Introduction to the Molecules Special Edition Entitled ‘Heparan Sulfate and Heparin: Challenges and Controversies’: Some Outstanding Questions in Heparan Sulfate and Heparin Research

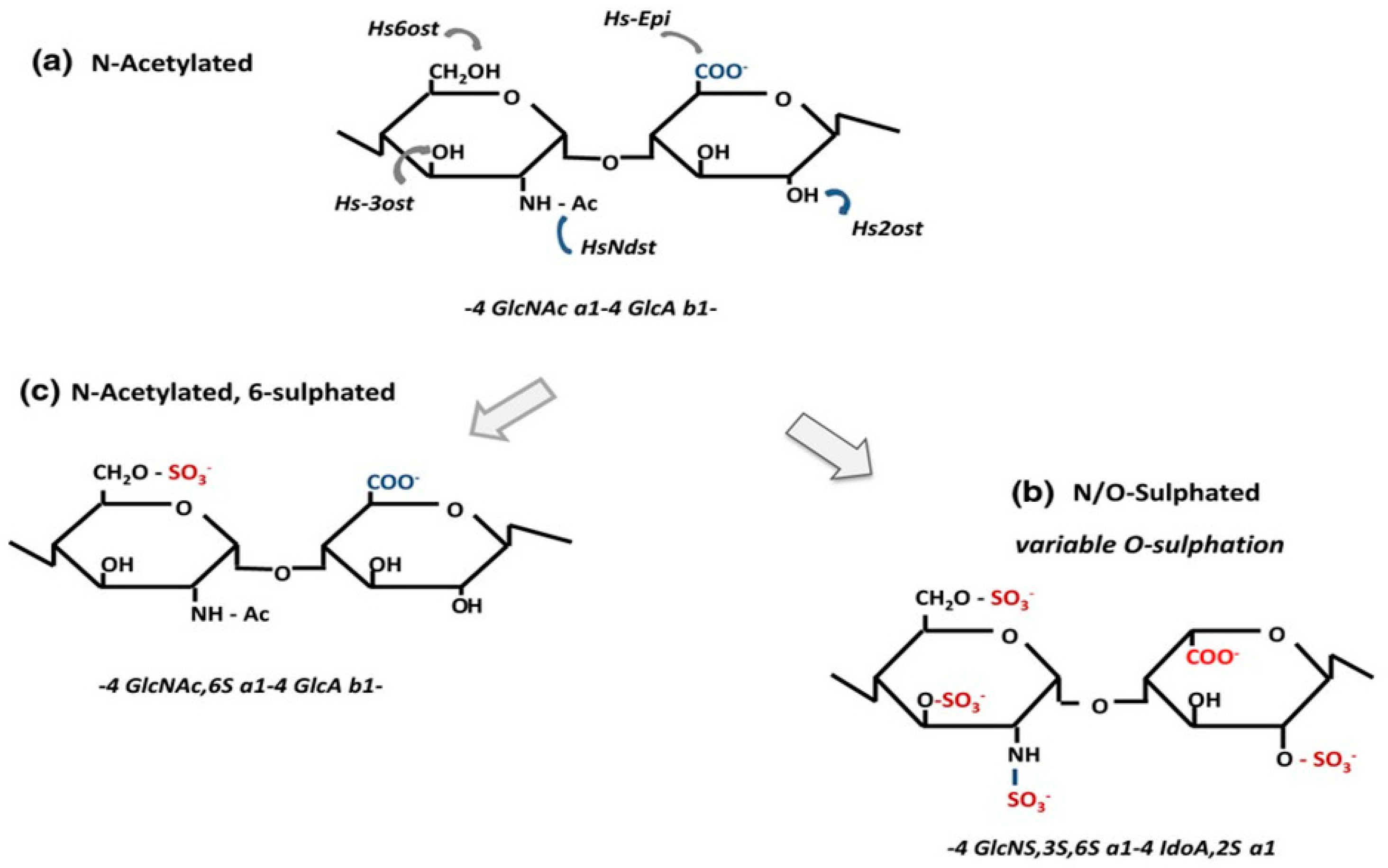{kind=link}
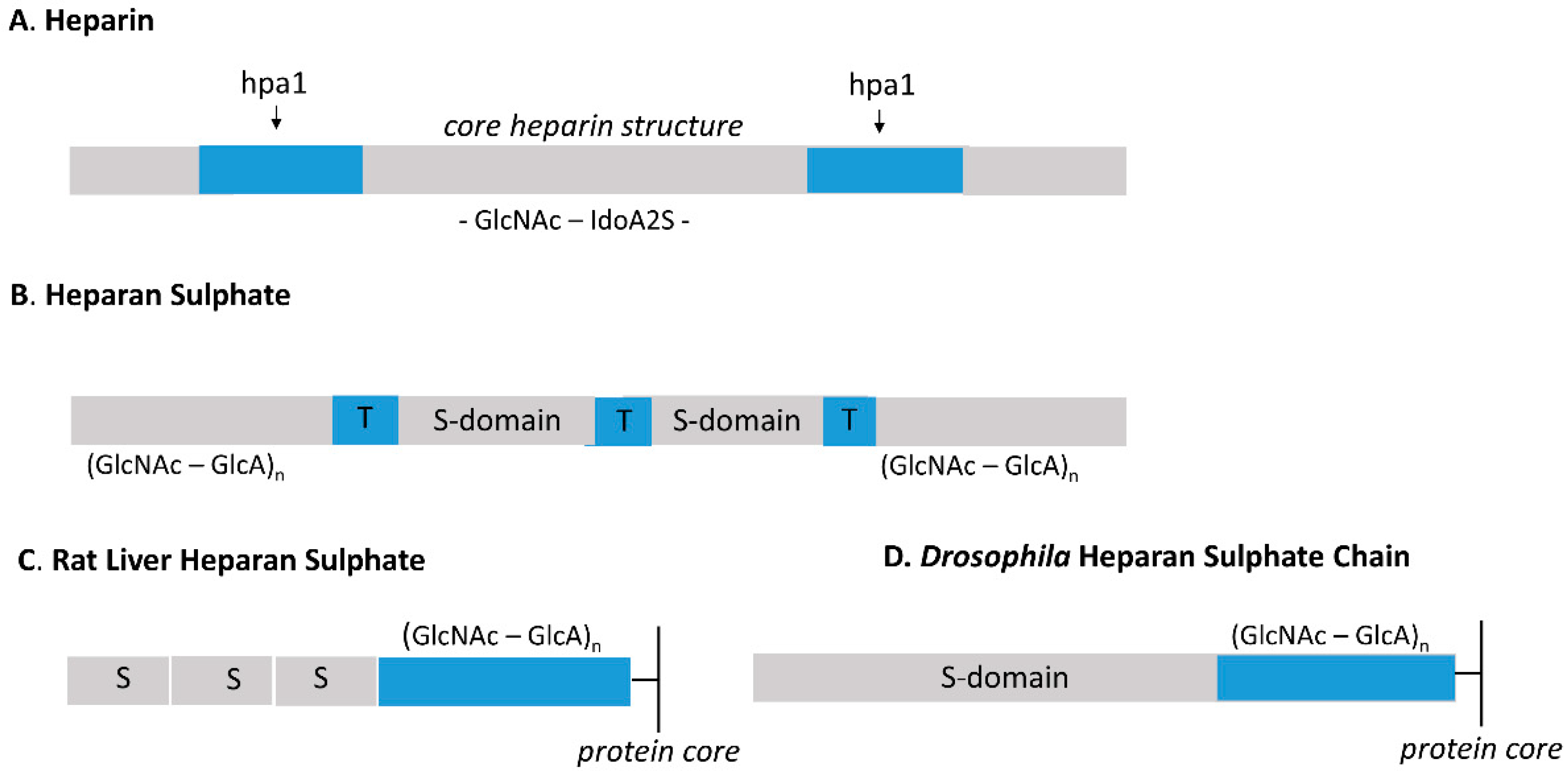{kind=link}
Funding
Conflicts of Interest
References
- Kreuger, J.; Kjellen, L. Heparan sulfate biosynthesis—Regulation and variability. J. Histochem. Cytochem. 2012, 60, 898–907. [Google Scholar] [CrossRef]
- Li, J.-P.; Kusche-Gullberg, M. Heparan sulfate: Biosynthesis, structure, and function. Int. Rev. Cell Mol. Biol. 2016, 325, 215–273. [Google Scholar] [PubMed]
- Casu, B.; Lindahl, U. Structure and biological interactions of heparin and heparan sulfate. Adv. Carbohydr. Chem. Biochem. 2001, 57, 159–206. [Google Scholar] [PubMed]
- Skidmore, M.A.; Guimond, S.E.; Rudd, T.R.; Fernig, D.G.; Turnbull, J.E.; Yates, E.A. The activities of heparan sulfate and its analogue heparin are dictated by biosynthesis, sequence, and conformation. Connect. Tissue Res. 2008, 49, 140–144. [Google Scholar] [CrossRef]
- Forster, M.; Mulloy, B. Computational approaches to the identification of heparin binding sites on the surfaces of proteins. Biochem. Soc. Trans. 2006, 34, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J. Fell-Muir Lecture: Heparan sulphate and the art of cell regulation: A polymer chain conducts the protein orchestra. Int. J. Exp. Path. 2015, 96, 203–231. [Google Scholar] [CrossRef] [PubMed]
- Ronnberg, E.; Melo, F.; Pejler, G. Mast cell proteoglycans. J. Histochem. Cytochem. 2012, 60, 950–962. [Google Scholar] [CrossRef]
- Mulloy, B. Structure and physicochemical characterization of heparin. In Heparin: A Century of Progress; Lever, R., Mulloy, B., Page, C.P., Eds.; Springer Verlag: Berlin/Heidelberg, Germany, 2012; pp. 77–98. [Google Scholar]
- Stringer, S.; Kandola, B.; Pye, D.; Gallagher, J. Heparin sequencing. Glycobiology 2003, 13, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Conrad, H.E. Heparin Binding Proteins; Academic Press: London, UK, 1997; pp. 61–107. [Google Scholar]
- Wang, A.; Sankaranarayanan, N.V.; Yanagishita, M.; Templeton, D.M.; Desai, U.R.; Sugahara, K.; Wang, C.P.; Hascall, V.C. Heparin interaction with a receptor on hyperglycemic dividing cells prevents intracellular hyaluronan synthesis and autophagy responses in models of type 1 diabetes. Matrix Biol. 2015, 48, 36–41. [Google Scholar] [CrossRef]
- Gong, F.; Jemth, P.; Escobar-Galvis, M.L.; Vlodavsky, I.; Horner, A.; Lindahl, U.; Li, J. Processing of macromolecular heparin by heparinase. J. Biol. Chem. 2003, 278, 35152–35158. [Google Scholar] [CrossRef] [PubMed]
- Radoff, S.; Danifeshefsky, I. Location on heparin of the oligosaccharide section essential for anticoagulant activity. J. Biol. Chem. 1984, 259, 166–172. [Google Scholar]
- Yamada, S.; Yamane, Y.; Tsuda, H.; Yoshida, K.; Sugahara, K. A major common trisulfated hexasaccharide core sequence, hexuronic acid(2-sulfate)-glucosamine(N-sulfate)-iduronic acid-N-acetylglucosamine-glucuronic acid-glucosamine(N-sulfate), isolated from the low sulfated irregular region of porcine intestinal heparin. J. Biol. Chem. 1998, 273, 1863–1871. [Google Scholar]
- Sugahara, K.; Tsuda, H.; Yoshida, K.; de Beer, T.; Vliegenthart, J. Structure determination of the octa- and decasaccharide sequences isolated from the carbohydrate-protein linkage region of porcine intestinal heparin. J. Biol. Chem. 1995, 270, 22914–22923. [Google Scholar] [CrossRef]
- Gallagher, J.; Walker, A. Molecular distinctions between heparan sulphate and heparin. Analysis of sulphation patterns indicates that heparan sulphate and heparin are separate families of N-sulphated polysaccharides. Biochem. J. 1985, 23, 665–674. [Google Scholar] [CrossRef]
- Turnbull, J.; Gallagher, J. Distribution of iduronate 2-sulphate residues in heparan sulphate. Evidence for an ordered polymeric structure. Biochem. J. 1991, 273, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.; Merry, C.; Lyon, M.; Thompson, J.E.; Roberts, I.S.; Gallagher, J.T. A new model for the domain structure of heparan sulfate based on the novel specificity of K5 lyase. J. Biol. Chem. 2004, 279, 27239–27245. [Google Scholar] [CrossRef]
- Merry, C.; Lyon, M.; Deakin, J.; Hopwood, J.; Gallagher, J.T. Highly sensitive sequencing of the sulphated domains of heparan sulphate. J. Biol. Chem. 1999, 274, 18455–18462. [Google Scholar] [CrossRef]
- Sanderson, P.N.; Huckerby, T.N.; Nieduszynski, I.A. Very-high-field n.m.r. studies of bovine lung heparan sulphate tetrasaccharides produced by nitrous acid deaminative cleavage. Determination of saccharide sequence, uronate composition and degrees of sulphation. Biochem. J. 1984, 223, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Zaia, J. Organ-specific heparan sulphate structural phenotypes. J. Biol. Chem. 2009, 284, 11806–11814. [Google Scholar] [CrossRef]
- Ledin, J.; Staatz, W.; Li, J.P.; Gotte, M.; Selleck, S.; Kjellen, L.; Spillman, D. Heparan sulfate structure in mice with genetically modified heparan sulfate production. J. Biol. Chem. 2004, 279, 42732–42741. [Google Scholar] [CrossRef]
- Lyon, M.; Steward, W.P.; Hampson, I.; Gallagher, J.T. Identification of an extended N-acetylated sequence adjacent to the protein-linkage region of heparan sulphate. Biochem. J. 1987, 242, 493–498. [Google Scholar] [CrossRef]
- Naimy, H.; Buczek-Thomas, J.A.; Nugent, M.; Leymarie, N.; Zaia, J. Highly sulfated nonreducing end-derived heparan sulfate domains bind fibroblast growth factor-2 with high affinity and are enriched in biologically active fractions. J. Biol. Chem. 2011, 286, 19311–19319. [Google Scholar] [CrossRef]
- Lyon, M.; Deakin, J.; Gallagher, J.T. Liver heparan sulphate structure: a novel molecular design. J. Biol. Chem. 1994, 269, 11208–11215. [Google Scholar]
- Parra, A.; Veraldi, N.; Locatelli, M.; Fini, M.; Martini, L.; Torri, G.; Sangiorgi, L.; Bisio, A. Heparin-like heparin sulfate from rabbit cartilage. Glycobiology 2012, 22, 248–257. [Google Scholar] [CrossRef]
- Stringer, S.; Mayer-Proschel, M.; Kalyani, A.; Rao, M.; Gallagher, J.T. Heparin is a unique marker of progenitors in the glial cell lineage. J. Biol. Chem. 1999, 274, 25455–25460. [Google Scholar] [CrossRef]
- Kusche-Gullberg, M.; Nybakken, K.; Perrimon, N.; Lindahl, U. Drosophila heparan sulfate, a novel design. J. Biol. Chem. 2012, 287, 21950–21956. [Google Scholar] [CrossRef]
- Rogerson, E.; Pelletier, J.; Acosta-Serrano, A.; Rose, C.; Taylor, S.; Guimond, S.; Lima, M.; Skidmore, M.; Yates, E. Variations in the peritrophic matrix composition of heparin sulfate from the tsetse fly, Glossina morsitans morsitans. Pathogens 2018, 7, 32. [Google Scholar] [CrossRef]
- Dietrich, C.P.; de Paiva, J.; Moraes, C.T.; Takahashi, H.T.; Porcionatto, M.A.; Nader, H.B. Isolation and characterization of a heparin with high anticoagulant activity from Anomalocardia brasiliana. Biochim. Biophys. Acta 1985, 843, 1–7. [Google Scholar] [CrossRef]
- Van Vactor, D.; Wall, D.P.; Johnson, K.G. Heparan sulfate proteoglycans and the emergence of neuronal connectivity. Curr. Opin. Neurobiol. 2006, 16, 40–51. [Google Scholar] [CrossRef]
