2-Hydroxysorangiadenosine: Structure and Biosynthesis of a Myxobacterial Sesquiterpene–Nucleoside


 , and
, and
Abstract
:1. Introduction
2. Results
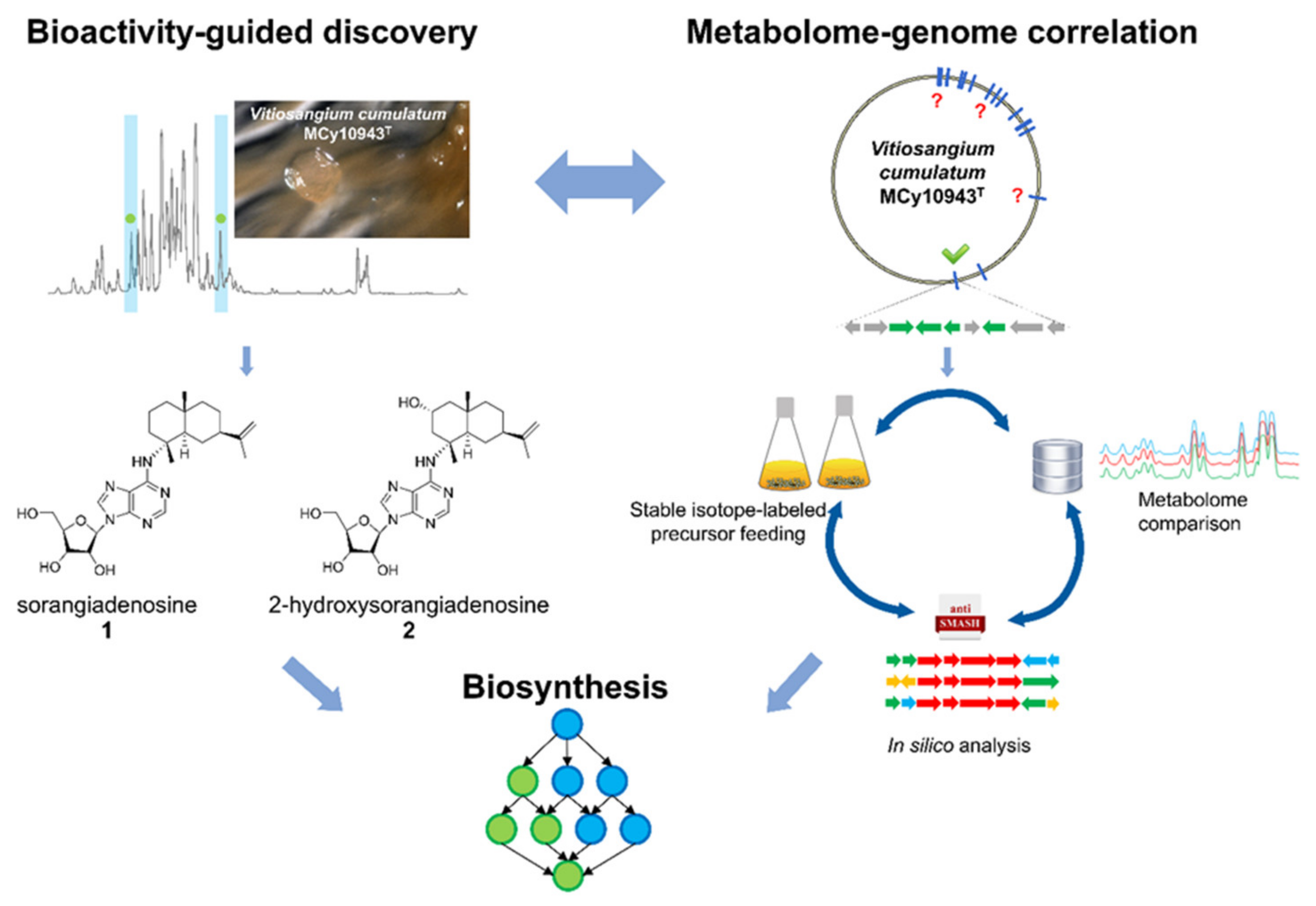
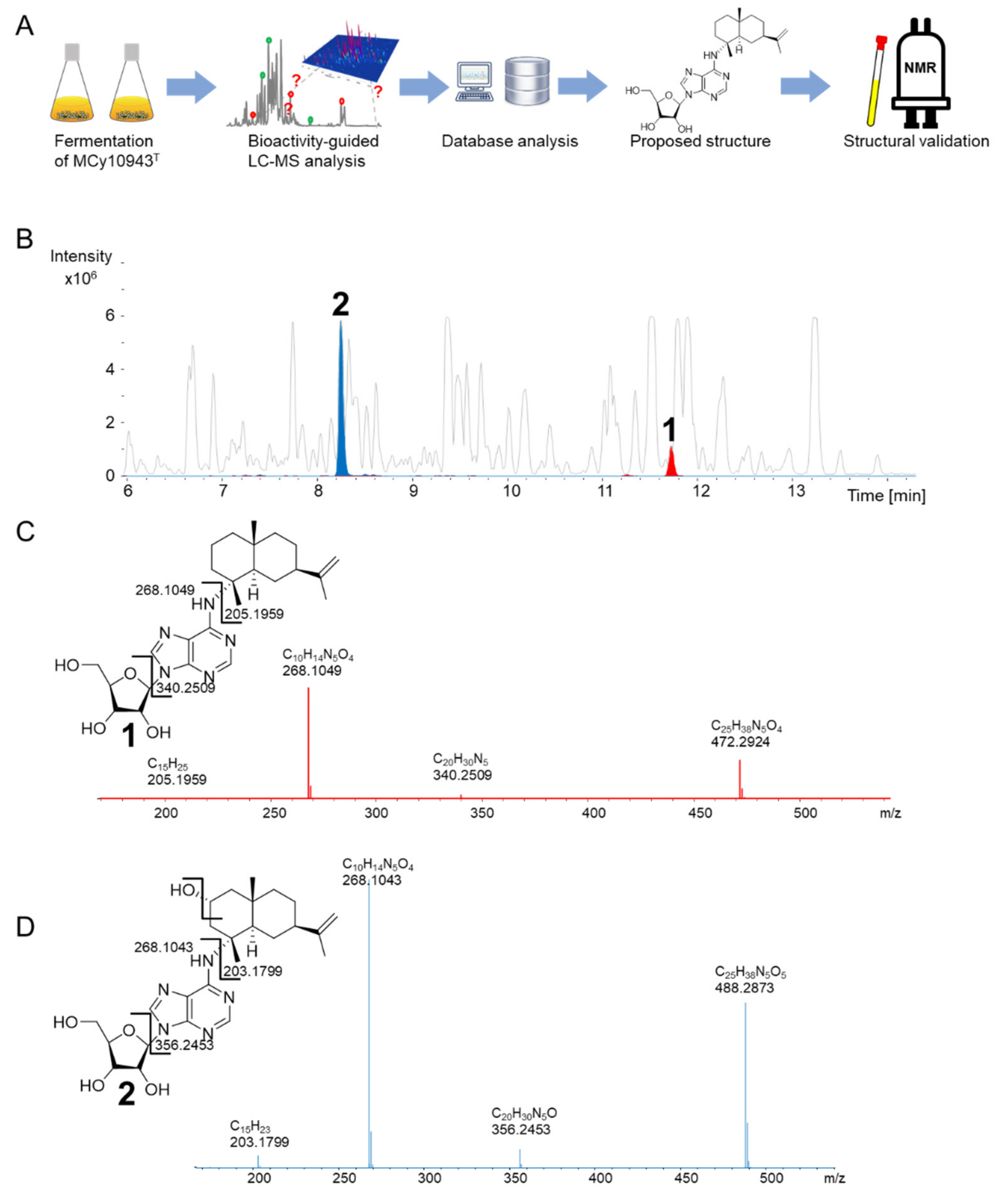
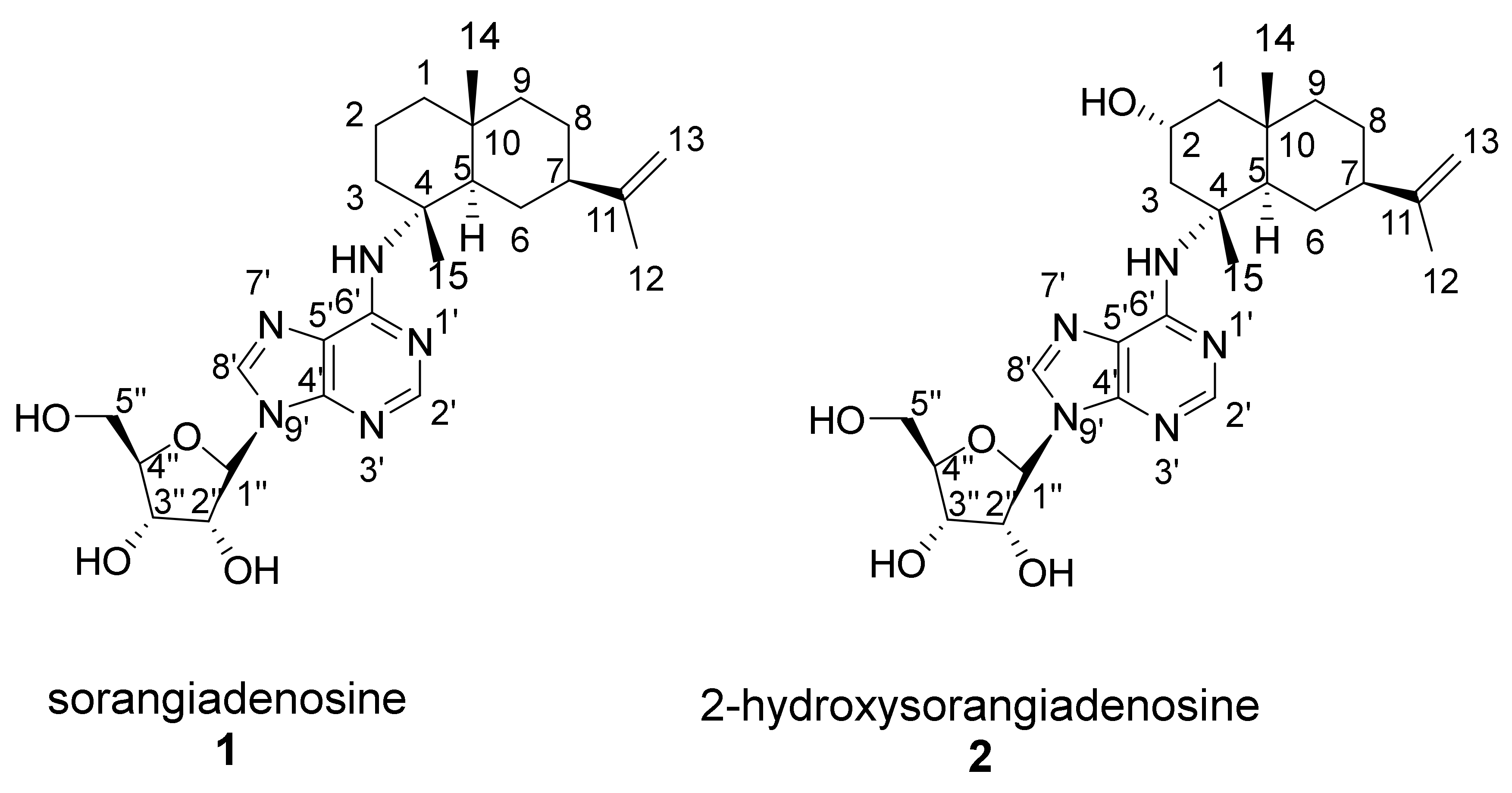
2.1. Discovery of 2-Hydroxysorangiadenosine and Sorangiadenosine
2.2. Biological Activity of 2-Hydroxysorangiadenosine and Sorangiadenosine
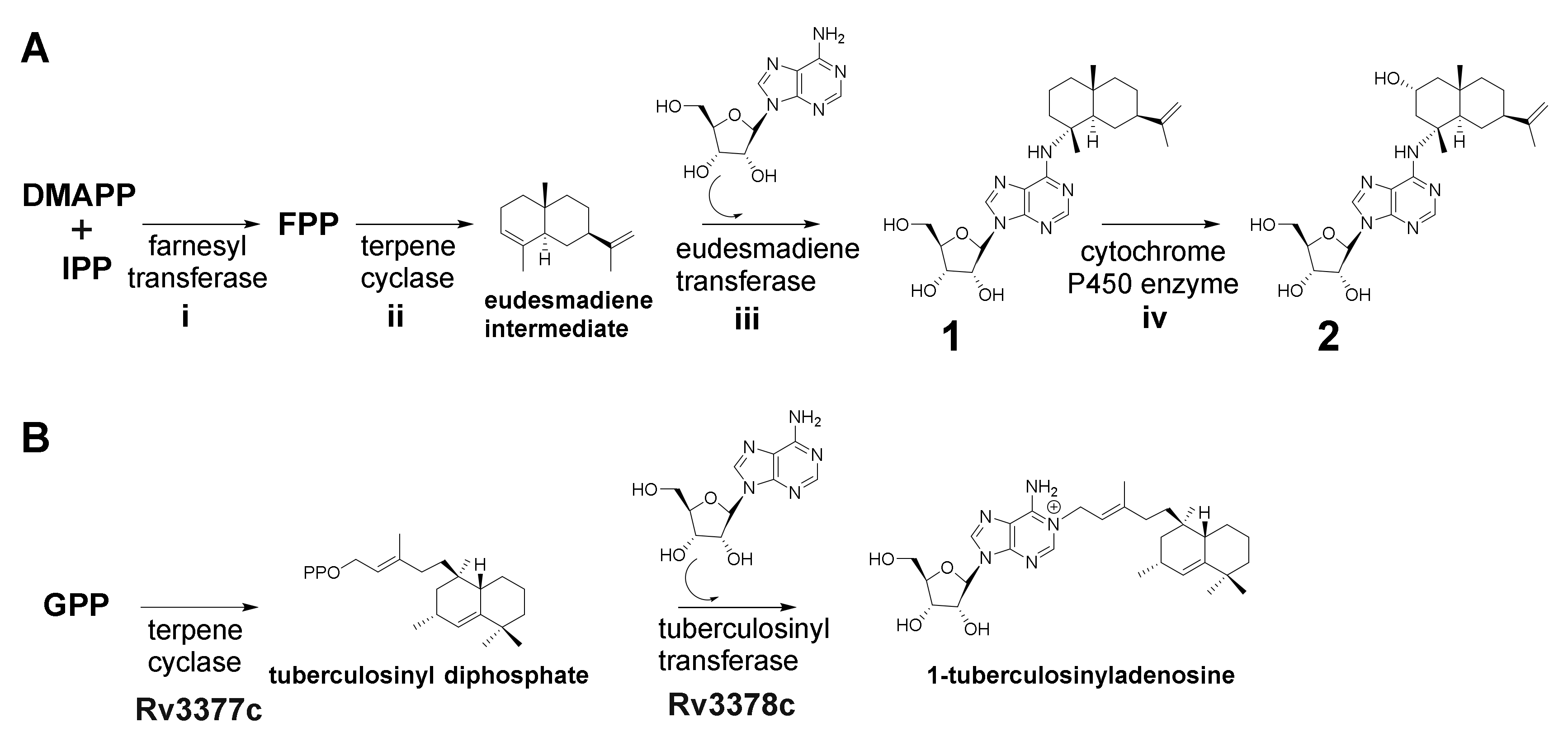
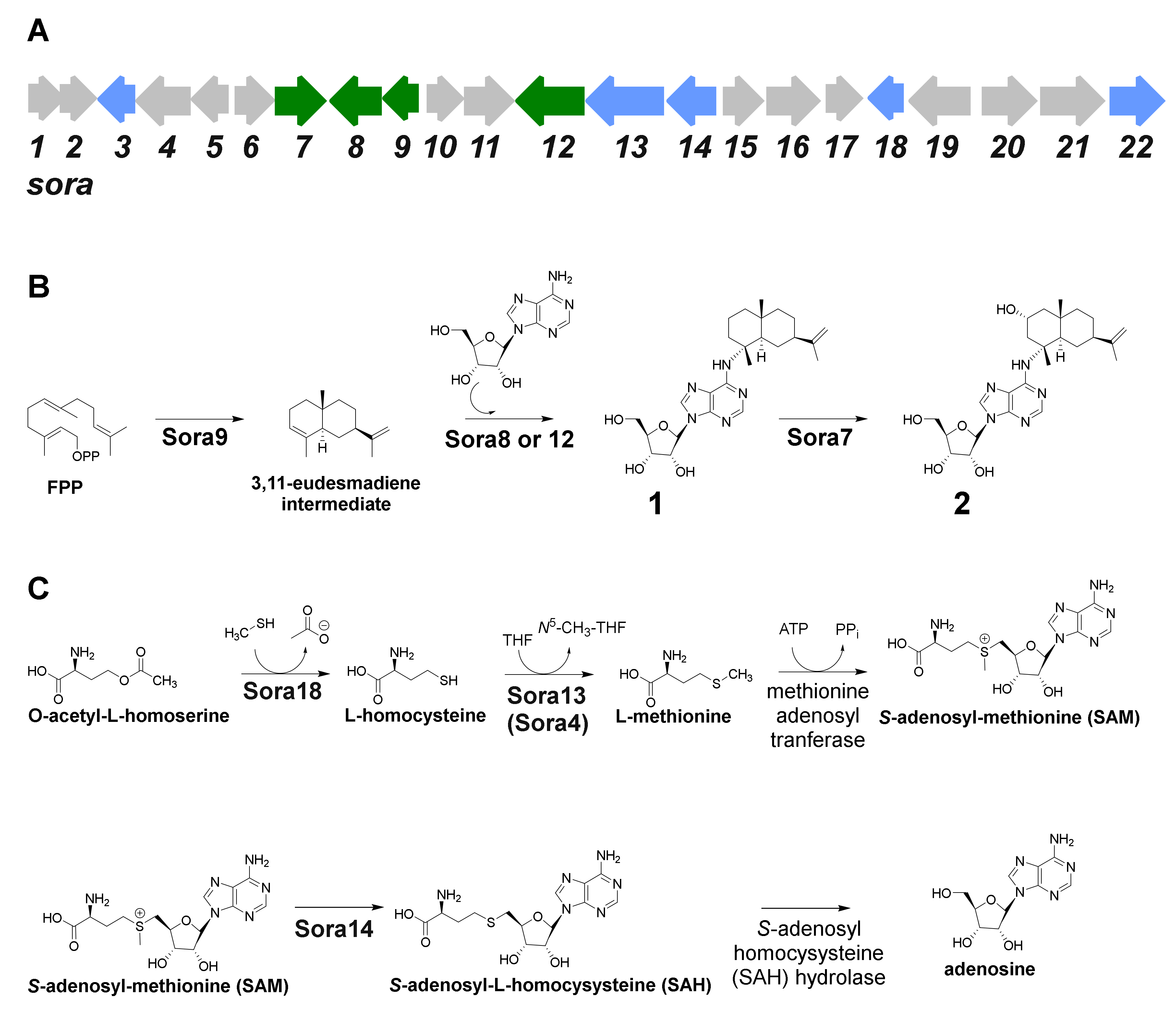
2.3. Identification of the 2-Hydroxysorangiadenosine Biosynthetic Gene Cluster
3. Discussion
4. Materials and Methods
4.1. Applied Software, Sequence Analysis and Bioinformatics Methods
4.2. Maintenance of Bacterial Cultures and Feeding Experiments with Stable Isotope-Labeled Building Blocks
4.3. Analysis of Secondary Metabolism of Broth Extracts
4.4. Compound Isolation
4.5. NMR-Based Structure Elucidation, Chiroptical and CD Measurement
4.6. Bioactivity Profiling
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Munoz-Dorado, J.; Marcos-Torres, F.J.; Garcia-Bravo, E.; Moraleda-Munoz, A.; Perez, J. Myxobacteria: Moving, Killing, Feeding, and Surviving Together. Front. Microbiol. 2016, 7, 2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, P.; Dey, A.; Vassallo, C.N.; Wall, D. How Myxobacteria Cooperate. J. Mol. Biol. 2015, 427, 3709–3721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, J.; Fayad, A.A.; Müller, R. Natural products from myxobacteria: Novel metabolites and bioactivities. Nat. Prod. Rep. 2017, 34, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Gerth, K.; Bedorf, N.; Irschik, H.; Höfle, G.; Reichenbach, H. The soraphens: A family of novel antifungal compounds from Sorangium cellulosum (Myxobacteria). I. Soraphen A1α: Fermentation, isolation, biological properties. J. Antibiot. 1994, 47, 23–31. [Google Scholar] [CrossRef]
- Baumann, S.; Herrmann, J.; Raju, R.; Steinmetz, H.; Mohr, K.I.; Hüttel, S.; Harmrolfs, K.; Stadler, M.; Müller, R. Cystobactamids: Myxobacterial topoisomerase inhibitors exhibiting potent antibacterial activity. Angew. Chem. Int. Ed. 2014, 53, 14605–14609. [Google Scholar] [CrossRef]
