Other Three Citrus Species
2.2.3. Discrimination of Pomelo from the Other Three Citrus Species
2.2.4. Discrimination of Lemon from the Other Three Citrus Species
2.2.5. Discrimination of Clementine Mandarin from LSM and SW
2.3. Four Biomarkers for the Identification of Four Citrus Species
2.4. Discrimination of Wild and Cultivar Germplasms
3. Discussion
3.1. Biomarkers for Discriminating between Different Citrus Germplasms
3.2. Biomarkers May Be Responsible for the Citrus Species-Specific Odor Notes
3.3. Protection and Utilization of Wild Citrus Germplasms
4. Materials and Methods
4.1. Materials
4.2. Extraction and Determination of Volatiles
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, Q.; Chen, L.L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.B.; Hao, B.H.; Lyon, M.P. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, H.; Pang, Y.; Cheng, Y.; Deng, X.; Xu, J. Comparative study of flavonoid production in lycopene-accumulated and blonde-flesh sweet oranges (Citrus sinensis) during fruit development. Food Chem. 2015, 184, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, X.; Tang, Z.; Yuan, Y.; Xu, Y.; He, J.; Jiang, X.; Peng, S.-A.; Li, L.; Butelli, E. Subfunctionalization of the Ruby2–Ruby1 gene cluster during the domestication of citrus. Nat. plants 2018, 4, 930. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yan, F.; Gao, H.; He, M.; Wang, Z.; Cheng, Y.; Deng, X.; Xu, J. Features of citrus terpenoid production as revealed by carotenoid, limonoid and aroma profiles of two pummelos (Citrus maxima) with different flesh color. J. Sci. Food Agr. 2015, 95, 111–119. [Google Scholar] [CrossRef]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant. Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- Wang, S.; Tu, H.; Wan, J.; Chen, W.; Liu, X.; Luo, J.; Xu, J.; Zhang, H. Spatio-temporal distribution and natural variation of metabolites in citrus fruits. Food Chem. 2016, 199, 8–17. [Google Scholar] [CrossRef]
- Xu, J.; Ma, L.; Jiang, D.; Zhu, S.; Yan, F.; Xie, Y.; Xie, Z.; Guo, W.; Deng, X. Content evaluation of 4 furanocoumarin monomers in various citrus germplasms. Food Chem. 2015, 187, 75–81. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, Y.; Liu, C.; Chen, S.; Hu, S.; Xie, Z.; Deng, X.; Xu, J. Comprehensive comparative analysis of volatile compounds in citrus fruits of different species. Food Chem. 2017, 230, 316–326. [Google Scholar] [CrossRef]
- Baldwin, I.T. Plant volatiles. Curr. Biol. 2010, 20, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Huffaker, A.; Köllner, T.G.; Weckwerth, P.; Robert, C.A.M.; Spencer, J.L.; Lipka, A.E.; Schmelz, E.A. Selinene Volatiles Are Essential Precursors for Maize Defense Promoting Fungal Pathogen Resistance. Plant. Physiol. 2017, 175, 1455–1468. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of plant volatiles. Plant. Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.; San Andres, V.; Cervera, M.; Redondo, A.; Alquezar, B.; Shimada, T.; Gadea, J.; Rodrigo, M.J.; Zacarias, L.; Palou, L.; et al. Terpene down-regulation in orange reveals the role of fruit aromas in mediating interactions with insect herbivores and pathogens. Plant. Physiol. 2011, 156, 793–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Liu, C.; He, M.; Li, J.; Cai, Y.; Ma, Y.; Xu, J. Largely different contents of terpenoids in beef red-flesh tangerine and its wild type. BMC Plant. Biol. 2017, 17, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Liu, C.; Yao, J.-L.; Deng, C.H.; Chen, S.; Chen, J.; Wang, Z.; Yu, Q.; Cheng, Y.; Xu, J. Citrus mangshanensis Pollen Confers a Xenia Effect on Linalool Oxide Accumulation in Pummelo Fruit by Enhancing the Expression of a Cytochrome P450 78A7 Gene CitLO1. J. Agr. Food Chem. 2019, 67, 9468–9476. [Google Scholar] [CrossRef]