- Meneghetti, M.C.; Hughes, A.J.; Rudd, T.R.; Nader, H.B.; Powell, A.K.; Yates, E.A.; Lima, M.A. Heparan sulfate and heparin interactions with proteins. J. R. Soc. Interface 2015, 12, 0589. [Google Scholar] [CrossRef]
- Ori, A.; Wilkinson, M.C.; Fernig, D.G. The hepanosome and regulation of cell functions: Structures, functions and challenges. Front. Biosci. 2008, 13, 4309–4338. [Google Scholar] [CrossRef]
- Mizumoto, M.; Kitagawa, H.; Sugahara, K. Biosynthesis of heparin and heparin sulfate. In Chemistry and Biology of Heparin and Heparan Sulfate; Garg, H.G., Linhardt, R.J., Hales, C.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 203–244. [Google Scholar]
- Rudd, T.R.; Yates, E.A. A highly efficient tree structure for the biosynthesis of heparin sulfate accounts for the commonly observed disaccharides and suggests a mechanism for domain synthesis. Mol. Biosyst. 2012, 8, 1499–1506. [Google Scholar] [CrossRef]
- Meneghetti, M.C.Z.; Ferreira, T.G.; Tashima, A.K.; Chavante, S.F.; Yates, E.A.; Liu, J.; Nader, H.B.; Lima, M.A. Insights into the role of 3-O-sulfotransferases in heparan sulfate biosynthesis. Org. Biomol. Chem. 2017, 15, 6792–6799. [Google Scholar] [CrossRef]
- Zhang, X.; Pagadala, V.; Jester, H.M.; Lim, A.M.; Pham, T.Q.; Goulas, A.M.P.; Liu, J.; Linhardt, R.J. Chemoenzymatic synthesis of heparan sulfate and heparin oligosaccharides: paving the way to a diverse library for glycobiologists. Chem. Sci. 2017, 8, 7932–7940. [Google Scholar] [CrossRef]
- Rudd, T.R.; Guimond, S.E.; Skidmore, M.A.; Duchesne, L.; Guerrini, M.; Torri, G.; Cosentino, C.; Brown, A.; Clarke, D.T.; Turnbull, J.E.; et al. Influence of substitution pattern and cation binding on conformation and activity in heparin derivatives. Glycobiology 2007, 17, 983–993. [Google Scholar] [CrossRef] [Green Version]
- Guimond, S.E.; Rudd, T.R.; Skidmore, M.A.; Ori, A.; Gaudesi, D.; Cosentino, C.; Guerrini, M.; Edge, R.; Collison, D.; McInnes, E.; et al. Cations modulate polysaccharide structure to determine FGF-FGFR signaling: A comparison of signaling and inhibitory polysaccharide interactions with FGF-1 in solution. Biochemistry 2009, 48, 4772–4779. [Google Scholar] [CrossRef]
- Rudd, T.R.; Yates, E.A. Conformational degeneracy restricts the effective information content of heparan sulfate. Mol. Biosyst. 2010, 6, 902–908. [Google Scholar] [CrossRef]
- Hughes, A.; Meneghetti, M.; Hung, T.-Y.; Huang, S.-C.; Elli, S.; Guerrini, M.; Rudd, T.; Lima, M.; Yates, E. Investigating the relationship between temperature, conformation and Ca binding in heparin model oligosaccharides. Carbohydr. Res. 2017, 390, 56–66. [Google Scholar]
- Akhtar, F.; Wan, X.; Wu, G.; Kesse, S.; Wang, S.; He, S. Low molecular weight heparins: Reduced size particulate systems for improved therapeutic outcomes. Molecules 2018, 23, 1757. [Google Scholar] [CrossRef]
- Hricovini, M.; Hricovini, M. Solution conformation of heparin tetrasaccharide. DFT analysis of structure and spin-spin coupling constants. Molecules 2018, 23, 3042. [Google Scholar] [CrossRef]
- Veraldi, N.; Parra, A.; Urso, E.; Cosentino, C.; Locatelli, M.; Corsini, S.; Pedrini, E.; Naggi, A.; Bisio, A.; Sangiorgi, L. Structural features of heparin sulfate from multiple osteochondromas and chondrosarcomas. Molecules 2018, 23, 3277. [Google Scholar] [CrossRef]
- Ponert, J.M.; Gockel, L.M.; Henze, S.; Schlesinger, M. Unfractionated and low molecular weight heparin reduce platelet induced endothelial-mesenchymal transition in pancreatic and prostate cancer cells. Molecules 2018, 23, 2690. [Google Scholar] [CrossRef]
- Gockel, L.M.; Ponert, J.M.; Schwarz, S.; Schlesinger, M.; Bendas, G. The low molecular weight heparin tinzaparin attenuates platelet activation in terms of metastatic niche formation. Molecules 2018, 23, 2753. [Google Scholar] [CrossRef]
- Hellec, C.; Diawara, M.; Carpentier, M.; Denys, A.; Allain, F. The pro-tumoral activity of heparan sulfate 3-O-sulfotransferase 3B (HS3ST3B) in breast cancer MDA-MB-231 cells is dependent on the expression of neuropilin-1. Molecules 2018, 23, 2718. [Google Scholar] [CrossRef]
- Lanzi, C.; Cassinelli, G. Heparan sulfate mimetics in cancer therapy. Molecules 2018, 23, 2915. [Google Scholar] [CrossRef]
- Chhabra, M.; Ferro, V. The development of assays for heparanase enzymatic activity: Towards a gold standard. Molecules 2018, 23, 2971. [Google Scholar] [CrossRef]
- Boittier, E.D.; Gandi, N.S.; Ferro, V.; Coombe, D.R. Cross-species analysis of GAG binding proteins reveals some animal models are ‘more equal than others’. Molecules 2019, 24, 924. [Google Scholar] [CrossRef]
