
Ingredients in Zijuan Pu’er Tea Extract Alleviate β-Amyloid Peptide Toxicity in a Caenorhabditis elegans Model of Alzheimer’s Disease Likely through DAF-16

 and
and
Abstract
:1. Introduction
2. Results
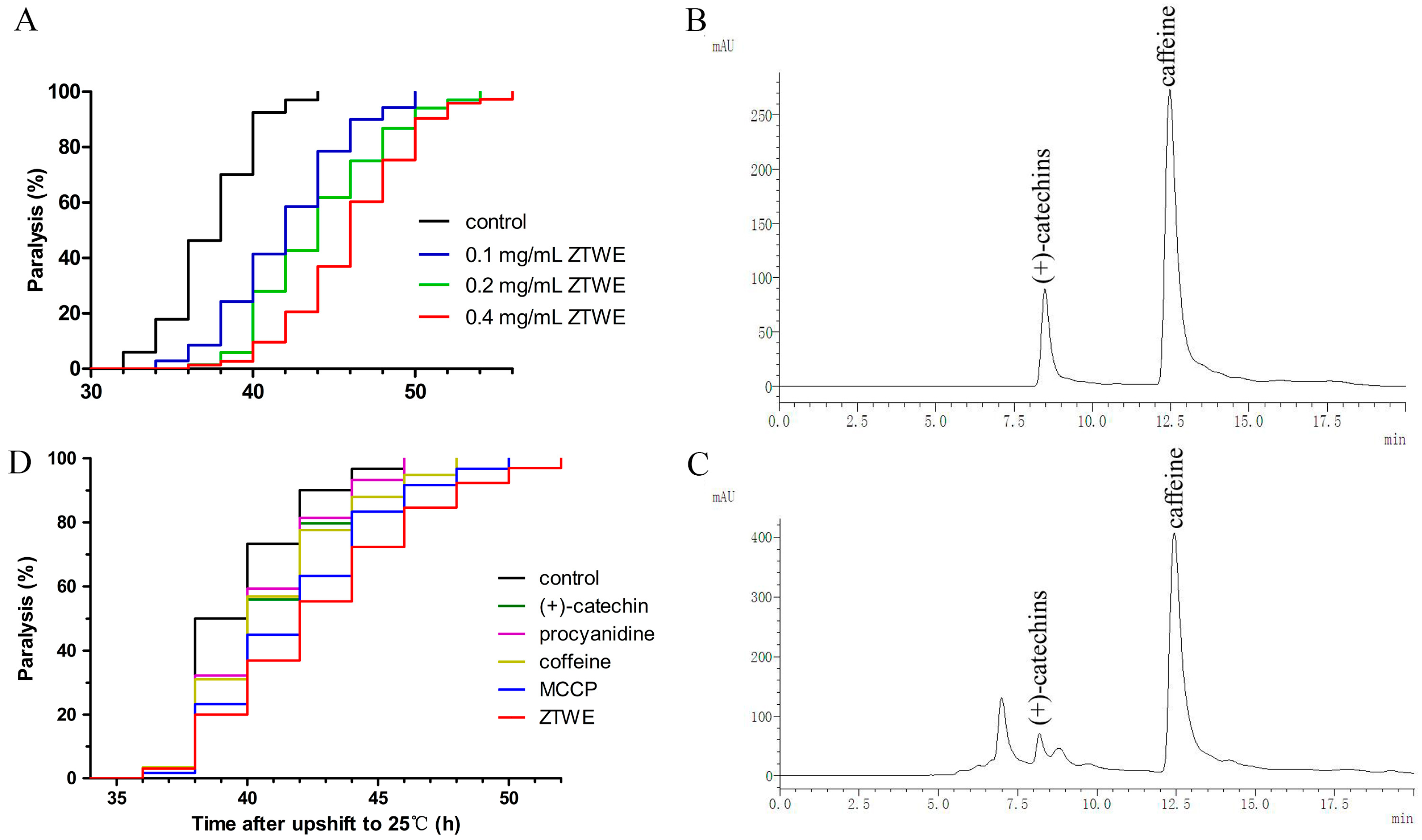
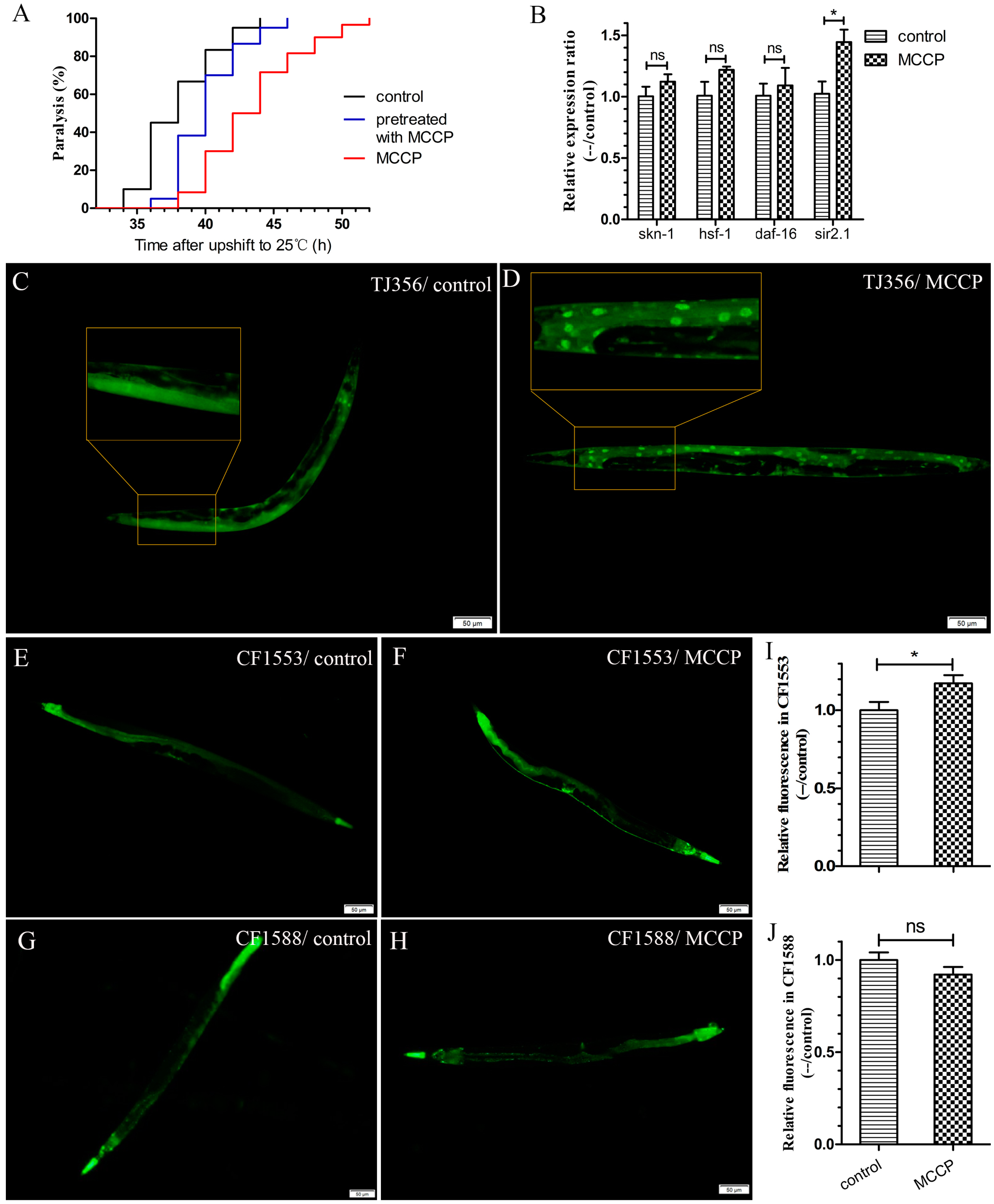
2.1. ZTWE Delays the Progression of Paralysis in AD Worms
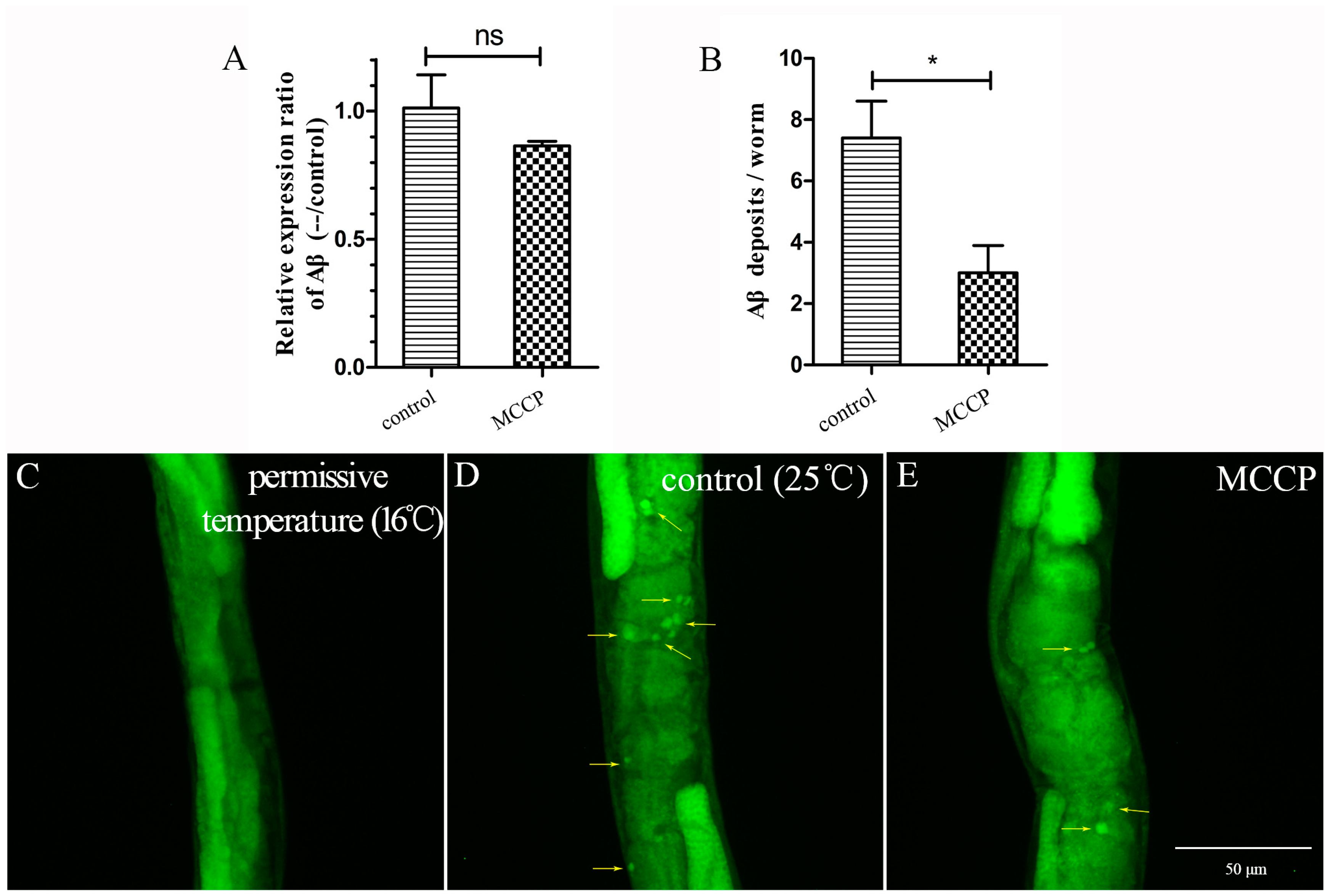
2.2. MCCP Reduces Aβ Deposits
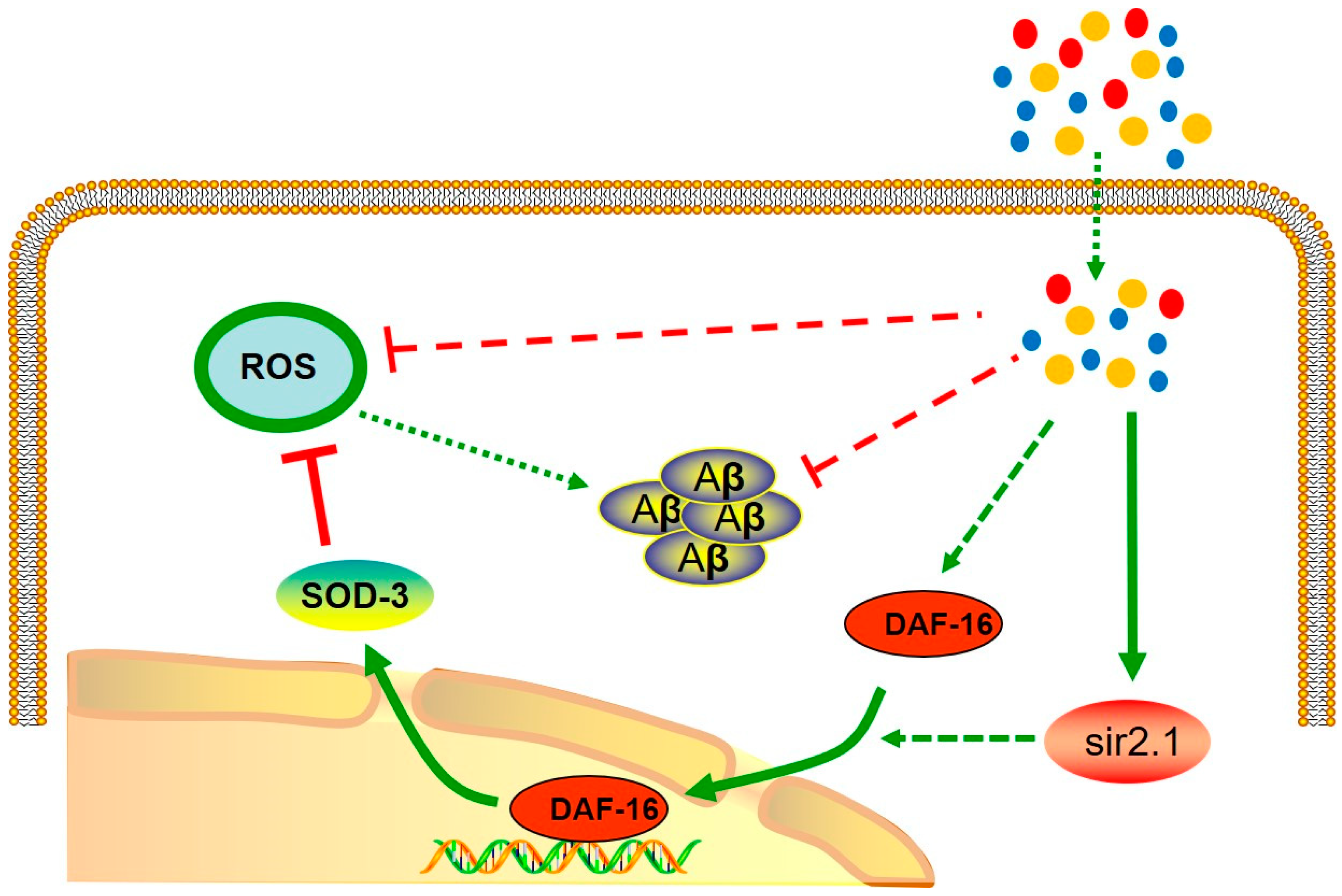
2.3. MCCP Might Requires DAF-16 to Protect AD Worms against Aβ Toxicity
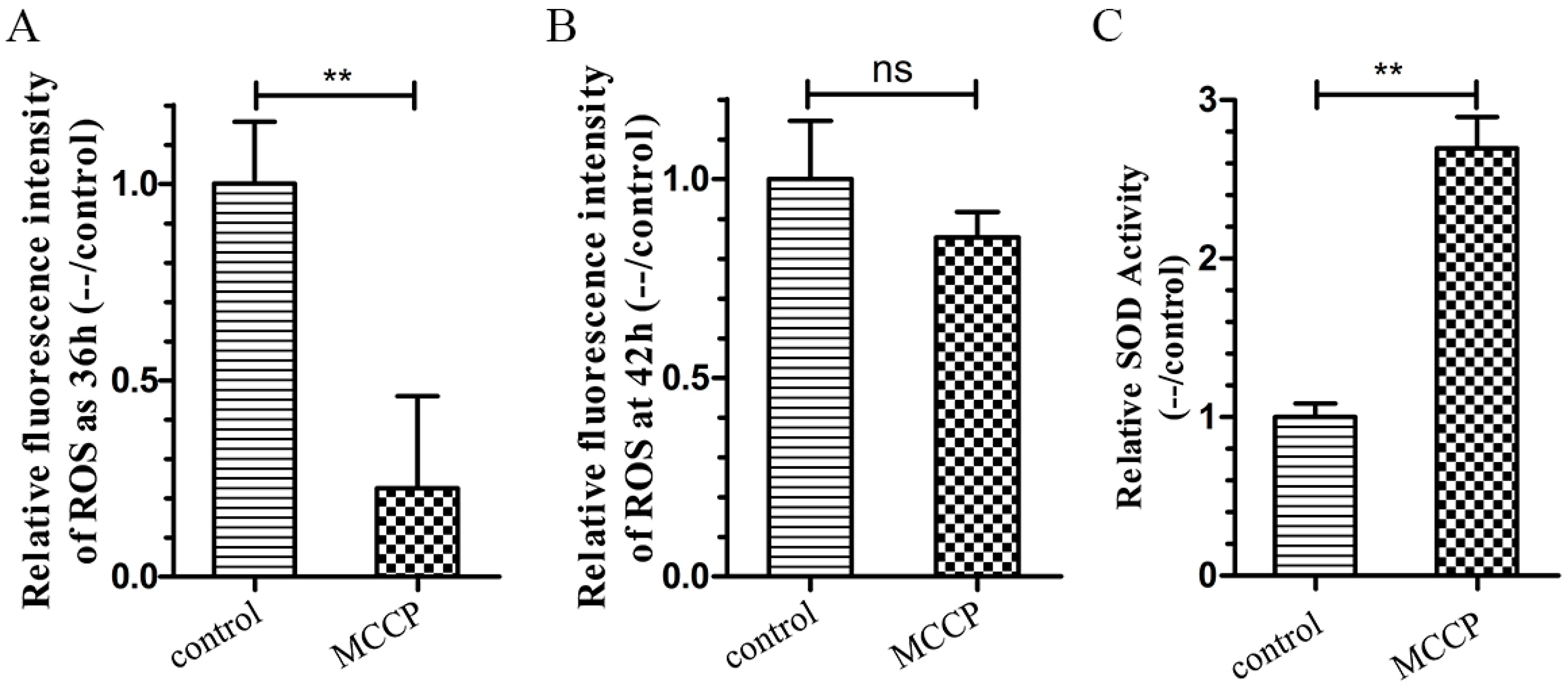
2.4. MCCP Increases Oxidative Stress Resistance in Transgenic C. elegans
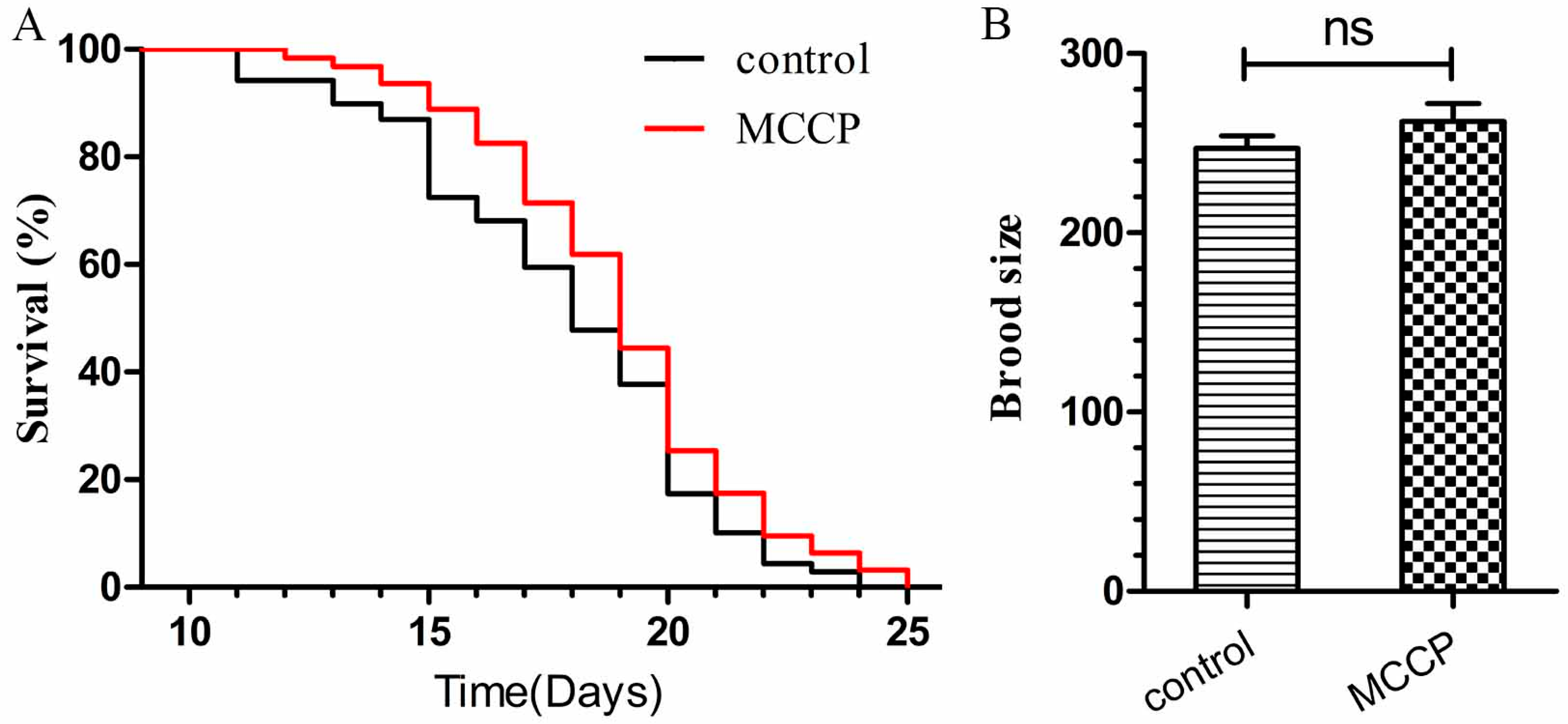
2.5. MCCP Has No Toxicity Effects to Wild Type C. elegans
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Preparation of Zijuan Pu’er Tea Extract
4.3. Life Span Assays
4.4. Brood Size Assays
4.5. Worm Paralysis Assays
4.6. Staining of Aβ
4.7. DAF-16 Localization via Fluorescence Microscopy
4.8. SOD-3 Expression via Fluorescence Microscopy
4.9. Measurement of ROS and SOD Activity in C. elegans
4.10. Expression Analysis by Quantitative Real Time RCR
| amy-1: | forward, 5′-CCGACATGACTCAGGATATGAAGT-3′, |
| reverse, 5′-CACCATGAGTCCAATGATTGCA-3′; | |
| sir-2.1: | forward, 5′-AGAACGCGCATTTCGCCATATTAAG-3′, |
| reverse, 5′-ATACTGACACTCCAGCGCCAG-3′; | |
| skn-1: | forward, 5′-AGTGTCGGCGTTCCAGATTTC-3′, |
| reverse, 5′-GTCGACGAACTTGCGAATCA-3′; | |
| hsf-1: | forward, 5′-TTGACGACGACAAGCTTCCAGT-3′, |
| reverse, 5′-AAAGCTTGCACCAGAATCATCCC-3′; | |
| daf-16: | forward, 5′-TTTCCGTCCCCGAACTCAA-3′, |
| reverse, 5′-ATTCGCCAACCCATGATGG-3′; | |
| act-1: | forward, 5′-CCAGGAATTGCTGATCGTATGCAGAA-3′, |
| reverse, 5′-TGGAGAGGGAAGCGAGGATAGA-3′. |
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Goedert, M.; Spillantini, M.G. A Century of Alzheimer’s Disease. Science 2006, 314, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Penke, B.; Tóth, A.M.; Földi, I.; Szűcs, M.; Janáky, T. Intraneuronal β-amyloid and its interactions with proteins and subcellular organelles. Electrophoresis 2012, 33, 3608–3616. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.M.; Townsend, M.; Podlisny, M.B.; Shankar, G.M.; Fadeeva, J.V.; El, A.O.; Hartley, D.M.; Selkoe, D.J. Certain inhibitors of synthetic amyloid beta-peptide (Abeta) fibrillogenesis block oligomerization of natural Abeta and thereby rescue long-term potentiation. J. Neurosci. 2005, 25, 2455. [Google Scholar] [CrossRef] [PubMed]
- Sangha, J.S.; Wally, O.; Banskota, A.H.; Stefanova, R.; Hafting, J.T.; Critchley, A.T.; Prithiviraj, B. A Cultivated Form of a Red Seaweed (Chondrus crispus), Suppresses β-Amyloid-Induced Paralysis in Caenorhabditis elegans. Marine Drugs 2015, 13, 6407–6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemere, C.A.; Masliah, E. Can Alzheimer disease be prevented by amyloid-β immunotherapy? Nat. Rev. Neurology 2010, 6, 108. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Abbas, S.; Wink, M. Epigallocatechin gallate inhibits beta amyloid oligomerization in Caenorhabditis elegans and affects the daf-2/insulin-like signaling pathway. Phytomedicine 2010, 17, 902–909. [Google Scholar] [CrossRef]
- Yang, Y.; Yong, J.Q.; Yip, Y.W.Y.; Chan, K.P.; Kai, O.C.; Chu, W.K.; Ng, T.K.; Chi, P.P.; Sun, O.C. Green tea catechins are potent anti-oxidants that ameliorate sodium iodate-induced retinal degeneration in rats. Sci. Rep. 2016, 6, 29546. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.; Liu, J.; Jie, R.; Fan, H.; Ou, H.; Ding, Y.; Zhang, Y.; Ma, R.; An, Y.; Liu, J. Green Tea Catechin-Based Complex Micelles Combined with Doxorubicin to Overcome Cardiotoxicity and Multidrug Resistance. Theranostics 2016, 6, 1277–1292. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, A.; Watanabe, T.; Fujita, T.; Hasegawa, T.; Saito, M.; Suganuma, M. Green tea aroma fraction reduces β-amyloid peptide-induced toxicity in Caenorhabditis elegans transfected with human β-amyloid minigene. Biosci. Biotechnol. Biochem. 2014, 78, 1206–1211. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.P.; Peng, C.X.; Gao, B.; Gong, J.S. Influence of large molecular polymeric pigments isolated from fermented Zijuan tea on the activity of key enzymes involved in lipid metabolism in rat. Exp. Gerontol. 2012, 47, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.P.; Fan, C.; Dong, W.M.; Gao, B.; Yuan, W.; Gong, J.S. Free radical scavenging and anti-oxidative activities of an ethanol-soluble pigment extract prepared from fermented Zijuan Pu-erh tea. Food Chem. Toxicol. 2013, 59, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Shen, X.; Shoji, T.; Kanda, T.; Zhou, J.; Zhao, L. Characterization and Activity of Anthocyanins in Zijuan Tea (Camellia sinensis var. kitamura). J. Agric. Food Chem. 2013, 61, 3306–3310. [Google Scholar] [CrossRef] [PubMed]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discovery 2006, 5, 387–398. [Google Scholar] [CrossRef]
- Wolozin, B.; Gabel, C.; Ferree, A.; Guillily, M.; Ebata, A. Watching Worms Whither: Modeling Neurodegeneration in C. elegans. Progress in Molecular Biology and Translational Science; Academic Press: Cambridge, MA, USA, 2011; Volume 100, pp. 499–514. [Google Scholar]
- Ewald, C.Y.; Li, C. Understanding the molecular basis of Alzheimer’s disease using a Caenorhabditis elegans model system. Brain Struct. Funct. 2010, 214, 263–283. [Google Scholar] [CrossRef] [PubMed]
- Lublin, A.L.; Link, C.D. Alzheimer’s disease drug discovery: In vivo screening using Caenorhabditis elegans as a model for β-amyloid peptide-induced toxicity. Drug Discovery Today Technol. 2013, 10, e115–e119. [Google Scholar] [CrossRef] [PubMed]
- Sola, I.; Viayna, E.; Gómez, T.; Galdeano, C.; Cassina, M.; Camps, P.; Romeo, M.; Diomede, L.; Salmona, M.; Franco, P.; et al. Multigram Synthesis and in Vivo Efficacy Studies of a Novel Multitarget Anti-Alzheimer’s Compound. Molecules 2015, 20, 4492–4515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y. Alzheimer’s disease, the nematode Caenorhabditis elegans, and ginkgo biloba leaf extract. Life Sci. 2006, 78, 2066–2072. [Google Scholar] [CrossRef]
- Link, P.; Wetterauer, B.; Fu, Y.; Wink, M. Extracts of Glycyrrhiza uralensis and isoliquiritigenin counteract amyloid-β toxicity in Caenorhabditis elegans. Planta Med. 2015, 81, 357–362. [Google Scholar] [CrossRef]
- Guo, H.; Cao, M.; Zou, S.; Ye, B.; Dong, Y. Cranberry Extract Standardized for Proanthocyanidins Alleviates β-Amyloid Peptide Toxicity by Improving Proteostasis Through HSF-1 in Caenorhabditis elegans Model of Alzheimer’s Disease. J. Gerontol. Series A 2015, 71, 1564–1573. [Google Scholar] [CrossRef]
- Azevêdo, J.C.S.; Borges, K.C.; Genovese, M.I.; Correia, R.T.P.; Vattem, D.A. Neuroprotective effects of dried camu-camu ( Myrciaria dubia HBK McVaugh) residue in C. elegans. Food Res. Int. 2015, 73, 135–141. [Google Scholar] [CrossRef]
- Wang, X.; Cao, M.; Dong, Y. Royal jelly promotes DAF-16-mediated proteostasis to tolerate β-amyloid toxicity in C. elegans model of Alzheimer’s disease. Oncotarget 2016, 7, 54183–54193. [Google Scholar] [PubMed]
- Ganguly, G.; Chakrabarti, S.; Chatterjee, U.; Saso, L. Proteinopathy, oxidative stress and mitochondrial dysfunction: Cross talk in Alzheimer’s disease and Parkinson’s disease. Drug Des. Dev. Ther. 2017, 11, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Dostal, V.; Roberts, C.M.; Link, C.D. Genetic mechanisms of coffee extract protection in a Caenorhabditis elegans model of β-amyloid peptide toxicity. Genetics 2010, 186, 857. [Google Scholar] [CrossRef] [PubMed]
- Regitz, C.; Wenzel, U. Amyloid-beta (Aβ 1–42)-induced paralysis in Caenorhabditis elegans is reduced by restricted cholesterol supply. Neurosci. Lett. 2014, 576, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Ala, B.; Mohan, V.; H Robert, H.; Leonard, G. C. elegans SIR-2.1 interacts with 14-3-3 proteins to activate DAF-16 and extend life span. Cell 2006, 125, 1165–1177. [Google Scholar]
- Henderson, S.T.; Johnson, T.E. daf-16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans. Curr. Biol. 2001, 11, 1975–1980. [Google Scholar] [CrossRef]
- Iwatsubo, T.; Odaka, A.; Suzuki, N.; Mizusawa, H.; Nukina, N.; Ihara, Y. Visualization of A beta 42(43) and A beta 40 in senile plaques with end-specific A beta monoclonals: Evidence that an initially deposited species is A beta 42(43). Neuron 1994, 13, 45–53. [Google Scholar] [CrossRef]
- Vassar, R.; Bennett, B.D.; BabuKhan, S.; Kahn, S.; Mendiaz, E.A.; Denis, P.; Teplow, D.B.; Ross, S.; Amarante, P.; Loeloff, R. β-Secretase Cleavage of Alzheimer’s Amyloid Precursor Protein by the Transmembrane Aspartic Protease BACE. Science 1999, 286, 735–741. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The Amyloid Hypothesis of Alzheimer’s Disease: Progress and Problems on the Road to Therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Pereira, C.; Agostinho, P.; Moreira, P.I.; Cardoso, S.M.; Oliveira, C.R. Alzheimer’s disease-associated neurotoxic mechanisms and neuroprotective strategies. Curr. Drug Targets CNS Neurological Disorders 2005, 4. [Google Scholar] [CrossRef]
- Peng, C.; Chan, H.Y.; Li, Y.M.; Huang, Y.; Chen, Z.Y. Black tea theaflavins extend the lifespan of fruit flies. Exp. Gerontol. 2009, 44, 773–783. [Google Scholar] [CrossRef]
- Shukla, V.; Mishra, S.K.; Pant, H.C. Oxidative Stress in Neurodegeneration. Adv. Pharmacol. Sci. 2011, 2011, 572634. [Google Scholar] [CrossRef] [PubMed]
- Dumont, M.; Wille, E.; Stack, C.; Calingasan, N.Y.; Beal, M.F.; Lin, M.T. Reduction of oxidative stress, amyloid deposition, and memory deficit by manganese superoxide dismutase overexpression in a transgenic mouse model of Alzheimer’s disease. FASEB J. 2009, 23, 2459–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascella, R.; Evangelisti, E.; Zampagni, M.; Becatti, M.; D’Adamio, G.; Goti, A.; Liguri, G.; Fiorillo, C.; Cecchi, C. S-linolenoyl glutathione intake extends life-span and stress resistance via Sir-2.1 upregulation in Caenorhabditis elegans. Free Radical Biol. Med. 2014, 73, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.I.; Pincus, Z.; Slack, F.J. Longevity and stress in Caenorhabditis elegans. Aging 2011, 3, 733–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Link, C.D. C. elegans models of age-associated neurodegenerative diseases: Lessons from transgenic worm models of Alzheimer’s disease. Exp. Gerontol. 2006, 41, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Fu, X.; Yi, L.; Du, F.; Hua, W.; Shu, X.; Li, W.; Ma, J. 2-SeCD treatment extends lifespan, improves healthspan and enhances resistance to stress in Caenorhabditis elegans. Rsc Adv. 2017, 7, 48245–48252. [Google Scholar] [Green Version]
- Kim, Y.; Bandyopadhyay, J.; Cho, I.; Lee, J.; Park, D.H.; Cho, J.H. Nucleolar GTPase NOG-1 Regulates Development, Fat Storage, and Longevity through Insulin/IGF Signaling in C. elegans. Mol. Cells 2014, 37, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, L.; Bai, S.; Xing, S.; Li, W.; Ma, J.; Fu, X. Knockdown of LMW-PTP enhances stress resistance in Caenorhabditis elegans. Int. J. Biol. Macromol. 2018, 113, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Fu, X.; Jiang, L.; Lu, W.; Bai, S.; Yan, J.; Shu, X.; Li, W.; Ma, J. Arbutin increasesCaenorhabditis eleganslongevity and stress resistance. PeerJ 2017, 5, e4170. [Google Scholar]