- Gerth, K.; Steinmetz, H.; Höfle, G.; Jansen, R. Chlorotonil A, a Macrolide with a Unique gem-Dichloro-1,3-dione Functionality from Sorangium cellulosum, So ce1525. Angew. Chem. Int. Ed. Engl. 2008, 47, 600–602. [Google Scholar] [CrossRef]
- Plaza, A.; Garcia, R.; Bifulco, G.; Martinez, J.P.; Hüttel, S.; Sasse, F.; Meyerhans, A.; Stadler, M.; Müller, R. Aetheramides A and B, potent HIV-inhibitory depsipeptides from a myxobacterium of the new genus “Aetherobacter”. Org. Lett. 2012, 14, 2854–2857. [Google Scholar] [CrossRef]
- Sasse, F.; Steinmetz, H.; Heil, J.; Höfle, G.; Reichenbach, H. Tubulysins, new cytostatic peptides from myxobacteria acting on microtubuli. Production, isolation, physico-chemical and biological properties. J. Antibiot. 2000, 53, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, A.; Chai, Y.; Pistorius, D.; Elnakady, Y.A.; Herrmann, J.E.; Weissman, K.J.; Kazmaier, U.; Müller, R. Pretubulysin, a potent and chemically accessible tubulysin precursor from Angiococcus disciformis. Angew. Chem. Int. Ed. Engl. 2009, 48, 4422–4425. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Wenzel, S.C.; Bode, H.B.; Müller, R.; Schulz, S. Biosynthesis of volatiles by the myxobacterium Myxococcus xanthus. ChemBioChem 2004, 5, 778–787. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Bode, H.B.; Wenzel, S.C.; Müller, R.; Schulz, S. Biosynthesis and identification of volatiles released by the myxobacterium Stigmatella aurantiaca. ChemBioChem 2005, 6, 2023–2033. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S.; Bode, H.B.; Mahmud, T.; Müller, R.; Schulz, S. A novel type of geosmin biosynthesis in myxobacteria. J. Org. Chem. 2005, 70, 5174–5182. [Google Scholar] [CrossRef] [PubMed]
- Brock, N.L.; Ravella, S.R.; Schulz, S.; Dickschat, J.S. A Detailed View of 2-Methylisoborneol Biosynthesis. Angew. Chem. Int. Ed. Engl. 2013, 52, 2100–2104. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S.; Nawrath, T.; Thiel, V.; Kunze, B.; Müller, R.; Schulz, S. Biosynthesis of the off-flavor 2-methylisoborneol by the myxobacterium Nannocystis exedens. Angew. Chem. Int. Ed. Engl. 2007, 46, 8287–8290. [Google Scholar] [CrossRef]