- Xu, R.; Ori, A.; Rudd, T.R.; Uniewicz, K.A.; Ahmed, Y.A.; Guimond, S.E.; Skidmore, M.A.; Siligardi, G.; Yates, E.A.; Fernig, D.G. Diversification of the structural determinants of fibroblast growth factor-heparin interactions: Implications for binding specificity. J. Biol. Chem. 2012, 287, 40061–40073. [Google Scholar] [CrossRef]
- Li, Y.; Sun, C.; Yates, E.A.; Jiang, C.; Wilkinson, M.C.; Fernig, D.G. Heparin binding preference and structures in the fibroblast growth factor family parallel their evolutionary diversification. Open Biol. 2016, 6, 150275. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Rudd, T.R.; Hughes, A.J.; Siligardi, G.; Fernig, D.G.; Yates, E.A. Analysis of the fibroblast growth factor receptor (FGFR) signaling network with heparin as coreceptor: Evidence for the expansion of the core FGFR signalling network. FEBS J. 2013, 280, 2260–2270. [Google Scholar] [CrossRef]
- Veraldi, N.; Hughes, A.J.; Rudd, T.R.; Thomas, H.B.; Edwards, S.W.; Hadfield, L.; Skidmore, M.A.; Siligardi, G.; Cosentino, C.; Shute, J.K.; et al. Heparin derivatives for the targeting of multiple activities in the inflammatory response. Carbohydr. Polym. 2015, 117, 400–407. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yates, E.A.; Gallagher, J.T.; Guerrini, M. Introduction to the Molecules Special Edition Entitled ‘Heparan Sulfate and Heparin: Challenges and Controversies’: Some Outstanding Questions in Heparan Sulfate and Heparin Research. Molecules 2019, 24, 1399. https://doi.org/10.3390/molecules24071399
Yates EA, Gallagher JT, Guerrini M. Introduction to the Molecules Special Edition Entitled ‘Heparan Sulfate and Heparin: Challenges and Controversies’: Some Outstanding Questions in Heparan Sulfate and Heparin Research. Molecules. 2019; 24(7):1399. https://doi.org/10.3390/molecules24071399
Chicago/Turabian StyleYates, Edwin A, John T Gallagher, and Marco Guerrini. 2019. "Introduction to the Molecules Special Edition Entitled ‘Heparan Sulfate and Heparin: Challenges and Controversies’: Some Outstanding Questions in Heparan Sulfate and Heparin Research" Molecules 24, no. 7: 1399. https://doi.org/10.3390/molecules24071399
APA StyleYates, E. A., Gallagher, J. T., & Guerrini, M. (2019). Introduction to the Molecules Special Edition Entitled ‘Heparan Sulfate and Heparin: Challenges and Controversies’: Some Outstanding Questions in Heparan Sulfate and Heparin Research. Molecules, 24(7), 1399. https://doi.org/10.3390/molecules24071399