
Antiviral Activities of Silymarin and Derivatives

Abstract
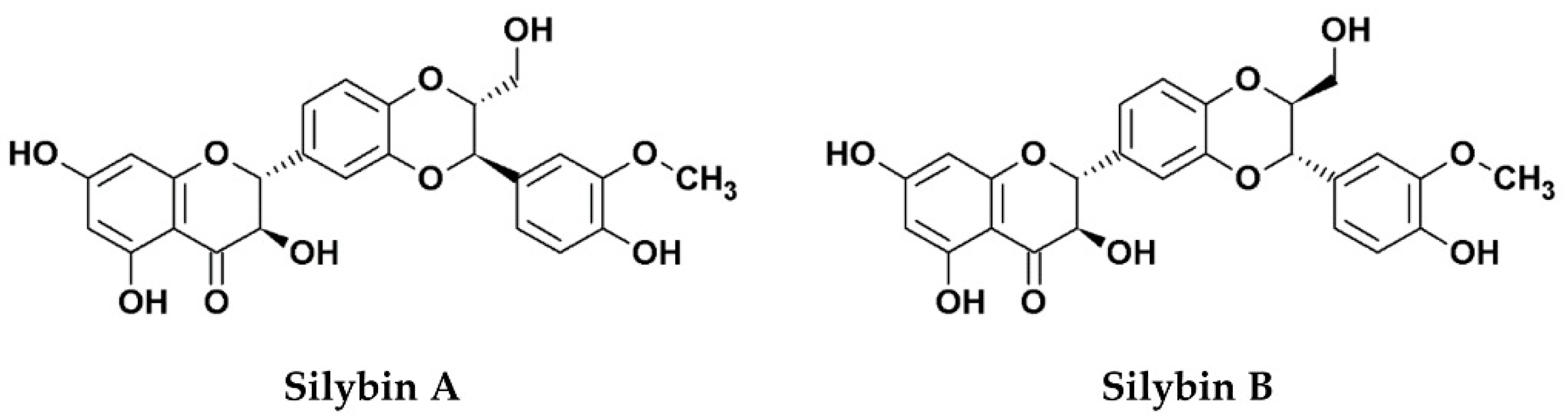
:1. Silymarin, Its Components, and Derivatives
2. Antiviral Activity of Silymarin and Its Derivatives In Vitro, In Silico, and In Vivo
2.1. The Flaviviridae Family
2.1.1. Hepatitis C Virus
2.1.2. Dengue Virus
2.2. Influenza A Virus
2.3. Human Immunodeficiency Virus
2.4. The Togaviridae Family
2.5. Hepatitis B Virus
3. Antiviral Activity of Silymarin and Its Derivatives in Clinical Trials
3.1. Chronic Hepatitis C
3.2. Liver Transplantation in Hepatitis C
3.3. HIV/HCV Coinfection
4. Challenges to Clinical Application and the Need to Enhance Bioavailability
5. Prospects of Silymarin and Derivatives in Antiviral Development
Author Contributions
Funding
Conflicts of Interest
References
- Federico, A.; Dallio, M.; Loguercio, C.; Tsai, T.-H.; Jeon, Y. Silymarin/Silybin and Chronic Liver Disease: A Marriage of Many Years. Molecules 2017, 22, 191. [Google Scholar] [CrossRef]
- Dixit, N.; Kohli, K.; Ahmad, S.; Baboota, S.; Ali, J. Silymarin: A review of pharmacological aspects and bioavailability enhancement approaches. Indian J. Pharmacol. 2007, 39, 172. [Google Scholar] [CrossRef]
- Bijak, M. Silybin, a Major Bioactive Component of Milk Thistle (Silybum marianum L. Gaernt.)-Chemistry, Bioavailability, and Metabolism. Molecules 2017, 22, 1942. [Google Scholar] [CrossRef] [PubMed]
- Strader, D.B.; Bacon, B.R.; Lindsay, K.L.; Brecque, D.R.; Morgan, T.; Wright, E.C.; Allen, J.; Khokar, M.F.; Hoofnagle, J.H.; Seeff, L.B.; et al. Use of complementary and alternative medicine in patients with liver disease. Am. J. Gastroenterol. 2002, 97, 2391–2397. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. Silibinin-C-2′,3-dihydrogensuccinate, Disodium Salt for the Prevention of Recurrent Hepatitis C in Liver Transplant Recipients. Available online: https://www.ema.europa.eu/en/medicines/human/orphan-designations/eu310828 (accessed on 23 March 2019).