- Schifrin, A.; Ly, T.T.; Günnewich, N.; Zapp, J.; Thiel, V.; Schulz, S.; Hannemann, F.; Khatri, Y.; Bernhardt, R. Characterization of the Gene Cluster CYP264B1-geoA from Sorangium cellulosum So ce56: Biosynthesis of (+)-Eremophilene and Its Hydroxylation. ChemBioChem 2015, 16, 337–344. [Google Scholar] [CrossRef]
- Schifrin, A.; Khatri, Y.; Kirsch, P.; Thiel, V.; Schulz, S.; Bernhardt, R. A single terpene synthase is responsible for a wide variety of sesquiterpenes in Sorangium cellulosum So ce56. Org. Biomol. Chem. 2016, 14, 3385–3393. [Google Scholar] [CrossRef]
- Ahn, J.W.; Jang, K.H.; Chung, S.C.; Oh, K.B.; Shin, J. Sorangiadenosine, a new sesquiterpene adenoside from the myxobacterium Sorangium cellulosum. Org. Lett. 2008, 10, 1167–1169. [Google Scholar] [CrossRef]
- Awal, R.P.; Garcia, R.; Gemperlein, K.; Wink, J.; Kunwar, B.; Parajuli, N.; Müller, R. Vitiosangium cumulatum gen. nov., sp. nov. and Vitiosangium subalbum sp. nov., soil myxobacteria, and emended descriptions of the genera Archangium and Angiococcus, and of the family Cystobacteraceae. Int. J. Syst. Evol. Microbiol. 2017, 67, 1422–1430. [Google Scholar] [CrossRef]
- Mai, N.T.; Cuc, N.T.; Quang, T.H. Chemical constituents of Datura metel L. Viet. J. Chem. 2017, 55, 188–195. [Google Scholar]
- Ciuffreda, P.; Casati, S.; Manzocchi, A. Complete 1H and 13C NMR spectral assignment of α- and β-adenosine, 2′-deoxyadenosine and their acetate derivatives. Magn. Reson. Chem. 2007, 45, 781–784. [Google Scholar] [CrossRef]
- Nakamukai, S.; Ise, Y.; Ohtsuka, S.; Okada, S.; Matsunaga, S. Isolation and identification of N6-isopentenyladenosine as the cytotoxic constituent of a marine sponge Oceanapia sp. Biosci. Biotechnol. Biochem. 2019, 83, 1985–1988. [Google Scholar] [CrossRef] [PubMed]
- Brunner, W.C.; Maestre, M.F. Circular dichroism of some mononucleosides. Biopolymers 1975, 14, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.S. Circular dichroism of nucleic acid monomers. I. Calculated adenosine and 2′-deoxyadenosine CD spectra. Biopolymers 1980, 19, 1017–1038. [Google Scholar] [CrossRef]