- Duan, L.; Guo, L.; Dou, L.-L.; Zhou, C.-L.; Xu, F.-G.; Zheng, G.-D.; Li, P.; Liu, E.-H. Discrimination of Citrus reticulata Blanco and Citrus reticulata ‘Chachi’ by gas chromatograph-mass spectrometry based metabolomics approach. Food Chem. 2016, 212, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolosi, E.; Deng, Z.N.; Gentile, A.; Malfa, S.L.; Continella, G.; Tribulato, E. Citrus phylogeny and genetic origin of important species as investigated by molecular markers. Theor. Appl. Genet. 2000, 100, 1155–1166. [Google Scholar] [CrossRef]
- Samaan, L.G. Studies on the origin of Clementine tangerine (Citrus reticulata Blanco). Euphytica 1982, 31, 167–173. [Google Scholar] [CrossRef]
- Choi, H.S. Volatile constituents of satsuma mandarins growing in Korea. Flavour Frag. J. 2004, 19, 406–412. [Google Scholar] [CrossRef]
- Lan-Phi, N.T.; Shimamura, T.; Ukeda, H.; Sawamura, M. Chemical and aroma profiles of yuzu (Citrus junos) peel oils of different cultivars. Food Chem. 2009, 115, 1042–1047. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Cheng, Y.; Zhang, H.; Deng, X.; Chen, F.; Xu, J. Volatile constituents of wild citrus Mangshanyegan (Citrus nobilis Lauriro) peel oil. J. Agr. Food Chem. 2012, 60, 2617–2628. [Google Scholar] [CrossRef] [PubMed]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Jacquemond, C.; Casanova, J. Volatile components of peel and leaf oils of lemon and lime species. J. Agr. Food Chem. 2002, 50, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Sharon-Asa, L.; Shalit, M.A.; Bar, E.; Holland, D.; Or, E.; Lavi, U. Citrus fruit flavor and aroma biosynthesis: Isolation, functional characterization, and developmental regulation of Cstps1, a key gene in the production of the sesquiterpene aroma compound valencene. Plant. J. 2003, 36, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Lei, Z.; Zhang, G.; Pilon, A.C.; Huhman, D.V.; Xie, R.; Xi, W.; Zhou, Z.; Sumner, L.W. Metabolite profiles of essential oils in citrus peels and their taxonomic implications. Metabolomics 2015, 11, 952–963. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, D.; Cheng, Y.; Deng, X.; Chen, F.; Fang, L.; Ma, Z.; Xu, J. Chemotaxonomic study of Citrus, Poncirus and Fortunella genotypes based on peel oil volatile compounds-deciphering the genetic origin of Mangshanyegan (Citrus nobilis Lauriro). PLoS ONE 2013, 8, e58411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Mas, M.C.; Rambla, J.L.; Alamar, M.C.; Gutierrez, A.; Granell, A. Comparative analysis of the volatile fraction of fruit juice from different Citrus species. PLoS ONE 2011, 6, e22016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 12. [Google Scholar] [CrossRef]
- Shen, S.L.; Yin, X.R.; Zhang, B.; Xie, X.L.; Jiang, Q.; Grierson, D.; Chen, K.S. CitAP2.10 activation of the terpene synthase CsTPS1 is associated with the synthesis of (+)-valencene in ‘Newhall’ orange. J. Exp. Bot. 2016, 67, 4105–4115. [Google Scholar] [CrossRef] [Green Version]
- Schnee, C.; Kollner, T.G.; Held, M.; Turlings, T.C.; Gershenzon, J.; Degenhardt, J. The products of a single maize sesquiterpene synthase form a volatile defense signal that attracts natural enemies of maize herbivores. Proc. Natl. Acad. Sci. USA 2006, 103, 1129–1134. [Google Scholar] [CrossRef] [Green Version]
- Oliveros, J.C. An Interactive Tool for Comparing Lists with Venn Diagrams. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 12 December 2019).
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics’ feature selection and multiple data integration. PLoS Comput Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds not are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Compounds | VIP | P-Value | ID | Compounds | VIP | P-Value |
|---|---|---|---|---|---|---|---|
| Loose-skin mandarin (LSM) | Lemon (Lem) | ||||||
| C44 | β-elemene | 1.98 | 3.85 × 10−10 | C89 | limettin | 2.51 | 8.00 × 10−16 |
| C38 | valencene | 1.75 | 4.67 × 10−6 | C22 | citronellal | 2.40 | 2.91 × 10−4 |
| C36 | (Z)-β-farnesene | 1.63 | 7.15 × 10−6 | C47 | trans-α-bergamotene | 2.30 | 1.26 × 10−13 |