- Alter, H.J.; Seeff, L.B. Recovery, persistence, and sequelae in hepatitis C virus infection: A perspective on long-term outcome. Semin. Liver Dis. 2000, 20, 17–35. [Google Scholar]
- Franco, L.; Palacios, G.; Martinez, J.A.; Vázquez, A.; Savji, N.; De Ory, F.; Sanchez-Seco, M.P.; Martín, D.; Lipkin, W.I.; Tenorio, A. First report of sylvatic DENV-2-associated dengue hemorrhagic fever in West Africa. PLoS Negl. Trop. Dis. 2011, 5, e1251. [Google Scholar] [CrossRef] [PubMed]
- Polyak, S.J.; Morishima, C.; Shuhart, M.C.; Wang, C.C.; Liu, Y.; Lee, D.Y. Inhibition of T-Cell Inflammatory Cytokines, Hepatocyte NF-κB Signaling, and HCV Infection by Standardized Silymarin. Gastroenterology 2007, 132, 1925–1936. [Google Scholar] [CrossRef]
- Morishima, C.; Lohmann, V.; Pal, S.; Liu, Y.; Polyak, S.J.; Lee, D.Y.W.; Graf, T.N.; Oberlies, N. Identification of hepatoprotective flavonolignans from silymarin. Proc. Natl. Acad. Sci. USA 2010, 107, 5995–5999. [Google Scholar]
- Wagoner, J.; Negash, A.; Kane, O.J.; Martinez, L.E.; Nahmias, Y.; Bourne, N.; Owen, D.M.; Grove, J.; Brimacombe, C.; McKeating, J.A.; et al. Multiple Effects of Silymarin on the Hepatitis C Virus Lifecycle. Hepatology 2010, 51, 1912–1921. [Google Scholar] [CrossRef] [Green Version]
- Ahmed–Belkacem, A.; Ahnou, N.; Barbotte, L.; Wychowski, C.; Pallier, C.; Brillet, R.; Pohl, R.; Pawlotsky, J. Silibinin and Related Compounds Are Direct Inhibitors of Hepatitis C Virus RNA-Dependent RNA Polymerase. Gastroenterology 2010, 138, 1112–1122. [Google Scholar] [CrossRef] [PubMed]
- Blaising, J.; Lévy, P.L.; Gondeau, C.; Phelip, C.; Varbanov, M.; Teissier, E.; Ruggiero, F.; Polyak, S.J.; Oberlies, N.H.; Ivanovic, T.; et al. Silibinin inhibits hepatitis C virus entry into hepatocytes by hindering clathrin-dependent trafficking. Cell. Microbiol. 2013, 15, 1866–1882. [Google Scholar] [CrossRef]
- Esser-Nobis, K.; Romero-Brey, I.; Ganten, T.M.; Gouttenoire, J.; Harak, C.; Klein, R.; Schemmer, P.; Binder, M.; Schnitzler, P.; Moradpour, D.; et al. Analysis of hepatitis C virus resistance to Silibinin in vitro and in vivo points to a novel mechanism involving nonstructural protein 4B. Hepatology 2013, 57, 953–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosert, R.; Egger, D.; Lohmann, V.; Bartenschlager, R.; Blum, H.E.; Bienz, K.; Moradpour, D. Identification of the Hepatitis C Virus RNA Replication Complex in Huh-7 Cells Harboring Subgenomic Replicons. J. Virol. 2003, 77, 5487–5492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.H.; Lin, C.C.; Hsu, W.C.; Chung, C.Y.; Lin, C.C.; Jassey, A.; Chang, S.P.; Tai, C.J.; Tai, C.J.; Shields, J.; et al. Highly bioavailable silibinin nanoparticles inhibit HCV infection. Gut 2017, 66, 1853–1861. [Google Scholar] [CrossRef]
- DebRoy, S.; Hiraga, N.; Imamura, M.; Hayes, C.N.; Akamatsu, S.; Canini, L.; Perelson, A.S.; Pohl, R.T.; Persiani, S.; Uprichard, S.L. Hepatitis C virus dynamics and cellular gene expression in uPA-SCID chimeric mice with humanized livers during intravenous silibinin monotherapy. J. Viral Hepat. 2016, 23, 708–717. [Google Scholar] [CrossRef]
- Wagoner, J.; Morishima, C.; Graf, T.N.; Oberlies, N.H.; Teissier, E.; Pécheur, E.-I.; Tavis, J.E.; Polyak, S.J. Differential In Vitro Effects of Intravenous versus Oral Formulations of Silibinin on the HCV Life Cycle and Inflammation. PLoS ONE 2011, 6, e16464. [Google Scholar] [CrossRef] [PubMed]