- Kampkötter, A.; Nkwonkam, C.G.; Zurawski, R.F.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. Investigations of protective effects of the flavonoids quercetin and rutin on stress resistance in the model organism Caenorhabditis elegans. Toxicology 2007, 234, 113–123. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Sample of the Zijuan Pu’er tea powder is available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard Curve | R2 | |
|---|---|---|
| (+)-catechins | Y = (1.53376 × 107)X + 47476.3 | 0.9997 |
| caffeine | Y = (7.26906 × 107)X + 245960 | 0.9995 |
| procyanidins | Y = 0.242X + 0.0041 | 0.9983 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, F.; Zhou, L.; Jiao, Y.; Bai, S.; Wang, L.; Ma, J.; Fu, X. Ingredients in Zijuan Pu’er Tea Extract Alleviate β-Amyloid Peptide Toxicity in a Caenorhabditis elegans Model of Alzheimer’s Disease Likely through DAF-16. Molecules 2019, 24, 729. https://doi.org/10.3390/molecules24040729
Du F, Zhou L, Jiao Y, Bai S, Wang L, Ma J, Fu X. Ingredients in Zijuan Pu’er Tea Extract Alleviate β-Amyloid Peptide Toxicity in a Caenorhabditis elegans Model of Alzheimer’s Disease Likely through DAF-16. Molecules. 2019; 24(4):729. https://doi.org/10.3390/molecules24040729
Chicago/Turabian StyleDu, Fangzhou, Lin Zhou, Yan Jiao, Shuju Bai, Lu Wang, Junfeng Ma, and Xueqi Fu. 2019. "Ingredients in Zijuan Pu’er Tea Extract Alleviate β-Amyloid Peptide Toxicity in a Caenorhabditis elegans Model of Alzheimer’s Disease Likely through DAF-16" Molecules 24, no. 4: 729. https://doi.org/10.3390/molecules24040729
APA StyleDu, F., Zhou, L., Jiao, Y., Bai, S., Wang, L., Ma, J., & Fu, X. (2019). Ingredients in Zijuan Pu’er Tea Extract Alleviate β-Amyloid Peptide Toxicity in a Caenorhabditis elegans Model of Alzheimer’s Disease Likely through DAF-16. Molecules, 24(4), 729. https://doi.org/10.3390/molecules24040729