
Rutamarin: Efficient Liquid–Liquid Chromatographic Isolation from Ruta graveolens L. and Evaluation of Its In Vitro and In Silico MAO-B Inhibitory Activity

 , , , , ,
, , , , , 
Abstract
:
1. Introduction
2. Results and Discussion
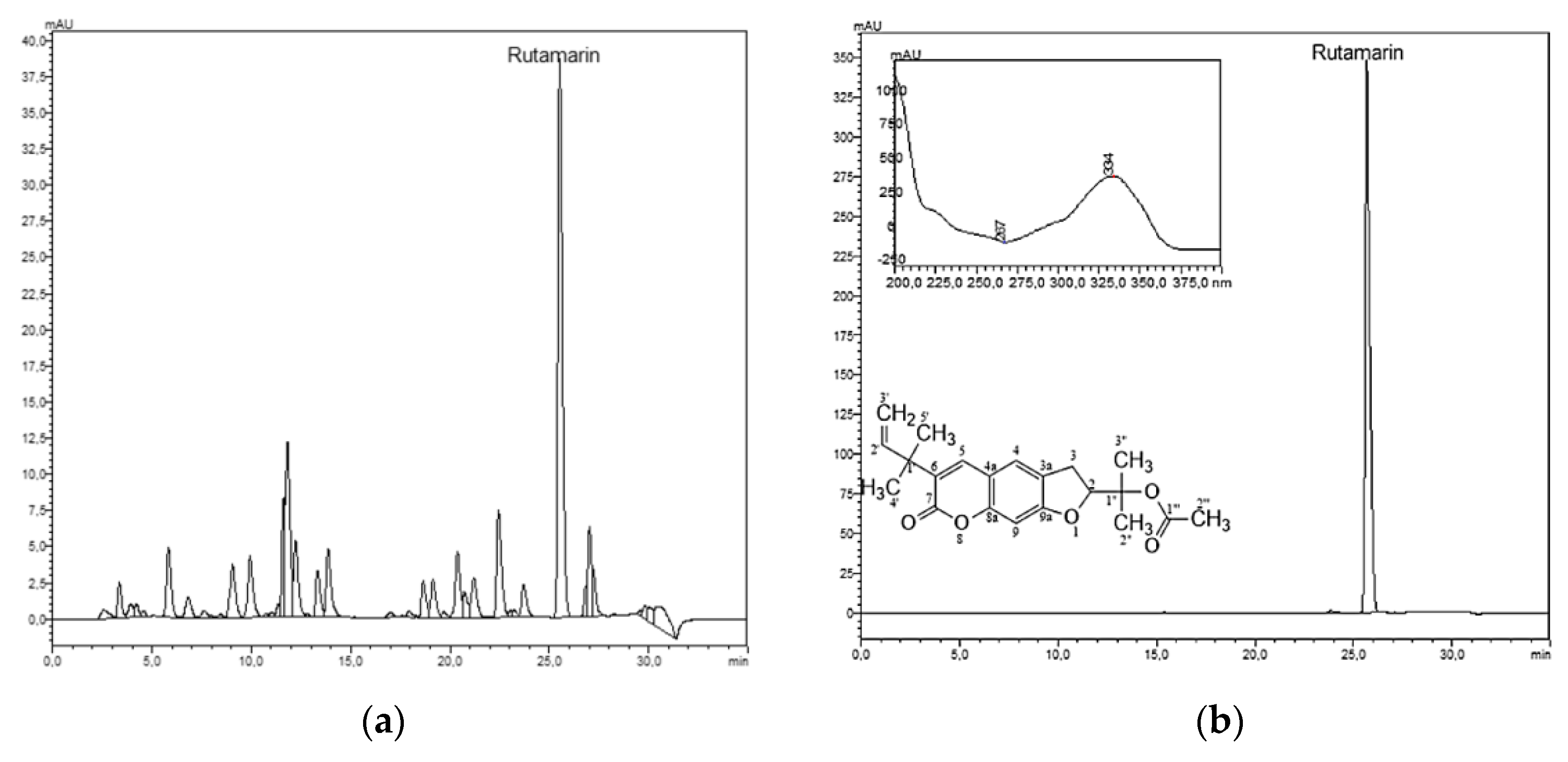
2.1. Phytochemical Study: Liquid–Liquid Chromatography Isolation of Rutamarin
2.2. In Vitro Study: hMAO-A and hMAO-B Inhibitory Activity
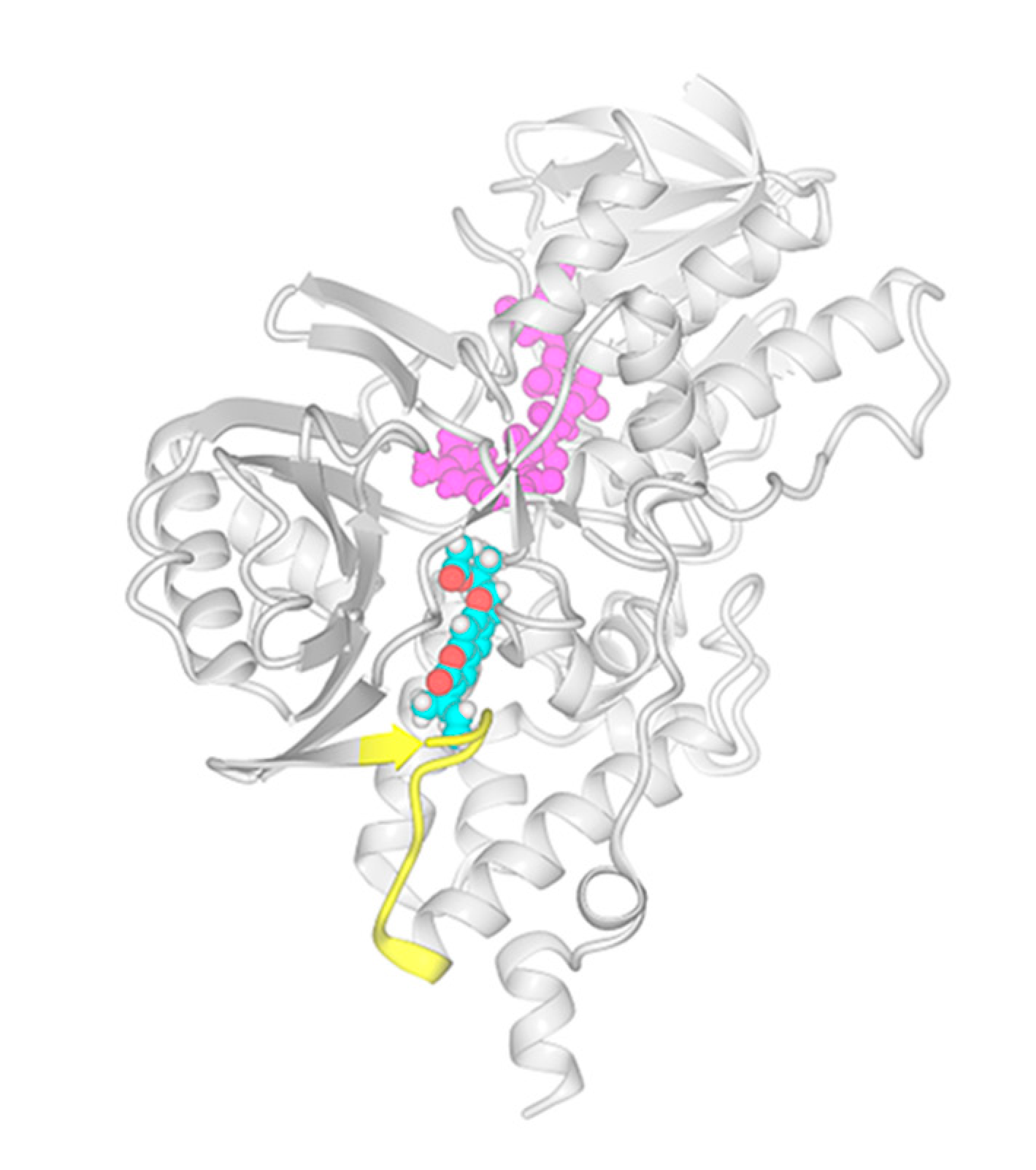
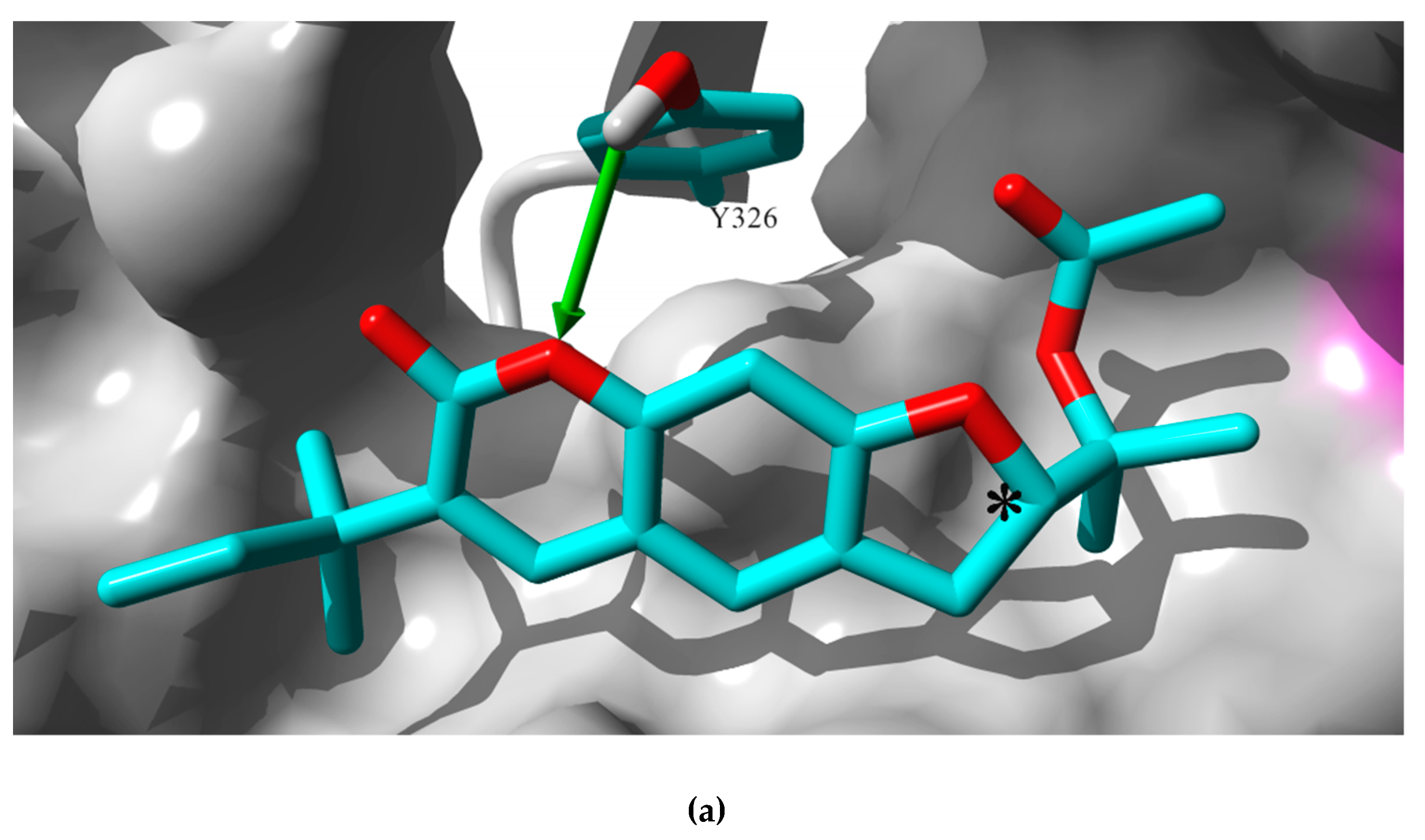
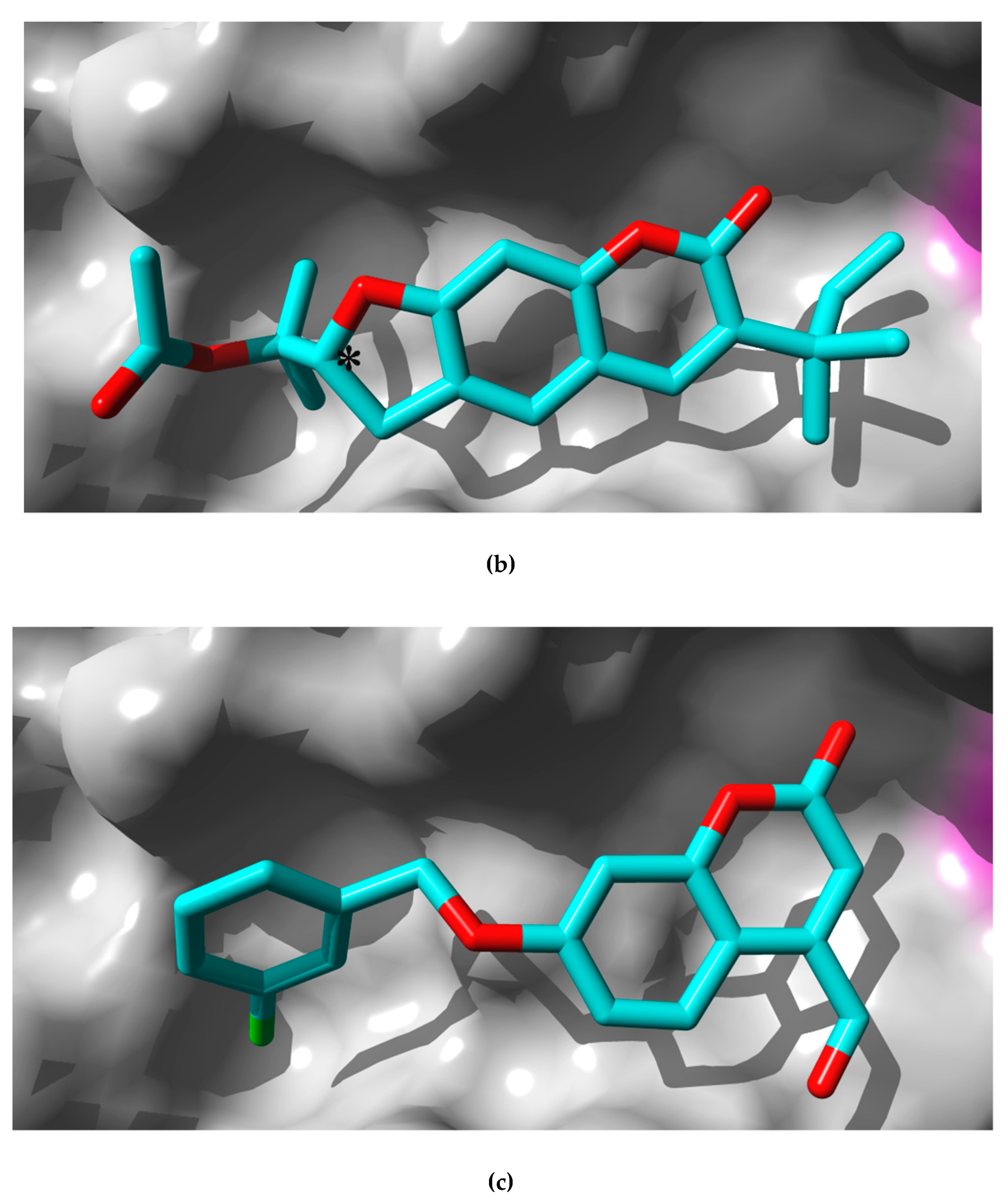
2.3. In Silico Study: Molecular Mechanisms of Rutamarin Interactions with hMAO-B
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material and Extraction
3.3. Liquid–Liquid Chromatographic Separation
3.4. HPLC–DAD Analysis
3.5. Structure Elucidation
3.6. In Vitro Enzymatic Activity Assay
3.7. Docking of Rutamarin into hMAO-B Binding Site
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| CNS | Central nervous system |
| H2O2 | Hydrogen peroxide |
| HEMWat | n-hexane/ethyl acetate/methanol/water |
| LLC | liquid–liquid chromatography |
| MAOs | Monoamine oxidases |
| PD | Parkinson’s disease |
| WHO | World Health Organization |
References
- Association, A. Alzheimer’s disease facts and figures. Alzheimers Dement. 2019, 15, 321–387. [Google Scholar]
- Edmondson, D.E.; Binda, C. Monoamine oxidases. In Membrane Protein Complexes: Structure and Function; Springer: Berlin, Germany, 2018; pp. 117–139. [Google Scholar]
- Kennedy, B.P.; Ziegler, M.G.; Alford, M.; Hansen, L.A.; Thal, L.J.; Masliah, E. Early and persistent alterations in prefrontal cortex MAO A and B in Alzheimer’s disease. J. Neural Transm. 2003, 110, 789–801. [Google Scholar] [CrossRef]
- Burke, W.J.; Li, S.W.; Chung, H.D.; Ruggiero, D.A.; Kristal, B.S.; Johnson, E.M.; Lampe, P.; Kumar, V.B.; Franko, M.; Williams, E.A. Neurotoxicity of MAO metabolites of catecholamine neurotransmitters: Role in neurodegenerative diseases. Neurotoxicology 2004, 25, 101–115. [Google Scholar] [CrossRef]
- Liu, Z.; Cai, W.; Lang, M.; Yan, R.; Li, Z.; Zhang, G.; Yu, P.; Wang, Y.; Sun, Y.; Zhang, Z. Neuroprotective effects and mechanisms of action of multifunctional agents targeting free radicals, monoamine oxidase B and cholinesterase in Parkinson’s disease model. J. Mol. Neurosci. 2017, 61, 498–510. [Google Scholar] [CrossRef]
- He, Q.; Liu, J.; Lan, J.S.; Ding, J.; Sun, Y.; Fang, Y.; Jiang, N.; Yang, Z.; Sun, Y.; Jin, Y. Coumarin-dithiocarbamate hybrids as novel multitarget AChE and MAO-B inhibitors against Alzheimer’s disease: Design, synthesis and biological evaluation. Bioorg. Chem. 2018, 81, 512–528. [Google Scholar] [CrossRef]
- Tao, D.; Wang, Y.; Bao, X.O.; Yang, B.B.; Gao, F.; Wang, L.; Zhang, L.; Li, L. Discovery of coumarin Mannich base derivatives as multifunctional agents against monoamine oxidase B and neuroinflammation for the treatment of Parkinson’s disease. Eur. J. Med. Chem. 2019, 173, 203–212. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, P.; Hu, Y.; Liu, T.; Sun, J.; Wang, X. Synthesis and biological evaluation of 3-arylcoumarins as potential anti-Alzheimer’s disease agents. J. Enzym. Inhib. Med. Chem. 2019, 34, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Huong, D.T.L.; Choi, H.C.; Rho, T.C.; Lee, H.S.; Lee, M.K.; Kim, Y.H. Inhibitory activity of monoamine oxidase by coumarins from Peucedanum japonicum. Arch. Pharm. Res. 1999, 22, 324–326. [Google Scholar] [CrossRef]
- Jeong, S.H.; Han, X.H.; Hong, S.S.; Hwang, J.S.; Hwang, J.H.; Lee, D.; Lee, M.K.; Ro, J.S.; Hwang, B.Y. Monoamine oxidase inhibitory coumarins from the aerial parts of Dictamnus albus. Arch. Pharm. Res. 2006, 29, 1119. [Google Scholar] [CrossRef]
- Jo, Y.S.; Huong, D.T.L.; Bae, K.H.; Lee, M.K.; Kim, Y.H. Monoamine oxidase inhibitory coumarin from Zanthoxylum schinifolium. Planta Med. 2002, 68, 84–85. [Google Scholar] [CrossRef]
- Hossain, C.F.; Okuyama, E.; Yamazaki, M. A new series of coumarin derivatives having monoamine oxidase inhibitory activity from Monascus anka. Chem. Pharm. Bull. 1996, 44, 1535–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande Tovar, C.D.; Delgado-Ospina, J.; Navia Porras, D.P.; Peralta-Ruiz, Y.; Cordero, A.P.; Castro, J.I.; Valencia, C.; Noé, M.; Mina, J.H.; