
Natural Products and Obesity: A Focus on the Regulation of Mitotic Clonal Expansion during Adipogenesis


Abstract
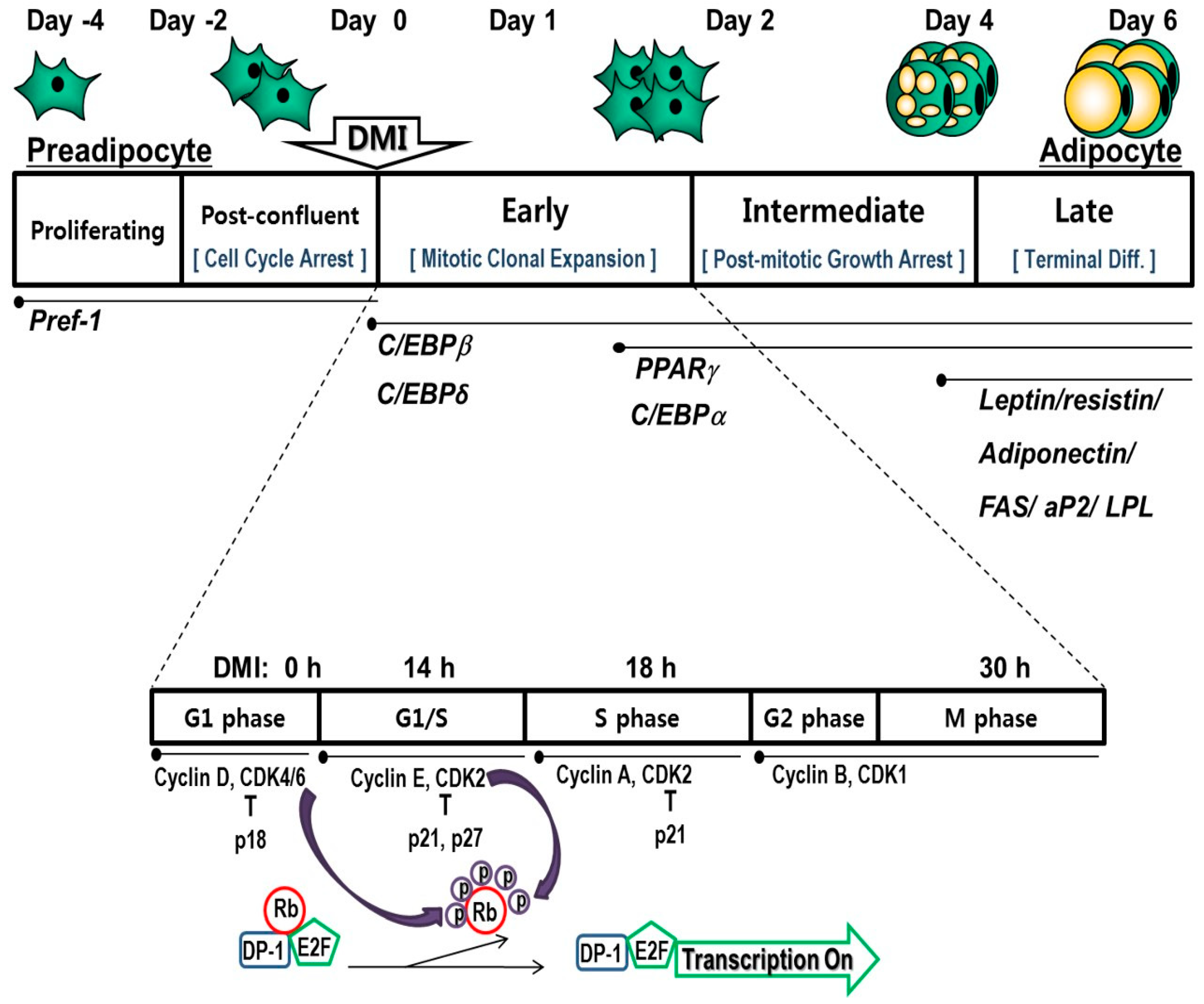
:1. Introduction
2. Adipose Biology during the Development of Obesity
3. Regulation of Adipogenesis
3.1. Arrest and Progression of the Cell Cycle and Its Associated Regulatory Proteins
3.2. Cascade of Transcriptional Factors during Mitotic Clonal Expansion
3.2.1. CCAAT/Enhancer Binding Protein β (C/EBPβ)
3.2.2. CCAAT/Enhancer Binding Protein α (C/EBPα)
3.2.3. Peroxisome Proliferator-Activated Receptor γ (PPARγ)
3.2.4. Gene Markers for Terminal Differentiation
3.3. Signaling Pathways Involved in Adipogenesis
3.3.1. Phosphoinositide 3-Kinase/Protein Kinase B (PI3K/AKT) Pathway
3.3.2. Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase (MAPK/ERK) Pathway
3.3.3. Wingless/INT-1 Protein (Wnt)/β-Catenin Signaling
3.3.4. AMP-Activated Protein Kinase (AMPK) Pathway
4. Regulation of Adipogenesis by Various Phytochemicals
4.1. Cell-Cycle Arrest and Its Regulatory Proteins
4.2. Cell Proliferation
4.3. Transcription Factors
4.4. Intracellular Signaling Pathways
4.4.1. Phosphoinositide 3-Kinase/Protein Kinase B (PI3K/AKT) Pathway
4.4.2. Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase (MAPK/ERK) Pathway
4.4.3. Wingless/INT-1 Protein (Wnt)/β-Catenin Signaling
4.4.4. AMP-Activated Protein Kinase (AMPK) Pathway
5. Inhibition of Adipogenesis by Natural Product-Derived Bioactive Components
5.1. Combination of Different Phytochemicals
5.2. Combination of Phytochemicals with Other Compounds Isolated from Natural Product
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Batsis, J.A.; Nieto-Martinez, R.E.; Lopez-Jimenez, F. Metabolic syndrome: From global epidemiology to individualized medicine. Clin. Pharmacol. Ther. 2007, 82, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity: Implications for metabolic syndrome, diabetes, hypertension, dyslipidemia, atherosclerosis, and cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; McPherson, K.; Marsh, T.; Gortmaker, S.L.; Brown, M. Health and economic burden of the projected obesity trends in the USA and the UK. Lancet 2011, 378, 815–825. [Google Scholar] [CrossRef]
- Finkelstein, E.A.; Khavjou, O.A.; Thompson, H.; Trogdon, J.G.; Pan, L.; Sherry, B.; Dietz, W. Obesity and severe obesity forecasts through 2030. Am. J. Prev. Med. 2012, 42, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.J. Beneficial health effects of modest weight loss. Int. J. Obes. Relat. Metab. Disord. 1992, 16, 397–415. [Google Scholar]
- Kiess, W.; Reich, A.; Muller, G.; Meyer, K.; Galler, A.; Bennek, J.; Kratzsch, J. Clinical aspects of obesity in childhood and adolescence—Diagnosis, treatment and prevention. Int. J. Obes. Relat. Metab. Disord. 2001, 25 (Suppl. 1), S75–S79. [Google Scholar] [CrossRef]
- Stern, J.S.; Hirsch, J.; Blair, S.N.; Foreyt, J.P.; Frank, A.; Kumanyika, S.K.; Madans, J.H.; Marlatt, G.A.; St Jeor, S.T.; Stunkard, A.J. Weighing the options: Criteria for evaluating weight-management programs. The committee to develop criteria for evaluating the outcomes of approaches to prevent and treat obesity. Obes. Res. 1995, 3, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Cheung, B.M.Y.; Cheung, T.T.; Samaranayake, N.R. Safety of antiobesity drugs. Ther. Adv. Drug. Saf. 2013, 4, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.F.; Ashraf, S.A.; Ahmad, F.A.; Ansari, J.A.; Siddiquee, M.R.A. Nutraceutical market and its regulation. Am. J. Food Tech. 2011, 6, 342–347. [Google Scholar] [CrossRef]
- Sun, N.N.; Wu, T.Y.; Chau, C.F. Natural Dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, S.; Chen, Y.; Saravanan, D.; Sundram, K.M.; Yoga Latha, L. Extraction, isolation and characterization of bioactive compounds from plants’ extracts. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Catalan, V.; Gomez-Ambrosi, J.; Fruhbeck, G. Visceral and subcutaneous adiposity: Are both potential therapeutic targets for tackling the metabolic syndrome? Curr. Pharm. Des. 2007, 13, 2169–2175. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Arner, P.; Spalding, K.L. Fat cell turnover in humans. Biochem. Biophys. Res. Commun. 2010, 396, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Reuss, L.; Wang, Y. Potential of natural products in the inhibition of adipogenesis through regulation of PPARγ expression and/or its transcriptional activity. Molecules 2016, 21, 1278. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. Molecular regulation of adipogenesis. Annu. Rev. Cell Dev. Biol. 2000, 16, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Otto, T.C.; Lane, M.D. Adipose development: From stem cell to adipocyte. Crit. Rev. Biochem. Mol. Biol. 2005, 40, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.E.; Florine, D.L.; Wille, J.J., Jr.; Yun, K. Coupling of growth arrest and differentiation at a distinct state in the G1 phase of the cell cycle: GD. Proc. Natl. Acad. Sci. USA 1982, 79, 845–849. [Google Scholar] [CrossRef]
- Pairault, J.; Green, H. A study of the adipose conversion of suspended 3T3 cells by using glycerophosphate dehydrogenase as differentiation marker. Proc. Natl. Acad. Sci. USA 1979, 76, 5138–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichert, M.; Eick, D. Analysis of cell cycle arrest in adipocyte differentiation. Oncogene 1999, 18, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. Mitotic clonal expansion: A synchronous process required for adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, R.F.; Farmer, S.R. Role of PPARγ in regulating a cascade expression of cyclin-dependent kinase inhibitors, p18(INK4c) and p21(Waf1/Cip1), during adipogenesis. J. Biol. Chem. 1999, 274, 17088–17097. [Google Scholar] [CrossRef]
- Shoba, G.; Joy, D.; Joseph, T.; Majeed, M.; Rajendran, R.; Srinivas, P.S. Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta. Med. 1998, 64, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Wei, Y.; Chen, N.; Jiang, M.; Wu, J.; Liao, K. DNA synthesis and mitotic clonal expansion is not a required step for 3T3-L1 preadipocyte differentiation into adipocytes. J. Biol. Chem. 2001, 276, 11988–11995. [Google Scholar] [CrossRef] [PubMed]
- MacDougald, O.A.; Mandrup, S. Adipogenesis: Forces that tip the scales. Trends Endocrinol. Metab. 2002, 13, 5–11. [Google Scholar] [CrossRef]
- Wu, J.; Srinivasan, S.V.; Neumann, J.C.; Lingrel, J.B. The KLF2 transcription factor does not affect the formation of preadipocytes but inhibits their differentiation into adipocytes. Biochemistry 2005, 44, 11098–11105. [Google Scholar] [CrossRef] [PubMed]
- Mei, B.; Zhao, L.; Chen, L.; Sul, H.S. Only the large soluble form of preadipocyte factor-1 (Pref-1), but not the small soluble and membrane forms, inhibits adipocyte differentiation: Role of alternative splicing. Biochem. J. 2002, 364, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.S.; Smas, C.M.; Lee, K.; Villena, J.A.; Kim, K.H.; Yun, E.J.; Sul, H.S. Mice lacking paternally expressed Pref-1/Dlk1 display growth retardation and accelerated adiposity. Mol. Cell Biol. 2002, 22, 5585–5592. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Lane, M.D. Role of C/EBP homologous protein (CHOP-10) in the programmed activation of CCAAT/enhancer-binding protein-beta during adipogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 12446–12450. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.; Dalgin, G.; Xu, H.; Ting, C.N.; Leiden, J.M.; Hotamisligil, G.S. Function of GATA transcription factors in preadipocyte-adipocyte transition. Science 2000, 290, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of adipogenesis by Wnt signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Bucher, N.L.; Farmer, S.R. Induction of peroxisome proliferator-activated receptor gamma during the conversion of 3T3 fibroblasts into adipocytes is mediated by C/EBPbeta, C/EBPdelta, and glucocorticoids. Mol. Cell Biol. 1996, 16, 4128–4136. [Google Scholar] [CrossRef]
- Smas, C.M.; Chen, L.; Zhao, L.; Latasa, M.J.; Sul, H.S. Transcriptional repression of pref-1 by glucocorticoids promotes 3T3-L1 adipocyte differentiation. J. Biol. Chem. 1999, 274, 12632–12641. [Google Scholar] [CrossRef]
- Cao, Z.; Umek, R.M.; McKnight, S.L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991, 5, 1538–1552. [Google Scholar] [CrossRef]
- Jin, S.; Zhai, B.; Qiu, Z.; Wu, J.; Lane, M.D.; Liao, K. c-Crk, a substrate of the insulin-like growth factor-1 receptor tyrosine kinase, functions as an early signal mediator in the adipocyte differentiation process. J. Biol. Chem. 2000, 275, 34344–34352. [Google Scholar] [CrossRef]
- Huang, H.; Lane, M.D.; Tang, Q.Q. Effect of serum on the down-regulation of CHOP-10 during differentiation of 3T3-L1 preadipocytes. Biochem. Biophys. Res. Commun. 2005, 338, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef]
- Park, B.H.; Qiang, L.; Farmer, S.R. Phosphorylation of C/EBPβ at a consensus extracellular signal-regulated kinase/glycogen synthase kinase 3 site is required for the induction of adiponectin gene expression during the differentiation of mouse fibroblasts into adipocytes. Mol. Cell. Biol. 2004, 24, 8671–8680. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Gronborg, M.; Huang, H.; Kim, J.W.; Otto, T.C.; Pandey, A.; Lane, M.D. Sequential phosphorylation of CCAAT enhancer-binding protein β by MAPK and glycogen synthase kinase 3β is required for adipogenesis. Proc. Natl. Acad. Sci. USA 2005, 102, 9766–9771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Kim, J.W.; Gronborg, M.; Urlaub, H.; Lane, M.D.; Tang, Q.Q. Role of cdk2 in the sequential phosphorylation/activation of C/EBPβ during adipocyte differentiation. Proc. Natl. Acad. Sci. USA 2007, 104, 11597–11602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.Q.; Lane, M.D. Activation and centromeric localization of CCAAT/enhancer-binding proteins during the mitotic clonal expansion of adipocyte differentiation. Genes Dev. 1999, 13, 2231–2241. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Tang, Q.Q.; Li, X.; Lane, M.D. Effect of phosphorylation and S–S bond-induced dimerization on DNA binding and transcriptional activation by C/EBPβ. Proc. Natl. Acad. Sci. USA 2007, 104, 1800–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. CCAAT/enhancer-binding protein β is required for mitotic clonal expansion during adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 850–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.W.; Tang, Q.Q.; Vinson, C.; Lane, M.D. Dominant-negative C/EBP disrupts mitotic clonal expansion and differentiation of 3T3-L1 preadipocytes. Proc. Natl. Acad. Sci. USA 2004, 101, 43–47. [Google Scholar] [CrossRef]