- Qaddir, I.; Rasool, N.; Hussain, W.; Mahmood, S. Computer-aided analysis of phytochemicals as potential dengue virus inhibitors based on molecular docking, ADMET and DFT studies. J. Vector Borne Dis. 2017, 54, 255. [Google Scholar]
- Webster, R.G.; Govorkova, E.A. Continuing challenges in influenza. Ann. N. Y. Acad. Sci. 2014, 1323, 115–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, F.G.; Sugaya, N.; Hirotsu, N.; Lee, N.; De Jong, M.D.; Hurt, A.C.; Ishida, T.; Sekino, H.; Yamada, K.; Portsmouth, S.; et al. Baloxavir Marboxil for Uncomplicated Influenza in Adults and Adolescents. N. Engl. J. Med. 2018, 379, 913–923. [Google Scholar] [CrossRef]
- Gažák, R.; Purchartová, K.; Marhol, P.; Živná, L.; Sedmera, P.; Valentova, K.; Kato, N.; Matsumura, H.; Kaihatsu, K.; Křen, V. Antioxidant and antiviral activities of silybin fatty acid conjugates. Eur. J. Med. Chem. 2010, 45, 1059–1067. [Google Scholar] [CrossRef]
- Song, J.; Choi, H. Silymarin efficacy against influenza A virus replication. Phytomedicine 2011, 18, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.-P.; Wu, L.-Q.; Li, R.; Zhao, X.-F.; Wan, Q.-Y.; Chen, X.-X.; Li, W.-Z.; Wang, G.-F.; Li, K.-S. Identification of 23-(S)-2-Amino-3-Phenylpropanoyl-Silybin as an Antiviral Agent for Influenza A Virus Infection In Vitro and In Vivo. Antimicrob. Agents Chemother. 2013, 57, 4433–4443. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. Tissue distribution of silibinin, the major active constituent of silymarin, in mice and its association with enhancement of phase II enzymes: Implications in cancer chemoprevention. Carcinogenesis 1999, 20, 2101–2108. [Google Scholar] [CrossRef] [PubMed]
- Roe, B.; Hall, W.W. Cellular and molecular interactions in coinfection with hepatitis C virus and human immunodeficiency virus. Expert Rev. Mol. Med. 2008, 10, 10. [Google Scholar] [CrossRef]
- McClure, J.; Lovelace, E.S.; Elahi, S.; Maurice, N.J.; Wagoner, J.; Dragavon, J.; Mittler, J.E.; Kraft, Z.; Stamatatos, L.; Horton, H.; et al. Correction: Silibinin Inhibits HIV-1 Infection by Reducing Cellular Activation and Proliferation. PLoS ONE 2012, 7, 41832. [Google Scholar] [CrossRef]
- McClure, J.; Margineantu, D.H.; Sweet, I.R.; Polyak, S.J. Inhibition of HIV by Legalon-SIL is independent of its effect on cellular metabolism. Virology 2014, 449, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Lani, R.; Hassandarvish, P.; Chiam, C.W.; Moghaddam, E.; Chu, J.J.H.; Rausalu, K.; Merits, A.; Higgs, S.; VanLandingham, D.; Abu Bakar, S.; et al. Antiviral activity of silymarin against chikungunya virus. Sci. Rep. 2015, 5, 11421. [Google Scholar] [CrossRef] [Green Version]
- Camini, F.C.; Da Silva, T.F.; Caetano, C.C.D.S.; Almeida, L.T.; Ferraz, A.C.; Vitoreti, V.M.A.; Silva, B.D.M.; Silva, S.D.Q.; De Magalhães, J.C.; Magalhães, C.L.D.B. Antiviral activity of silymarin against Mayaro virus and protective effect in virus-induced oxidative stress. Antivir. Res. 2018, 158, 8–12. [Google Scholar] [CrossRef]
- Hyun, S.; Lee, S.; Ventura, W.R.; McMenamin, J. Knowledge, Awareness, and Prevention of Hepatitis B Virus Infection Among Korean American Parents. J. Immigr. Minor. Heal. 2017, 20, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umetsu, T.; Inoue, J.; Kogure, T.; Kakazu, E.; Ninomiya, M.; Iwata, T.; Takai, S.; Nakamura, T.; Sano, A.; Shimosegawa, T. Inhibitory effect of silibinin on hepatitis B virus entry. Biochem. Biophys. Rep. 2018, 14, 20–25. [Google Scholar] [CrossRef]
- Wu, Y.-F.; Fu, S.-L.; Kao, C.-H.; Yang, C.-W.; Lin, C.-H.; Hsu, M.-T.; Tsai, T.-F. Chemopreventive Effect of Silymarin on Liver Pathology in HBV X Protein Transgenic Mice. Cancer Res. 2008, 68, 2033–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanamly, M.; Tadros, F.; Labeeb, S.; Makld, H.; Mikhail, N.; Abdel-Hamid, M.; Shehata, M.; Abu-Baki, L.; Medhat, A.; Magder, L.; et al. Randomised double-blinded trial evaluating silymarin for chronic hepatitis C in an Egyptian village: Study description and 12-month results. Dig. Liver Dis. 2004, 36, 752–759. [Google Scholar] [CrossRef]
- Gabbay, E.; Zigmond, E.; Pappo, O.; Hemed, N.; Rowe, M.; Zabrecky, G.; Cohen, R.; Ilan, Y. Antioxidant therapy for chronic hepatitis C after failure of interferon: Results of phase II randomized, double-blind placebo controlled clinical trial. World J. Gastroenterol. 2007, 13, 5317–5323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fried, M.W.; Navarro, V.J.; Afdhal, N.; Belle, S.H.; Wahed, A.S.; Hawke, R.L.; Doo, E.; Meyers, C.M.; Reddy, K.R.; Silymarin, N.; et al. Effect of silymarin (milk thistle) on liver disease in patients with chronic hepatitis C unsuccessfully treated with interferon therapy: A randomized controlled trial. JAMA 2012, 308, 274–282. [Google Scholar] [CrossRef]
- Malaguarnera, M.; Motta, M.; Vacante, M.; Malaguarnera, G.; Caraci, F.; Nunnari, G.; Gagliano, C.; Greco, C.; Chisari, G.; Drago, F.; et al. Silybin-vitamin E-phospholipids complex reduces liver fibrosis in patients with chronic hepatitis C treated with pegylated interferon alpha and ribavirin. Am. J. Transl. Res. 2015, 7, 2510–2518. [Google Scholar]
- Malaguarnera, G.; Bertino, G.; Chisari, G.; Motta, M.; Vecchio, M.; Vacante, M.; Caraci, F.; Greco, C.; Drago, F.; Nunnari, G.; et al. Silybin supplementation during HCV therapy with pegylated interferon-α plus ribavirin reduces depression and anxiety and increases work ability. BMC Psychiatry 2016, 16, 398. [Google Scholar] [CrossRef]
- Loguercio, C.; Festi, D.; Loguercio, C. Silybin and the liver: From basic research to clinical practice. World J. Gastroenterol. 2011, 17, 2288–2301. [Google Scholar] [CrossRef] [PubMed]
- Ferenci, P.; Scherzer, T.M.; Kerschner, H.; Rutter, K.; Beinhardt, S.; Hofer, H.; Schöniger–Hekele, M.; Holzmann, H.; Steindl–Munda, P. Silibinin is a potent antiviral agent in patients with chronic hepatitis C not responding to pegylated interferon/ribavirin therapy. Gastroenterology 2008, 135, 1561–1567. [Google Scholar] [CrossRef]
- Guedj, J.; Dahari, H.; Pohl, R.-T.; Ferenci, P.; Perelson, A.S. Understanding silibinin’s modes of action against HCV using viral kinetic modeling. J. Hepatol. 2012, 56, 1019–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biermer, M.; Berg, T. Rapid Suppression of Hepatitis C Viremia Induced by Intravenous Silibinin Plus Ribavirin. Gastroenterology 2009, 137, 390–391. [Google Scholar] [CrossRef] [PubMed]