- Miles, D.W.; Farmer, M.; Eyring, H. Calculations of the circular dichroism of adenosine derivatives constrained in the syn form. Proc. Natl. Acad. Sci. USA 1980, 77, 3398–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, K. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 2013, 12, 371–387. [Google Scholar] [CrossRef]
- Rojas, E.R.; Billings, G.; Odermatt, P.D.; Auer, G.K.; Zhu, L.; Miguel, A.; Chang, F.; Weibel, D.B.; Theriot, J.A.; Huang, K.C. The outer membrane is an essential load-bearing element in Gram-negative bacteria. Nature 2018, 559, 617–621. [Google Scholar] [CrossRef]
- Bode, H.B.; Ring, M.W.; Schwär, G.; Altmeyer, M.O.; Kegler, C.; Jose, I.R.; Singer, M.; Müller, R. Identification of additional players in the alternative biosynthesis pathway to isovaleryl-CoA in the myxobacterium Myxococcus xanthus. ChemBioChem 2009, 10, 128–140. [Google Scholar] [CrossRef]
- Li, Y.; Luxenburger, E.; Müller, R. An alternative isovaleryl CoA biosynthetic pathway involving a previously unknown 3-methylglutaconyl CoA decarboxylase. Angew. Chem. Int. Ed. Engl. 2012, 52, 1304–1308. [Google Scholar] [CrossRef]
- Li, R.; Wijma, H.J.; Song, L.; Cui, Y.; Otzen, M.; Tian, Y.; Du, J.; Li, T.; Niu, D.; Chen, Y.; et al. Computational redesign of enzymes for regio- and enantioselective hydroamination. Nat. Chem. Biol. 2018, 14, 664–670. [Google Scholar] [CrossRef]
- Severin, R.; Doye, S. The catalytic hydroamination of alkynes. Chem. Soc. Rev. 2007, 36, 1407–1420. [Google Scholar] [CrossRef]
- Helfrich, E.J.N.; Lin, G.-M.; Voigt, C.A.; Clardy, J. Bacterial terpene biosynthesis: Challenges and opportunities for pathway engineering. Beilstein J. Org. Chem. 2019, 15, 2889–2906. [Google Scholar] [CrossRef] [PubMed]
- Layre, E.; Lee, H.J.; Young, D.C.; Martinot, A.J.; Buter, J.; Minnaard, A.J.; Annand, J.W.; Fortune, S.M.; Snider, B.B.; Matsunaga, I.; et al. Molecular profiling of Mycobacterium tuberculosis identifies tuberculosinyl nucleoside products of the virulence-associated enzyme Rv3378c. Proc. Natl. Acad. Sci. USA 2014, 111, 2978–2983. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, D.C.; Young, J.; Carmichael, R.; Tan, J.; Taylor, R.E. Draft Genome Sequence of Gephyronic Acid Producer Cystobacter violaceus Strain Cb vi76. Genome Announc. 2014, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adaikpoh, B.I.; Dowd, S.E.; Stevens, D.C. Draft Genome Sequence of Archangium sp. Strain Cb G35. Genome Announc. 2017, 5, e01678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giglio, S.; Jiang, J.; Saint, C.P.; Cane, D.E.; Monis, P.T. Isolation and characterization of the gene associated with geosmin production in cyanobacteria. Environ. Sci. Technol. 2008, 42, 8027–8032. [Google Scholar]
- Jiang, J.Y.; He, X.F.; Cane, D.E. Biosynthesis of the earthy odorant geosmin by a bifunctional Streptomyces coelicolor enzyme. Nat. Chem. Biol. 2007, 3, 711–715. [Google Scholar] [CrossRef] [Green Version]
- Botella, J.A.; Murillo, F.J.; Ruiz-Vazquez, R. A cluster of structural and regulatory genes for light-induced carotenogenesis in Myxococcus xanthus. Eur. J. Biochem. 1995, 233, 238–248. [Google Scholar] [CrossRef]
- Lopez-Rubio, J.J.; Elias-Arnanz, M.; Padmanabhan, S.; Murillo, F.J. A repressor-antirepressor pair links two loci controlling light-induced carotenogenesis in Myxococcus xanthus. J. Biol. Chem. 2002, 277, 7262–7270. [Google Scholar] [CrossRef] [Green Version]
- Perez-Marin, M.C.; Padmanabhan, S.; Polanco, M.C.; Murillo, F.J.; Elias-Arnanz, M. Vitamin B12 partners the CarH repressor to downregulate a photoinducible promoter in Myxococcus xanthus. Mol. Microbiol. 2008, 67, 804–819. [Google Scholar] [CrossRef]
- Cervantes, M.; Murillo, F.J. Role for Vitamin B12 in Light Induction of Gene Expression in the Bacterium Myxococcus xanthus. J. Bacteriol. 2002, 184, 2215–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickschat, J.S. Bacterial terpene cyclases. Nat. Prod. Rep. 2015, 33, 87–110. [Google Scholar] [CrossRef] [PubMed]
- Harada, F.; Gross, H.J.; Kimura, F.; Chang, S.H.; Nishimura, S.; RajBhandary, U.L. 2-Methylthio N6-(Δ2-isopentenyl) adenosine: A component of E. coli tyrosine transfer RNA. Biochem. Biophys. Res. Commun. 1968, 33, 299–306. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; Petrášek, J. Auxins and cytokinins—The dynamic duo of growth-regulating phytohormones heading for new shores. New Phytol. 2019, 221, 1187–1190. [Google Scholar] [CrossRef] [Green Version]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The yin-yang of hormones: Cytokinin and auxin interactions in plant development. Plant. Cell 2015, 27, 44–63. [Google Scholar] [CrossRef] [Green Version]
- MacLeod, J.K.; Summons, R.E.; Parker, C.W.; Letham, D.S. Lupinic acid, a purinyl amino acid and a novel metabolite of zeatin. J. Chem. Soc. Chem. Commun. 1975, 19, 809–810. [Google Scholar] [CrossRef]
- Abdjul, D.B.; Yamazaki, H.; Kanno, S.-I.; Takahashi, O.; Kirikoshi, R.; Ukai, K.; Namikoshi, M. Structures and Biological Evaluations of Agelasines Isolated from the Okinawan Marine Sponge Agelas nakamurai. J. Nat. Prod. 2015, 78, 1428–1433. [Google Scholar] [CrossRef]
- Cullen, E.; Devlin, J.P. Agelasine: A Novel Quaternary 9-Methyladenine from the Sponge Agelas dispar. Can. J. Chem. 1975, 53, 1690–1691. [Google Scholar] [CrossRef]
- Fathi-Afshar, R.; Allen, T.M. Biologically active metabolites from Agelas mauritiana. Can. J. Chem. 1988, 66, 45–50. [Google Scholar] [CrossRef]
- Yosief, T.; Rudi, A.; Kashman, Y. Asmarines A-F, novel cytotoxic compounds from the marine sponge Raspailia species. J. Nat. Prod. 2000, 63, 299–304. [Google Scholar] [CrossRef]
- Gordaliza, M. Terpenyl-purines from the sea. Mar. Drugs 2009, 7, 833–849. [Google Scholar] [CrossRef] [Green Version]
- Hug, J.J.; Müller, R. Host Development for Heterologous Expression and Biosynthetic Studies of Myxobacterial Natural Products: Comprehensive Natural Products III: Chemistry and Biology, Chapter 14818. Ref. Modul. Chem. Mol. Sci. Chem. Eng. 2020, in press. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds sorangiadenosine and 2-hydroxysorangiadenosine are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | MIC (µg/mL) of Sorangiadenosine (1) | MIC (µg/mL) of 2-hydroxysorangiadenosine (2) |
|---|---|---|
| B. subtilis DSM 10 | 16 | 64 |
| E. coli WT (DSM 1116) | >128 | >128 |
| E. coli acrB JW0451-2 | >128 | >128 |
| P. aeruginosa PA14 | >128 | >128 |
| S. aureus Newman | 32 | 128 |
| C. freundii DSM 30039 | >128 | >128 |
| A. baumanii DSM 30007 | >128 | >128 |
| M. hiemalis DSM 2656 | 128 | >128 |
| P. anomala DSM 6766 | >128 | >128 |
| M. smegmatis MC2 155 | >128 | >128 |
| C. albicans DSM 1665 | 128 | >128 |
| Cancer Cell Line | IC50 (µg/mL) of Sorangiadenosine 1 | IC50 (µg/mL) of 2-hydroxysorangiadenosine 2 | IC50 (µg/mL) of Doxirubicin |
|---|---|---|---|
| HCT-116 | 30.00 | 52.00 | 0.6 |
| KB-3-1 | 39.46 | >111.1 | 0.09 |
| No. | Gene Cluster | Size (bp) | Location | Terpene Cyclase | Associated Biosynthesis 1 |
|---|---|---|---|---|---|
| 1 | Terpene | 27920 | 653776–674051 | 1 × Type I, 2 × Type II | This study |
| 2 | Terpene | 42274 | 1269631–1311904 | 1 × Type I | Geosmin [36,37] |
| 3 | Terpene | 41089 | 3415277–3456365 | 1 × Type I | Genome-Metab |
| 4 | Terpene | 41071 | 3447909–3488979 | 1 × Type I | |
| 5 | Terpene | 40978 | 3603511–3644488 | 1 × Type I | Genome-Metab |
| 6 | Terpene | 41041 | 4852259–4893299 | 1 × Type I | Genome-Metab |
| 7 | Terpene/Type_III_PKS | 69038 | 7156603–7225640 | 1 × Type II | Carotenoid [38,39,40,41] |
| 8 | Terpene/TfuA-rel. | 50108 | 8404785–8454892 | 1 × Type I | |
| 9 | Terpene/RiPP | 78053 | 12603875–12681927 | 1 × Type I | Geosmin [36,37] |
| Gene | Size (aa) | Proposed Function | Closest Homologue | Coverage/Similarity (%) |
|---|---|---|---|---|
| sora1 | 378 | Oxidoreductase | WP_108075222 | 100/95.99 |
| sora2 | 72 | Hypothetical protein | WP_108075223 | 100/93.90 |
| sora3 | 173 | Hypothetical protein | WP_108075224 | 100/82.92 |
| sora4 | 364 | Methionine synthase (MetE) | WP_108075225 | 100/95.87 |
| sora5 | 304 | Hydrolase | WP_108075226 | 100/90.43 |
| sora6 | 284 | Hypothetical protein | WP_108075227 | 95/92.25 |
| sora7 | 457 | Cytochrome P450 enzyme | WP_108069092 | 98/95.67 |
| sora8 | 518 | Eudesmadiene transferase | WP_047856205 | 99/80.31 |
| sora9 | 333 | Terpene cyclase | WP_073564250 | 99.78/79.09 |
| sora10 | 387 | Hypothetical protein | WP_095976240 | 91/56.91 |
| sora11 | 609 | Sensory transducer | WP_146210122 | 92/74.69 |
| sora12 | 509 | Eudesmadiene transferase | WP_108075230 | 100/73.80 |
| sora13 | 1171 | Methionine synthase (MetH) | WP_108075232 | 100/97.61 |
| sora14 | 299 | SAM-dependent methyltransferase | WP_108075233 | 100/97.32 |
| sora15 | 292 | Patatin lipid acyl hydrolases | WP_108075234 | 100/98.97 |
| sora16 | 375 | Dehydrogenase | WP_052519033 | 91/82.11 |
| sora17 | 205 | TetR transcriptional regulator | WP_073564266 | 100/79.62 |
| sora18 | 147 | O-acetylhomoserine sulfhydrylase | WP_108075236 | 100/97.26 |
| sora19 | 302 | Hypothetical protein | WP_108075237 | 100/88.70 |
| sora20 | 449 | Thioredoxin | WP_140874099 | 99/54.81 |
| sora21 | 279 | Hypothetical protein | WP_158079939 | 100/77.34 |
| sora22 | 630 | Phosphotransferase | WP_073564278 | 100/79.83 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okoth, D.A.; Hug, J.J.; Garcia, R.; Spröer, C.; Overmann, J.; Müller, R. 2-Hydroxysorangiadenosine: Structure and Biosynthesis of a Myxobacterial Sesquiterpene–Nucleoside. Molecules 2020, 25, 2676. https://doi.org/10.3390/molecules25112676
Okoth DA, Hug JJ, Garcia R, Spröer C, Overmann J, Müller R. 2-Hydroxysorangiadenosine: Structure and Biosynthesis of a Myxobacterial Sesquiterpene–Nucleoside. Molecules. 2020; 25(11):2676. https://doi.org/10.3390/molecules25112676
Chicago/Turabian StyleOkoth, Dorothy A., Joachim J. Hug, Ronald Garcia, Cathrin Spröer, Jörg Overmann, and Rolf Müller. 2020. "2-Hydroxysorangiadenosine: Structure and Biosynthesis of a Myxobacterial Sesquiterpene–Nucleoside" Molecules 25, no. 11: 2676. https://doi.org/10.3390/molecules25112676
APA StyleOkoth, D. A., Hug, J. J., Garcia, R., Spröer, C., Overmann, J., & Müller, R. (2020). 2-Hydroxysorangiadenosine: Structure and Biosynthesis of a Myxobacterial Sesquiterpene–Nucleoside. Molecules, 25(11), 2676. https://doi.org/10.3390/molecules25112676