- Sprott, K.M.; Chumley, M.J.; Hanson, J.M.; Dobrowsky, R.T. Decreased activity and enhanced nuclear export of CCAAT-enhancer-binding protein beta during inhibition of adipogenesis by ceramide. Biochem. J. 2002, 365, 181–191. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Zhang, J.W.; Daniel Lane, M. Sequential gene promoter interactions of C/EBPβ, C/EBPα, and PPARγ during adipogenesis. Biochem. Biophys. Res. Commun. 2004, 319, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Darlington, G.J.; Wang, N.; Hanson, R.W. C/EBP α: A critical regulator of genes governing integrative metabolic processes. Curr. Opin. Genet. Dev. 1995, 5, 565–570. [Google Scholar] [CrossRef]
- Shao, D.; Lazar, M.A. Peroxisome proliferator activated receptor γ, CCAAT/enhancer-binding protein α, and cell cycle status regulate the commitment to adipocyte differentiation. J. Biol. Chem. 1997, 272, 21473–21478. [Google Scholar] [CrossRef] [PubMed]
- Christy, R.J.; Kaestner, K.H.; Geiman, D.E.; Lane, M.D. CCAAT/enhancer binding protein gene promoter: Binding of nuclear factors during differentiation of 3T3-L1 preadipocytes. Proc. Natl. Acad. Sci. USA 1991, 88, 2593–2597. [Google Scholar] [CrossRef] [PubMed]
- Gale, S.E.; Frolov, A.; Han, X.; Bickel, P.E.; Cao, L.; Bowcock, A.; Schaffer, J.E.; Ory, D.S. A regulatory role for 1-acylglycerol-3-phosphate-O-acyltransferase 2 in adipocyte differentiation. J. Biol. Chem. 2006, 281, 11082–11089. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.K.; Lee, M.Y.; Kim, J.W.; Kim, M.; Moon, J.S.; Lee, Y.J.; Ahn, Y.H.; Kim, K.S. Lipin1 is a key factor for the maturation and maintenance of adipocytes in the regulatory network with CCAAT/enhancer-binding protein α and peroxisome proliferator-activated receptor γ2. J. Biol. Chem. 2008, 283, 34896–34906. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Reue, K. Biochemistry, physiology, and genetics of GPAT, AGPAT, and lipin enzymes in triglyceride synthesis. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1195–E1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.R.; Barrick, C.; Kim, K.A.; Lindner, J.; Blondeau, B.; Fujimoto, Y.; Shiota, M.; Kesterson, R.A.; Kahn, B.B.; Magnuson, M.A. Deletion of PPARγ in adipose tissues of mice protects against high fat diet-induced obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2005, 102, 6207–6212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Londos, C.; Brasaemle, D.L.; Schultz, C.J.; Segrest, J.P.; Kimmel, A.R. Perilipins, ADRP, and other proteins that associate with intracellular neutral lipid droplets in animal cells. Semin. Cell Dev. Biol. 1999, 10, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Koerner, A.; Kratzsch, J.; Kiess, W. Adipocytokines: Leptin—the classical, resistin—the controversical, adiponectin—the promising, and more to come. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 525–546. [Google Scholar] [CrossRef] [PubMed]
- Payne, V.A.; Au, W.S.; Lowe, C.E.; Rahman, S.M.; Friedman, J.E.; O’Rahilly, S.; Rochford, J.J. C/EBP transcription factors regulate SREBP1c gene expression during adipogenesis. Biochem. J. 2009, 425, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Zagariya, A.; Mungre, S.; Lovis, R.; Birrer, M.; Ness, S.; Thimmapaya, B.; Pope, R. Tumor necrosis factor alpha gene regulation: Enhancement of C/EBPβ-induced activation by c-Jun. Mol. Cell Biol. 1998, 18, 2815–2824. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBPα and PPARγ controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPα induces adipogenesis through PPARγ: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Sakamoto, K. (−)-Epigallocatechin gallate suppresses adipocyte differentiation through the MEK/ERK and PI3K/Akt pathways. Cell Biol. Int. 2012, 36, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.M.; Lee, Y.S.; Sin, D.M.; Lee, S.; Lee, M.K.; Lee, Y.M.; Hong, J.T.; Yun, Y.P.; Yoo, H.S. Sulforaphane inhibits mitotic clonal expansion during adipogenesis through cell cycle arrest. Obesity 2012, 20, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.N.; Ross, S.E.; Longo, K.A.; Bajnok, L.; Hemati, N.; Johnson, K.W.; Harrison, S.D.; MacDougald, O.A. Regulation of Wnt signaling during adipogenesis. J. Biol. Chem. 2002, 277, 30998–31004. [Google Scholar] [CrossRef] [PubMed]
- Muise-Helmericks, R.C.; Grimes, H.L.; Bellacosa, A.; Malstrom, S.E.; Tsichlis, P.N.; Rosen, N. Cyclin D expression is controlled post-transcriptionally via a phosphatidylinositol 3-kinase/Akt-dependent pathway. J. Biol. Chem. 1998, 273, 29864–29872. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.; Medema, R.H.; Garcia-Cao, I.; Dubuisson, M.L.; Barradas, M.; Glassford, J.; Rivas, C.; Burgering, B.M.; Serrano, M.; Lam, E.W. Inhibition of the phosphoinositide 3-kinase pathway induces a senescence-like arrest mediated by p27Kip1. J. Biol. Chem. 2000, 275, 21960–21968. [Google Scholar] [CrossRef] [PubMed]
- Diehl, J.A.; Cheng, M.; Roussel, M.F.; Sherr, C.J. Glycogen synthase kinase-3β regulates cyclin D1 proteolysis and subcellular localization. Genes Dev. 1998, 12, 3499–3511. [Google Scholar] [CrossRef] [PubMed]
- Magun, R.; Burgering, B.M.; Coffer, P.J.; Pardasani, D.; Lin, Y.; Chabot, J.; Sorisky, A. Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3-L1 preadipose cells causes spontaneous differentiation. Endocrinology 1996, 137, 3590–3593. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, R.S.; Orena, S.J.; Rafidi, K.; Torchia, A.J.; Stock, J.L.; Hildebrandt, A.L.; Coskran, T.; Black, S.C.; Brees, D.J.; Wicks, J.R.; et al. Severe diabetes, age-dependent loss of adipose tissue, and mild growth deficiency in mice lacking Akt2/PKBβ. J. Clin. Investig. 2003, 112, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakae, J.; Kitamura, T.; Kitamura, Y.; Biggs, W.H., 3rd; Arden, K.C.; Accili, D. The forkhead transcription factor Foxo1 regulates adipocyte differentiation. Dev. Cell 2003, 4, 119–129. [Google Scholar] [CrossRef]
- Munekata, K.; Sakamoto, K. Forkhead transcription factor Foxo1 is essential for adipocyte differentiation. In Vitro Cell Dev. Biol. Anim. 2009, 45, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Prusty, D.; Park, B.H.; Davis, K.E.; Farmer, S.R. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor γ (PPARγ) and C/EBPα gene expression during the differentiation of 3T3-L1 preadipocytes. J. Biol. Chem. 2002, 277, 46226–46232. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, N.; Phillips, B.W.; Massiera, F.; Villageois, P.; Wdziekonski, B.; Saint-Marc, P.; Nichols, J.; Aubert, J.; Saeki, K.; Yuo, A.; et al. Activation of extracellular signal-regulated kinases and CREB/ATF-1 mediate the expression of CCAAT/enhancer binding proteins β and -δ in preadipocytes. Mol. Endocrinol. 2001, 15, 2037–2049. [Google Scholar]
- Worster, D.T.; Schmelzle, T.; Solimini, N.L.; Lightcap, E.S.; Millard, B.; Mills, G.B.; Brugge, J.S.; Albeck, J.G. Akt and ERK control the proliferative response of mammary epithelial cells to the growth factors IGF-1 and EGF through the cell cycle inhibitor p57Kip2. Sci. Signal. 2012, 5, ra19. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.; Kishida, S.; Yamamoto, H.; Murai, H.; Koyama, S.; Kikuchi, A. Axin, a negative regulator of the Wnt signaling pathway, forms a complex with GSK-3β and β-catenin and promotes GSK-3β-dependent phosphorylation of β-catenin. EMBO J. 1998, 17, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Krupnik, V.E.; Sokol, S.Y. Axis determination in Xenopus involves biochemical interactions of axin, glycogen synthase kinase 3 and β-catenin. Curr. Biol. 1998, 8, 591–594. [Google Scholar] [CrossRef]
- Sakanaka, C.; Weiss, J.B.; Williams, L.T. Bridging of β-catenin and glycogen synthase kinase-3β by axin and inhibition of β-catenin-mediated transcription. Proc. Natl. Acad. Sci. USA 1998, 95, 3020–3023. [Google Scholar] [CrossRef] [PubMed]
- Tauriello, D.V.; Jordens, I.; Kirchner, K.; Slootstra, J.W.; Kruitwagen, T.; Bouwman, B.A.; Noutsou, M.; Rudiger, S.G.; Schwamborn, K.; Schambony, A.; et al. Wnt/β-catenin signaling requires interaction of the Dishevelled DEP domain and C terminus with a discontinuous motif in Frizzled. Proc. Natl. Acad. Sci. USA 2012, 109, E812–E820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestwich, T.C.; Macdougald, O.A. Wnt/β-catenin signaling in adipogenesis and metabolism. Curr. Opin. Cell Biol. 2007, 19, 612–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cselenyi, C.S.; Jernigan, K.K.; Tahinci, E.; Thorne, C.A.; Lee, L.A.; Lee, E. LRP6 transduces a canonical Wnt signal independently of Axin degradation by inhibiting GSK3’s phosphorylation of β-catenin. Proc. Natl. Acad. Sci. USA 2008, 105, 8032–8037. [Google Scholar] [CrossRef] [PubMed]
- Joiner, D.M.; Ke, J.; Zhong, Z.; Xu, H.E.; Williams, B.O. LRP5 and LRP6 in development and disease. Trends Endocrinol. Metab. 2013, 24, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Mushiake, S.; Bessho, K.; Murakami, M.; Namba, N.; Kokubu, C.; Michigami, T.; Ozono, K. Wnt/Lrp/β-catenin signaling suppresses adipogenesis by inhibiting mutual activation of PPARγ and C/EBPα. Biochem. Biophys. Res. Commun. 2007, 363, 276–282. [Google Scholar] [CrossRef] [PubMed]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef]
- Tetsu, O.; McCormick, F. β-Catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Rao, M.; Bouras, T.; Wang, C.; Wu, K.; Zhang, X.; Li, Z.; Yao, T.P.; Pestell, R.G. Cyclin D1 inhibits peroxisome proliferator-activated receptor γ-mediated adipogenesis through histone deacetylase recruitment. J. Biol. Chem. 2005, 280, 16934–16941. [Google Scholar] [CrossRef]
- Freytag, S.O.; Geddes, T.J. Reciprocal regulation of adipogenesis by Myc and C/EBP α. Science 1992, 256, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. AMP-activated protein kinase: An energy sensor that regulates all aspects of cell function. Genes Dev. 2011, 25, 1895–1908. [Google Scholar] [CrossRef]
- Zang, Y.; Yu, L.F.; Nan, F.J.; Feng, L.Y.; Li, J. AMP-activated protein kinase is involved in neural stem cell growth suppression and cell cycle arrest by 5-aminoimidazole-4-carboxamide-1-beta-d-ribofuranoside and glucose deprivation by down-regulating phospho-retinoblastoma protein and cyclin D. J. Biol. Chem. 2009, 284, 6175–6184. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Lee, M.H.; Hsu, C.C.; Wei, C.L.; Tsai, Y.C. Methyl cinnamate inhibits adipocyte differentiation via activation of the CaMKK2-AMPK pathway in 3T3-L1 preadipocytes. J. Agric. Food Chem. 2012, 60, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Yuan, H.D.; Kim, D.Y.; Quan, H.Y.; Chung, S.H. Cinnamaldehyde prevents adipocyte differentiation and adipogenesis via regulation of peroxisome proliferator-activated receptor-γ (PPAR-γ) and AMP-activated protein kinase (AMPK) pathways. J. Agric. Food Chem. 2011, 59, 3666–3673. [Google Scholar] [CrossRef] [PubMed]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; Machado De Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-gamma. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Ceddia, R.B. The role of AMP-activated protein kinase in regulating white adipose tissue metabolism. Mol. Cell Endocrinol. 2013, 366, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Rahman, N.; Jeon, M.; Kim, Y.S. Delphinidin, a major anthocyanin, inhibits 3T3-L1 pre-adipocyte differentiation through activation of Wnt/β-catenin signaling. Biofactors 2016, 42, 49–59. [Google Scholar]
- Kim, M.A.; Kang, K.; Lee, H.J.; Kim, M.; Kim, C.Y.; Nho, C.W. Apigenin isolated from Daphne genkwa Siebold et Zucc. inhibits 3T3-L1 preadipocyte differentiation through a modulation of mitotic clonal expansion. Life Sci. 2014, 101, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Rhee, D.K.; Kim, B.O.; Pyo, S. Inhibitory effect of sinigrin on adipocyte differentiation in 3T3-L1 cells: Involvement of AMPK and MAPK pathways. Biomed. Pharmacother. 2018, 102, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Le, T.T.; Chen, C.; Cheng, J.X.; Kim, K.H. Curcumin inhibits adipocyte differentiation through modulation of mitotic clonal expansion. J. Nutr. Biochem. 2011, 22, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.Y.; Yang, J.Y.; Rayalam, S.; Della-Fera, M.A.; Ambati, S.; Lewis, R.D.; Hamrick, M.W.; Hartzell, D.L.; Baile, C.A. Preventing bone loss and weight gain with combinations of vitamin D and phytochemicals. J. Med. Food 2011, 14, 1352–1362. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, V.K.; Kumar, D.; Yadav, P.; Kumar, S.; Beg, M.; Shankar, K.; Varshney, S.; Rajan, S.; Srivastava, A. Curcumin-3,4-Dichloro Phenyl Pyrazole (CDPP) overcomes curcumin’s low bioavailability, inhibits adipogenesis and ameliorates dyslipidemia by activating reverse cholesterol transport. Metabolism 2017, 73, 109–124. [Google Scholar] [CrossRef]
- Abood, S.; Veisaga, M.; López, L.; Barbieri, M. Dehydroleucodine inhibits mitotic clonal expansion during adipogenesis through cell cycle arrest. Phytother. Res. 2018, 32, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, H.S.; Lee, M.S.; Cho, Y.J.; Kim, Y.S.; Hwang, J.T.; Sung, M.J.; Kim, M.S.; Kwon, D.Y. Vitisin A inhibits adipocyte differentiation through cell cycle arrest in 3T3-L1 cells. Biochem. Biophys. Res. Commun. 2008, 372, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, L.; Ran, X.; Long, M.; Zhang, M.; Tao, Y.; Luo, X.; Wang, Y.; Ma, X.; Halmurati, U.; et al. Ellagic acid reduces adipogenesis through inhibition of differentiation-prevention of the induction of Rb phosphorylation in 3T3-L1 adipocytes. Evid. Based Complement. Alternat. Med. 2013, 2013, 287534. [Google Scholar] [CrossRef] [PubMed]
- Drira, R.; Chen, S.; Sakamoto, K. Oleuropein and hydroxytyrosol inhibit adipocyte differentiation in 3T3-L1 cells. Life Sci. 2011, 89, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.; Chen, Y.Y.; Lee, P.S.; Kalyanam, N.; Ho, C.T.; Liou, W.S.; Yu, R.C.; Pan, M.H. Bisdemethoxycurcumin inhibits adipogenesis in 3T3-L1 preadipocytes and suppresses obesity in high-fat diet-fed C57BL/6 mice. J. Agric. Food Chem. 2016, 64, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Bae, E.J. Inhibition of mitotic clonal expansion mediates fisetin-exerted prevention of adipocyte differentiation in 3T3-L1 cells. Arch. Pharm. Res. 2013, 36, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Min, S.Y.; Yang, H.; Seo, S.G.; Shin, S.H.; Chung, M.Y.; Kim, J.; Lee, S.J.; Lee, H.J.; Lee, K.W. Cocoa polyphenols suppress adipogenesis in vitro and obesity in vivo by targeting insulin receptor. Int. J. Obes. 2013, 37, 584–592. [Google Scholar] [CrossRef]
- Shin, S.H.; Seo, S.G.; Min, S.; Yang, H.; Lee, E.; Son, J.E.; Kwon, J.Y.; Yue, S.; Chung, M.Y.; Kim, K.H.; et al. Caffeic acid phenethyl ester, a major component of propolis, suppresses high fat diet-induced obesity through inhibiting adipogenesis at the mitotic clonal expansion stage. J. Agric. Food Chem. 2014, 62, 4306–4312. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.Y.; Wei, L.; Castro-Munozledo, F.; Koo, W.L. (-)-Epigallocatechin-3-gallate blocks 3T3-L1 adipose conversion by inhibition of cell proliferation and suppression of adipose phenotype expression. Life Sci. 2011, 89, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, Y.J.; Choi, H.; Ko, E.H.; Kim, J.W. Reactive oxygen species facilitate adipocyte differentiation by accelerating mitotic clonal expansion. J. Biol. Chem. 2009, 284, 10601–10609. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.Y.; Seo, S.G.; Heo, Y.S.; Yue, S.; Cheng, J.X.; Lee, K.W.; Kim, K.H. Piceatannol, natural polyphenolic stilbene, inhibits adipogenesis via modulation of mitotic clonal expansion and insulin receptor-dependent insulin signaling in early phase of differentiation. J. Biol. Chem. 2012, 287, 11566–11578. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Seo, S.G.; Yue, S.; Cheng, J.X.; Lee, K.W.; Kim, K.H. An inhibitory effect of resveratrol in the mitotic clonal expansion and insulin signaling pathway in the early phase of adipogenesis. Nutr. Res. 2012, 32, 607–616. [Google Scholar] [CrossRef]
- Choi, H.S.; Jeon, H.J.; Lee, O.H.; Lee, B.Y. Dieckol, a major phlorotannin in Ecklonia cava, suppresses lipid accumulation in the adipocytes of high-fat diet-fed zebrafish and mice: Inhibition of early adipogenesis via cell-cycle arrest and AMPKα activation. Mol. Nutr. Food Res. 2015, 59, 1458–1471. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Yoon, B.K.; Park, H.; Seok, J.W.; Choi, H.; Yu, J.H.; Choi, Y.; Song, S.J.; Kim, A.; Kim, J.W. Caffeine inhibits adipogenesis through modulation of mitotic clonal expansion and the AKT/GSK3 pathway in 3T3-L1 adipocytes. BMB Rep. 2016, 49, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Mitterberger, M.C.; Zwerschke, W. Mechanisms of resveratrol-induced inhibition of clonal expansion and terminal adipogenic differentiation in 3T3-L1 preadipocytes. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1356–1376. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.Y.; Choi, K.M.; Lee, Y.S.; Yu, J.Y.; Shin, D.M.; Lee, S.; Yoo, K.S.; Lee, Y.M.; Yun, Y.P.; Yoo, H.S. Rhamnetin-induced suppression of clonal expansion during early stage of adipogenesis. Arch. Pharm. Res. 2012, 35, 1083–1089. [Google Scholar] [CrossRef]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar]
- Birsoy, K.; Berry, R.; Wang, T.; Ceyhan, O.; Tavazoie, S.; Friedman, J.M.; Rodeheffer, M.S. Analysis of gene networks in white adipose tissue development reveals a role for ETS2 in adipogenesis. Development 2011, 138, 4709–4719. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.P.; Kwon, J.; Shin, J. Berberine exerts anti-adipogenic activity through up-regulation of C/EBP inhibitors, CHOP and DEC2. Biochem. Biophys. Res. Commun. 2011, 413, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Harmon, A.W.; Patel, Y.M.; Harp, J.B. Genistein inhibits CCAAT/enhancer-binding protein beta (C/EBPbeta) activity and 3T3-L1 adipogenesis by increasing C/EBP homologous protein expression. Biochem. J. 2002, 367, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.M.; Cho, H.J.; Cho, H.; Kim, K.H.; Kim, J.B.; Park, H. Stra13/DEC1 and DEC2 inhibit sterol regulatory element binding protein-1c in a hypoxia-inducible factor-dependent mechanism. Nucleic Acids Res. 2008, 36, 6372–6385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulbagci, N.T.; Li, L.; Ling, B.; Gopinadhan, S.; Walsh, M.; Rossner, M.; Nave, K.A.; Taneja, R. SHARP1/DEC2 inhibits adipogenic differentiation by regulating the activity of C/EBP. EMBO Rep. 2009, 10, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Bordenave, N.; Ferruzzi, M.G.; Safavy, A.; Kim, K.H. Modification of curcumin with polyethylene glycol enhances the delivery of curcumin in preadipocytes and its antiadipogenic property. J. Agric. Food Chem. 2011, 59, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, E.; Lee, J.; Kim, S.; Huh, S.; Kim, Y.; Kim, Y.; Byun, S.Y.; Kim, Y.S.; Park, D. Isorhamnetin represses adipogenesis in 3T3-L1 cells. Obesity 2009, 17, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Son, H.J.; Ahn, J.; Jung, C.H.; Ha, T. Coumestrol modulates Akt and Wnt/β-catenin signaling during the attenuation of adipogenesis. Food Funct. 2016, 7, 4984–4991. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Curcumin-induced suppression of adipogenic differentiation is accompanied by activation of Wnt/β-catenin signaling. Am. J. Physiol. Cell Physiol. 2010, 298, C1510–C1516. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, Y.; Gong, Z.; Sheng, X.; Li, Z.; Zhang, W.; Qin, Y. Berberine inhibits 3T3-L1 adipocyte differentiation through the PPARγ pathway. Biochem. Biophys. Res. Commun. 2006, 348, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Kang, S.I.; Shin, H.S.; Yoon, S.A.; Kim, J.H.; Ko, H.C.; Kim, S.J. Sasa quelpaertensis Nakai extract and its constituent p-coumaric acid inhibit adipogenesis in 3T3-L1 cells through activation of the AMPK pathway. Food Chem. Toxicol. 2013, 59, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Ilavenil, S.; Kim da, H.; Srigopalram, S.; Arasu, M.V.; Lee, K.D.; Lee, J.C.; Lee, J.S.; Renganathan, S.; Choi, K.C. Potential application of p-coumaric acid on differentiation of C2C12 skeletal muscle and 3T3-L1 preadipocytes—An in Vitro and in silico approach. Molecules 2016, 21, 997. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, A.; Wu, D.; Kwan, P.; Meydani, M. Curcumin inhibits adipogenesis in 3T3-L1 adipocytes and angiogenesis and obesity in C57/BL mice. J. Nutr. 2009, 139, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Furuyashiki, T.; Nagayasu, H.; Aoki, Y.; Bessho, H.; Hashimoto, T.; Kanazawa, K.; Ashida, H. Tea catechin suppresses adipocyte differentiation accompanied by down-regulation of PPARγ2 and C/EBPα in 3T3-L1 cells. Biosci. Biotechnol. Biochem. 2004, 68, 2353–2359. [Google Scholar] [CrossRef] [PubMed]
- Harmon, A.W.; Harp, J.B. Differential effects of flavonoids on 3T3-L1 adipogenesis and lipolysis. Am. J. Physiol. Cell Physiol. 2001, 280, C807–C813. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Koch, E. Complex interactions between phytochemicals. The multi-target therapeutic concept of phytotherapy. Curr. Drug Targets 2011, 12, 122–132. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Cui, H.; Liu, G.; Zhao, X.; Li, W.; Piao, G. How can synergism of traditional medicines benefit from network pharmacology? Molecules 2017, 22, 1135. [Google Scholar] [CrossRef] [PubMed]
- Sonam, K.; Guleria, S. Synergistic antioxidant activity of natural products. Ann. Pharmacol. Pharm. 2017, 2, 1–6. [Google Scholar]
- Yang, J.-Y.; Della-Fera, M.A.; Rayalam, S.; Ambati, S.; Hartzell, D.L.; Park, H.J.; Baile, C.A. Enhanced inhibition of adipogenesis and induction of apoptosis in 3T3-L1 adipocytes with combinations of resveratrol and quercetin. Life Sci. 2008, 82, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Yang, J.Y.; Ambati, S.; Della-Fera, M.A.; Hausman, D.B.; Rayalam, S.; Baile, C.A. Combined effects of genistein, quercetin, and resveratrol in Human and 3T3-L1 Adipocytes. J. Med. Food 2008, 11, 773–783. [Google Scholar] [CrossRef]
- Kowalska, K.; Olejnik, A.; Szwajgier, D.; Olkowicz, M. Inhibitory activity of chokeberry, bilberry, raspberry and cranberry polyphenol-rich extract towards adipogenesis and oxidative stress in differentiated 3T3-L1 adipose cells. PLoS ONE 2017, 12, e0188583. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Della-Fera, M.A.; Rayalam, S.; Baile, C.A. Enhanced effects of xanthohumol plus honokiol on apoptosis in 3T3-L1 adipocytes. Obesity 2008, 16, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Rayalam, S.; Yang, J.Y.; Della-Fera, M.A.; Park, H.J.; Ambati, S.; Baile, C.A. Anti-obesity effects of xanthohumol plus guggulsterone in 3T3-L1 adipocytes. J. Med. Food 2009, 12, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Rayalam, S.; Della-Fera, M.A.; Ambati, S.; Yang, J.Y.; Park, H.J.; Baile, C.A. Enhanced effects of 1,25(OH)2D3 plus genistein on adipogenesis and apoptosis in 3T3-L1 adipocytes. Obesity 2008, 16, 539–546. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Family | Active Component | Tested Dose | Cell Proliferation | Cell Cycle | Cell-Cycle Regulators | Transcription Factor | Signaling Cascade | Intracellular Lipid Accumulation | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Alkaloids | Berberine | 0.625–10 μM | ↓PPARγ, C/EBPα mRNA and protein levels | ↓ | [129] | ||||
| Berberine | 1.5–12 μM | ↓PPARγ, C/EBPα mRNA levels ↑CHOP, DEC2 mRNA levels | ↓ | [121] | |||||
| Caffeine | 0.1–5 mM | ↓ | ↑p21, p27 protein levels ↓CDK2 protein level | ↓PPARγ, C/EBPα, C/EBPβ protein levels ↓C/EBPα, PPARα, FAS, aP2 mRNA levels ↑KLF2 mRNA level | ↓ p-AKT, p-GSK3β protein levels = p-ERK protein level | ↓ | [116] | ||
| Anthocyanins | Delphinidin | 10–150 μM | ↑ G0/G1 | ↑p27 protein level ↓CDK2, CDK6 protein levels= CDK4, Cyclin D3 protein levels ↑cyclin D1 mRNA & protein levels | ↓ C/EBPβ, C/EBPδ, C/EBPα mRNA levels ↓ PPARγ mRNA and protein levels | ↑Wnt1, Wnt10b, Fzd2, and Lrp5 mRNA levels ↑β-catenin mRNA & protein levels, ↑β-catenin nuclear translocation ↑ mRNA & nuclear translocation of β-catenin ↓GSK3β mRNA & protein levels ↑c-Myc protein levels | ↓ | [97] | |
| Asteraceae | Dehydroleu-codine | 5–12.5 μM | ↓ | ↑ G0/G1 | ↓CDK2, CDK4 protein level ↑p27 protein level | ↓C/EBPβ, PPARγ mRNA levels | ↓ p-AKT, p-Akt protein levels | ↓ | [103] |
| Coumarins | p-Coumaric acid | 125–1000 μg/mL | ↓PPARγ, C/EBPα protein levels | ↑p-AMPK | ↓ | [130] | |||
| Coumaric acid | 0.1–0.2 mM | ↓PPARγ2, C/EBPα mRNA and protein levels | ↓ | [131] | |||||
| Coumestans | Coumestrol | 20–60 μM | ↓PPARγ, C/EBPα protein levels | ↓ p-AKT, p-GSK3β ↑LRP6 protein level ↑β-catenin protein level ↑Wnt10b mRNA and protein levels ↑c-Myc, cyclin D1 mRNA and protein levels | ↓ | [127] | |||
| Phenolic acid | Curcumin | 5–20 μM | ↓ C/EBPα, PPARγ mNRA levels | ↓ | [132] | ||||
| Curcuminoids | Curcumin | 10–25 μM | ↓ C/EBPα, PPARγ protein levels | ↑Wnt10b, Fzd2, and Lrp5 mRNA levels ↑nuclear β-catenin protein ↓GSK-3β, CK1α, Axin protein levels ↑c-Myc, cyclin D1 mRNA and protein levels ↓p-ERK, p-JNK, p-p38 MAPK | ↓ | [128] | |||
| Curcumin | 5–30 μM | ↓ C/EBPβ, PPARγ, C/EBPα mRNA levels | ↓ | [125] | |||||
| Curcumin | 5–35 μM | ↓ | ↓S and/or G2/M | ↓Cyclin A, CDK2 protein levels | ↓ KLF5, C/EBPβ, PPARγ, C/EBPα mRNA levels | ↓ | [100] | ||
| Bisdemethoxycurcumin (BDMC) | 5–25 μM | ↑G0/G1 | ↓cyclin A, cyclin B protein levels ↑p21 protein level = CDK2 and 4, Cyclin D and E protein levels | ↓PPARγ, C/EBPα protein levels | ↓p-ERK1/2, p-JNK/ = p-p38 MAPK ↓p-Akt | ↓ | [107] | ||
| Curcumin-3,4-dichloro phenyl pyrazole | 5–20 uM | ↑G1, S | ↓CyclinD1, CyclinD3, CDK2, CDK4, CDK6 protein levels ↑p27 protein level | ↓PPARγ2, C/EBPα mRNA and protein levels | = Wnt3a, GATA, β-catenin, p-AMPK protein levels ↓ p-AKT, mTOR protein levels | ↓ | [102] | ||
| Flavanols | Catechin 3 -gallate (CG) | 5–30 μM | ↓ C/EBPα, PPARγ protein levels | ↓ | [133] | ||||
| Epicatechin 3-gallate (ECG) | 5–30 μM | ↓ C/EBPα, PPARγ protein levels | ↓ | [133] | |||||
| Epigallocatechin-3-gallate (EGCG) | 0.1–10 μM | ↓ | ↑G2/M | ↓ C/EBPα, PPARγ mRNA levels | ↓ | [111] | |||
| Epigallocatechin-3-gallate (EGCG) | 100 μM | ↓ | ↓G0/G1 ↑S | ↓ C/EBPα, PPARγ mRNA levels | ↓ FoxO1 mRNA level | ↓ | [66] | ||
| Flavones | Apigenin | 30–70 μM | ↓ | ↑G0/G1 | ↓CyclinD1, CDK4 protein levels ↑p27 protein level | ↓DNA-binding activity of C/EBPβ ↓C/EBPβ protein level ↑p- C/EBPβ, CHOP-10 protein levels | ↓ | [98] | |
| Flavonoids | Isorhamnetin | 1–50 μM | ↓C/EBPβ, C/EBPδ mRNA levels ↓PPARγ, C/EBPα mRNA levels | ↓ | [126] | ||||
| Flavonoids | Rhamnetin | 10–40 μM | ↓ | ↓PPARγ, C/EBPα mRNA and protein levels | ↓ | [118] | |||
| Flavonoids | Fisetin | 10–30 μM | ↓ | ↓cyclin A, cyclin D1, CDK4 protein levels ↑p27 protein level = Cyclin E, CDK2 protein levels | ↓PPARγ protein level | = p-ERK and p-AKT ↓ p-S6 (p70S6K activity) | ↓ | [108] | |
| Glucosinolates | Sinigrin (2-propenyl glucosinolate) | 1–100 μg/mL | ↑G0/G1 | ↑p21, p27 protein levels ↓CDK2 protein level | ↓p-C/EBPβ ↓PPARγ, C/EBPα protein & mRNA levels | ↓ p-ERK, p-JNK and p-p38 MAPK ↑p-AMPK | ↓ | [99] | |
| Flavonoids: Isoflavonoids | Genistein | 5–100 μM | - | - | ↓PPARγ protein level | ↓ | [134] | ||
| Genistein | 100 μM | ↓PPARγ, C/EBPα protein levels = C/EBPβ protein level ↓DNA-binding activity of C/EBPβ ↑CHOP protein level | ↓ | [122] | |||||
| Genistein | 50 μM | ↓ | ↑S | ↑cyclin A protein level = p27, p21, cyclin E, CDK2 protein levels | ↓Centromeric localization of C/EBPβ | = p-MAPK, GSK3β protein levels | ↓ | [112] | |
| Isothiocyanates | Sulforaphane | 5–20 μM | ↓ | ↑G0/G1 | ↑ p27 protein level / ↓p-Rb ↓ cyclin D1, CDK2, CDK4, cyclin A protein levels | ↓PPARγ, C/EBPα protein levels | ↓p-AKT, p-ERK | ↓ | [67] |
| Phenolic acid | Cocoa | 100–200 μg/mL | ↑G1-S | ↓PPARγ, C/EBPα mRNA and protein levels | ↓p- ERK, p-AKT, mTOR, p70S6K protein levels ↓ Insulin receptor kinase | ↓ | [109] | ||
| Phenolic acids | Caffeic acid phenethyl ester | 10–40 μM | ↑G1/S | ↓cyclin D1 mRNA and protein levels | ↓PPARγ, C/EBPα protein levels | ↓p-ERK, p-AKT | ↓ | [110] | |
| Secoiridoids | Hydroxytyrosol | 50–150 μM | ↑G0/G1 ↑S | ↓PPARγ, C/EBPα mRNA levels | ↓ | [106] | |||
| Oleuropein | 100–300 μM | ↑G0/G1 ↑S | ↓PPARγ, C/EBPα mRNA levels | ↓ | [106] | ||||
| Stilbenes | Vitisin A | 1–10 μM | ↓ | ↑G0/G1 | ↓p-Rb ↑p21 protein level | ↓PPARγ protein level and activity | = p-ERK, p-AKT | ↓ | [104] |
| Ellagic acid | 10–20 μM | ↑G0/G1 | ↓p-Rb, cyclin A protein levels | ↓C/EBPα protein level and DNA-binding activity ↓PPARγ mRNA and protein levels | ↓ | [105] | |||
| Piceatannol | 10–50 μM | NS | ↓S and G2/M | ↓PPARγ, C/EBPα protein and mRNA levels ↓C/EBPβ mRNA level | ↓ p-IR, p-IRS-1, p-AKT, p-ERK1/2 | ↓ | [113] | ||
| Resveratrol | 25–50 μM | ↓ | ↑G1/S | ↓ cyclin A, CDK2 protein levels | ↓PPARγ, C/EBPα protein levels | ↓p-AKT, p-IR ↓IR kinase activity | ↓ | [114] | |
| Resveratrol | 20 μM | ↓p-Rb, Cyclin A and D1, p21 protein levels | ↓PPARγ, C/EBPα protein level ↓PPARγ activity | ↓p-AKT, p-ERK ↑p-AMPK | ↓ | [117] | |||
| Tannins | Dieckol | 25–100 μM | ↓ | ↑G1↓S | ↓ cyclin A and D, p-Rb, CDK2 protein levels ↑p27 protein level | ↓C/EBPβ, C/EBPδ, KLF4, KLF5, ETS2 mRNA levels | ↓p- ERK, p-AKT ↑p-AMPK | ↓ | [115] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, E.; Kim, C.Y. Natural Products and Obesity: A Focus on the Regulation of Mitotic Clonal Expansion during Adipogenesis. Molecules 2019, 24, 1157. https://doi.org/10.3390/molecules24061157
Chang E, Kim CY. Natural Products and Obesity: A Focus on the Regulation of Mitotic Clonal Expansion during Adipogenesis. Molecules. 2019; 24(6):1157. https://doi.org/10.3390/molecules24061157
Chicago/Turabian StyleChang, Eugene, and Choon Young Kim. 2019. "Natural Products and Obesity: A Focus on the Regulation of Mitotic Clonal Expansion during Adipogenesis" Molecules 24, no. 6: 1157. https://doi.org/10.3390/molecules24061157
APA StyleChang, E., & Kim, C. Y. (2019). Natural Products and Obesity: A Focus on the Regulation of Mitotic Clonal Expansion during Adipogenesis. Molecules, 24(6), 1157. https://doi.org/10.3390/molecules24061157