- Rutter, K.; Scherzer, T.-M.; Beinhardt, S.; Kerschner, H.; Stättermayer, A.F.; Hofer, H.; Popow-Kraupp, T.; Steindl-Munda, P.; Ferenci, P. Intravenous silibinin as ‘rescue treatment’ for on-treatment non-responders to pegylated interferon/ribavirin combination therapy. Antivir. Ther. 2011, 16, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Biermer, M.; Schlosser, B.; Fülöp, B.; van Bömmel, F.; Brodzinski, A.; Heyne, R.; Keller, K.; Sarrazin, C.; Berg, T. High-dose silibinin rescue treatment for HCV-infected patients showing suboptimal virologic response to standard combination therapy. J. Viral Hepat. 2012, 19, 547–553. [Google Scholar] [CrossRef]
- Dahari, H.; Shteingart, S.; Gafanovich, I.; Cotler, S.J.; D’Amato, M.; Pohl, R.T.; Weiss, G.; Ashkenazi, Y.J.; Tichler, T.; Goldin, E.; et al. Sustained virological response with intravenous silibinin: Individualized IFN-free therapy via real-time modelling of HCV kinetics. Liver Int. 2015, 35, 289–294. [Google Scholar] [CrossRef]
- Verna, E.C.; Brown, R.S., Jr. Hepatitis C Virus Infection in Liver Transplant Candidates and Recipients. Available online: https://www.uptodate.com/contents/hepatitis-c-virus-infection-in-liver-transplant-candidates-and-recipients (accessed on 10 March 2019).
- Neumann, U.; Biermer, M.; Eurich, D.; Neuhaus, P.; Berg, T. Successful prevention of hepatitis C virus (HCV) liver graft reinfection by silibinin mono-therapy. J. Hepatol. 2010, 52, 951–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beinhardt, S.; Rasoul-Rockenschaub, S.; Scherzer, T.M.; Ferenci, P. Silibinin monotherapy prevents graft infection after orthotopic liver transplantation in a patient with chronic hepatitis C. J. Hepatol. 2011, 54, 591–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eurich, D.; Bahra, M.; Berg, T.; Boas-Knoop, S.; Biermer, M.; Neuhaus, R.; Neuhaus, P.; Neumann, U. Treatment of hepatitis C-virus-reinfection after liver transplant with silibinin in nonresponders to pegylated interferon-based therapy. Exp. Clin. Transplant. 2011, 9, 1–6. [Google Scholar] [PubMed]
- Aghemo, A.; Bhoori, S.; De Nicola, S.; Mazzaferro, V.; Colombo, M. Failure of Intravenous Silibinin Monotherapy to Prevent Hepatitis C Genotype 2A Liver Graft Reinfection. Hepat. Mon. 2012, 12, 411–414. [Google Scholar] [CrossRef] [Green Version]
- Knapstein, J.; Wörns, A.M.; Galle, P.R.; Zimmermann, T. Combination therapy with silibinin, pegylated interferon and ribavirin in a patient with hepatitis C virus genotype 3 reinfection after liver transplantation: A case report. J. Med. Case Rep. 2014, 8, 257. [Google Scholar] [CrossRef]
- Mariño, Z.; Crespo, G.; D’Amato, M.; Brambilla, N.; Giacovelli, G.; Rovati, L.; Costa, J.; Navasa, M.; Forns, X. Intravenous silibinin monotherapy shows significant antiviral activity in HCV-infected patients in the peri-transplantation period. J. Hepatol. 2013, 58, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Rendina, M.; D’Amato, M.; Castellaneta, A.; Castellaneta, N.M.; Brambilla, N.; Giacovelli, G.; Rovati, L.; Rizzi, S.F.; Zappimbulso, M.; Bringiotti, R.S.; et al. Antiviral activity and safety profile of silibinin in HCV patients with advanced fibrosis after liver transplantation: A randomized clinical trial. Transpl. Int. 2014, 27, 696–704. [Google Scholar] [CrossRef]
- Bárcena, R.; Moreno, A.; Rodriguez-Gandia, M.A.; Albillos, A.; Arocena, C.; Blesa, C.; García-Hoz, F.; Graus, J.; Nuño, J.; López-Hervás, P.; et al. Safety and anti-HCV effect of prolonged intravenous silibinin in HCV genotype 1 subjects in the immediate liver transplant period. J. Hepatol. 2013, 58, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Maier, I.; Wu, G.Y.; Maier, G.Y.W.I. Hepatitis C and HIV co-infection: A review. World J. Gastroenterol. 2002, 8, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Meissner, E.G. Update in HIV/HCV Co-Infection in the Direct Acting Antiviral Era. Curr. Opin. Gastroenterol. 2017, 33, 120–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payer, B.; Reiberger, T.; Rutter, K.; Beinhardt, S.; Staettermayer, A.; Peck-Radosavljevic, M.; Ferenci, P. Successful HCV eradication and inhibition of HIV replication by intravenous silibinin in an HIV–HCV coinfected patient. J. Clin. Virol. 2010, 49, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Braun, D.; Rauch, A.; Durisch, N.; Eberhard, N.; Anagnostopoulos, A.; Ledergerber, B.; Metzner, K.J.; Böni, J.; Weber, R.; Fehr, J. Efficacy of lead-in silibinin and subsequent triple therapy in difficult-to-treat HIV/hepatitis C virus-coinfected patients. HIV Med. 2014, 15, 625–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, D.L.; Rauch, A.; Aouri, M.; Durisch, N.; Eberhard, N.; Anagnostopoulos, A.; Ledergerber, B.; Müllhaupt, B.; Metzner, K.J.; Decosterd, L.; et al. A Lead-In with Silibinin Prior to Triple-Therapy Translates into Favorable Treatment Outcomes in Difficult-To-Treat HIV/Hepatitis C Coinfected Patients. PLoS ONE 2015, 10, e0133028. [Google Scholar] [CrossRef]
- Woo, J.S.; Kim, T.-S.; Park, J.-H.; Chi, S.-C. Formulation and biopharmaceutical evaluation of silymarin using SMEDDS. Arch. Pharmacal. Res. 2007, 30, 82–89. [Google Scholar] [CrossRef]
- Wu, J.-W.; Lin, L.-C.; Hung, S.-C.; Chi, C.-W.; Tsai, T.-H. Analysis of silibinin in rat plasma and bile for hepatobiliary excretion and oral bioavailability application. J. Pharm. Biomed. Anal. 2007, 45, 635–641. [Google Scholar] [CrossRef]
- Wu, J.-W.; Lin, L.-C.; Tsai, T.-H. Drug–drug interactions of silymarin on the perspective of pharmacokinetics. J. Ethnopharmacol. 2009, 121, 185–193. [Google Scholar] [CrossRef]
- Weyhenmeyer, R.; Mascher, H.; Birkmayer, J. Study on dose-linearity of the pharmacokinetics of silibinin diastereomers using a new stereospecific assay. Int. J. Clin. Pharmacol. Ther. Toxicol. 1992, 30, 134–138. [Google Scholar]
- Kidd, P.; Head, K. A review of the bioavailability and clinical efficacy of milk thistle phytosome: A silybin-phosphatidylcholine complex (Siliphos). Altern. Med. A J. Clin. Ther. 2005, 10, 193–203. [Google Scholar]
- Voinovich, D.; Perissutti, B.; Grassi, M.; Passerini, N.; Bigotto, A. Solid State Mechanochemical Activation of Silybum marianum Dry Extract with Betacyclodextrins: Characterization and Bioavailability of the Coground Systems. J. Pharm. Sci. 2009, 98, 4119–4129. [Google Scholar] [CrossRef]
- Kosina, P.; Kren, V.; Gebhardt, R.; Grambal, F.; Ulrichova, J.; Walterova, D. Antioxidant properties of silybin glycosides. Phytother. Res. 2002, 16 (Suppl. 1), S33–S39. [Google Scholar] [CrossRef]
- Mira, L.; Silva, M.; Manso, C. Scavenging of reactive oxygen species by silibinin dihemisuccinate. Biochem. Pharmacol. 1994, 48, 753–759. [Google Scholar] [CrossRef]
- Maheshwari, H.; Agarwal, R.; Patil, C.; Katare, O.P. Preparation and pharmacological evaluation of silibinin liposomes. Arzneimittelforschung 2003, 53, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Rai, A.; Reddy, N.D.; Raj, P.V.; Jain, P.; Deshpande, P.; Mathew, G.; Kutty, N.G.; Udupa, N.; Rao, C.M. Silymarin liposomes improves oral bioavailability of silybin besides targeting hepatocytes, and immune cells. Pharmacol. Rep. 2014, 66, 788–798. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-Q.; Hu, J.-H. Improvement of the Dissolution Rate of Silymarin by Means of Solid Dispersions. Chem. Pharm. Bull. (Tokyo) 2004, 52, 972–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, M.-F.; Jia, W.; Li, S.-S.; Xu, Z.-H.; Sun, X.; Wang, X.-R.; Zhang, Y.-Y.; Xie, G.-X. A new silymarin preparation based on solid dispersion technique. Adv. Ther. 2005, 22, 595–600. [Google Scholar] [CrossRef]
- Li, X.; Yuan, Q.; Huang, Y.; Zhou, Y.; Liu, Y. Development of Silymarin Self-Microemulsifying Drug Delivery System with Enhanced Oral Bioavailability. AAPS PharmSciTech 2010, 11, 672–678. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Wang, Y.; Que, L. Enhanced bioavailability of silymarin by self-microemulsifying drug delivery system. Eur. J. Pharm. Biopharm. 2006, 63, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Parveen, R.; Baboota, S.; Ali, J.; Ahuja, A.; Ahmad, S. Stability studies of silymarin nanoemulsion containing Tween 80 as a surfactant. J. Pharm. Bioallied Sci. 2015, 7, 321–324. [Google Scholar] [PubMed]
- Wang, Y.; Zhang, D.; Liu, Z.; Liu, G.; Duan, C.; Jia, L.; Feng, F.; Zhang, X.; Shi, Y.; Zhang, Q. In vitroandin vivoevaluation of silybin nanosuspensions for oral and intravenous delivery. Nanotechnology 2010, 21, 155104. [Google Scholar] [CrossRef]
- Hsu, W.-C.; Ng, L.-T.; Wu, T.-H.; Lin, L.-T.; Yen, F.-L.; Lin, C.-C.; Huang, L.-T. Characteristics and Antioxidant Activities of Silymarin Nanoparticles. J. Nanosci. Nanotechnol. 2012, 12, 2022–2027. [Google Scholar] [CrossRef] [PubMed]
- Polyak, S.J.; Ferenci, P.; Pawlotsky, J.M. Hepatoprotective and antiviral functions of silymarin components in hepatitis C virus infection. Hepatology 2013, 57, 1262–1271. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Virus | Substrate(s) | Method(s) | Suggested Mechanism | Reference |
|---|---|---|---|---|
| Hepatitis C virus (HCV) | Silymarin extract (MK-001) | Western blot and RT-PCR | Potentiation of the JAK-STAT antiviral signaling pathway | [8] |
| Silymarin and its-derived pure compounds | NS5B polymerase assay, luciferase reporter assay | Inhibition of HCV infection and the HCV-induced oxidative stress, as well as, the NS5B RdRp activity, NF-κB-dependent transcription, and T-cell receptor (TCR)-mediated proliferation | [9] | |
| Silymarin | NS5B polymerase assay, luciferase reporter assay, qPCR, and western blot | Inhibition of NS5B polymerase activity and blocking viral entry and transmission | [10] | |
| Silybin A, silybin B, and Legalon® SIL | RdRp Enzyme Assay, qPCR and luciferase reporter activity | Inhibition of the NS5B RNA-dependent RNA polymerase | [11] | |
| Silibinin and Legalon® SIL | HCV entry assay | Silibinin impeded HCV endosomal trafficking and blocked CME | [12] | |
| Silibinin | RT-PCR and luciferase reporter assay | Inhibition of HCV NS4B and hence the membranous web morphogenesis | [13] | |
| Silibinin nanoparticles | HCV entry assays and pharmacokinetic studies | Inhibition of HCV cell-to-cell spread and attenuation of HCV infection of PHHs | [15] | |
| Dengue virus (DENV) | Silymarin | Docking to NS4B | All three silymarin derivatives docked with high binding affinity (≥−8 kal/mol) to DENV NS4B | [18] |
| Influenza A virus (IAV) | Silymarin | CPE reduction method | Inhibition of late viral RNA synthesis | [22] |
| Silybin and amino acid derivatives (S0-S5) | CPE reduction method and plaque assay | S0 and S3 inhibited IAV replication and disrupted the formation of the Atg5-Atg12/Atg16L complex | [23] | |
| Human immunodeficiency virus (HIV) | Legalon® SIL | HIV replication in TZM-bl cells, peripheral blood mononuclear cells (PBMCs), and CEM | Attenuating cellular functions involved in T-cell activation, proliferation, and HIV infection | [26] |
| Silibinin and Legalon® SIL | HIV infection of PBMCs and CEM cells with respect to cell growth, ATP content, and metabolism | Perturbation of T-cell metabolism in vitro; Legalon® SIL additionally blocked HIV infection of T-cells | [27] | |
| Chikungunya virus (CHIKV) | Silymarin | CPE inhibition assay, RT-PCR and Western blot | Inhibition of CHIKV replication and proteinsynthesis | [28] |
| Mayaro virus (MAYV) | Silymarin | CPE inhibition, viral replication and plaque reduction assays in HepG2 cells | Inhibition of replication and ROS induction | [29] |
| Hepatitis B Virus (HBV) | Silibinin | HBV entry assay | Blockade of clathrin-mediated endocytosis | [31] |
| Virus | Substrate(s) | Analysis/Model | Route of Administration | Results | Reference |
|---|---|---|---|---|---|
| HCV | Legalon® SIL | HCV infection of uPA-SCID-chimeric mice with humanized livers | Intravenous | Legalon® SIL blocked HCV production and increased anti-inflammatory and anti-proliferative gene expressions without affecting serum albumin levels | [16] |
| IAV | Silybin derivatives (S0 and S3) | IAV infection of BALB/c mice | Oral | S0 and S3 increased the survival rate of mice (40% and 60% respectively), and S3 decreased virus titers in the lungs (100-fold) | [23] |
| HBV | Silymarin | HBV X protein (HBx) transgenic mice | Oral | Silymarin had no effect on HBx expression and late stage carcinogenesis, but recovered fatty acid change and liver pathology in the early stages of liver damage | [32] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-H.; Jassey, A.; Hsu, H.-Y.; Lin, L.-T. Antiviral Activities of Silymarin and Derivatives. Molecules 2019, 24, 1552. https://doi.org/10.3390/molecules24081552
Liu C-H, Jassey A, Hsu H-Y, Lin L-T. Antiviral Activities of Silymarin and Derivatives. Molecules. 2019; 24(8):1552. https://doi.org/10.3390/molecules24081552
Chicago/Turabian StyleLiu, Ching-Hsuan, Alagie Jassey, Hsin-Ya Hsu, and Liang-Tzung Lin. 2019. "Antiviral Activities of Silymarin and Derivatives" Molecules 24, no. 8: 1552. https://doi.org/10.3390/molecules24081552
APA StyleLiu, C. -H., Jassey, A., Hsu, H. -Y., & Lin, L. -T. (2019). Antiviral Activities of Silymarin and Derivatives. Molecules, 24(8), 1552. https://doi.org/10.3390/molecules24081552