| C61 | caryophyllene oxide | 1.63 | 2.76 × 10−4 | C54 | β-bisabolene | 2.22 | 1.26 × 10−13 |
| C56 | germacrene B | 1.60 | 5.95 × 10−7 | C45 | cis-α-bergamotene | 1.96 | 5.52 × 10−9 |
| C73 | 3-hexenal | 1.56 | 3.69 × 10−2 | C3 | β-pinene | 1.95 | 1.56 × 10−3 |
| C46 | γ-elemene | 1.54 | 9.47 × 10−5 | C35 | caryophyllene | 1.89 | 9.80 × 10−4 |
| C37 | α-caryophyllene | 1.51 | 7.68 × 10−10 | C13 | camphene | 1.81 | 2.00 × 10−3 |
| C40 | δ-elemene | 1.50 | 1.86 × 10−7 | C41 | α-cubebene | 1.71 | 1.15 × 10−4 |
| C10 | γ-terpinene | 1.50 | 8.75 × 10−4 | C18 | α-terpineol | 1.57 | 1.03 × 10−3 |
| Sweet orange (SW) | C36 | (Z)-β-farnesene | 1.56 | 4.24 × 10−3 | |||
| C38 | valencene | 2.54 | 6.19 × 10−9 | Pomelo (P) | |||
| C61 | caryophyllene oxide | 2.44 | 2.90 × 10−11 | C88 | nootkanone | 3.07 | 1.13 × 10−5 |
| C10 | γ-terpinene | 1.61 | 2.16 × 10−8 | C73 | 3-hexenal | 2.01 | 1.51 × 10−3 |
| C5 | α-phellandrene | 1.56 | 5.68 × 10−5 | C30 | trans-limonene oxide | 1.87 | 2.70 × 10−3 |
| C12 | α-thujene | 1.53 | 2.61 × 10−8 | C43 | β-cubebene | 1.77 | 2.07 × 10−3 |
| C30 | trans-limonene oxide | 1.53 | 9.49 × 10−4 | C58 | elemol | 1.77 | 5.16 × 10−4 |
| C49 | germacrene D | 1.52 | 1.38 × 10−7 | C42 | copaene | 1.61 | 4.16 × 10−3 |
| C6 | α-terpinene | 1.50 | 2.05 × 10−7 | ||||
| Citrus Germplasms | Actual | β-Elemene a | Valencene b | Nootkanone c | Limettin d |
|---|---|---|---|---|---|
| Fu tangerine | LSM | Yes (Y) | N | N | N |
| Huapi tangerine | LSM | Y | N | N | N |
| Zhuhong tangerine | LSM | Y | N | N | N |
| Nian tangerine | LSM | Y | N | N | N |
| Shatang tangerine | LSM | Y | N | N | N |
| Suhong tangerine | LSM | No (N) | N | N | N |
| Nanfengmi tangerine | LSM | Y | N | N | N |
| Huanongbendizao tangerine | LSM | Y | N | N | N |
| Tu tangerine | LSM | Y | N | N | N |
| Red tangerine | LSM | Y | N | N | N |
| Anliu sweet orange | SW | N | Y | N | N |
| Valencia sweet orange | SW | N | Y | N | N |
| Hamlin sweet orange | SW | Y | Y | N | N |
| Hong anliu sweet orange | SW | N | Y | N | N |
| Cara cara navel orange | SW | N | Y | N | N |
| Jincheng sweet orange | SW | N | Y | N | N |
| Washington navel orange | SW | N | Y | N | N |
| Seika navel orange | SW | N | Y | N | N |
| Newhall navel orange | SW | N | Y | N | N |
| Feicui pomelo | P | N | N | Y | N |
| Kaopan pomelo | P | N | N | Y | N |
| Huanong red pomelo | P | N | N | Y | N |
| Liangping pomelo | P | N | N | Y | N |
| Wanbai pomelo | P | N | N | Y | N |
| Low-acid pomelo | P | N | N | Y | N |
| Finger citron | Lem | N | N | N | Y |
| Tahiti lime | Lem | N | N | Y | Y |
| Kesai lime | Lem | Y | Y | N | Y |
| Verna lemon | Lem | N | N | N | Y |
| Eureka lemon | Lem | N | N | N | Y |
| Accuracy | 90.0% | 96.7% | 96.7% | 100.00% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wen, H.; Chen, J.; Peng, Z.; Shi, M.; Chen, M.; Yuan, Z.; Liu, Y.; Zhang, H.; Xu, J. Volatile Compounds in Fruit Peels as Novel Biomarkers for the Identification of Four Citrus Species. Molecules 2019, 24, 4550. https://doi.org/10.3390/molecules24244550
Zhang H, Wen H, Chen J, Peng Z, Shi M, Chen M, Yuan Z, Liu Y, Zhang H, Xu J. Volatile Compounds in Fruit Peels as Novel Biomarkers for the Identification of Four Citrus Species. Molecules. 2019; 24(24):4550. https://doi.org/10.3390/molecules24244550
Chicago/Turabian StyleZhang, Haipeng, Huan Wen, Jiajing Chen, Zhaoxin Peng, Meiyan Shi, Mengjun Chen, Ziyu Yuan, Yuan Liu, Hongyan Zhang, and Juan Xu. 2019. "Volatile Compounds in Fruit Peels as Novel Biomarkers for the Identification of Four Citrus Species" Molecules 24, no. 24: 4550. https://doi.org/10.3390/molecules24244550
APA StyleZhang, H., Wen, H., Chen, J., Peng, Z., Shi, M., Chen, M., Yuan, Z., Liu, Y., Zhang, H., & Xu, J. (2019). Volatile Compounds in Fruit Peels as Novel Biomarkers for the Identification of Four Citrus Species. Molecules, 24(24), 4550. https://doi.org/10.3390/molecules24244550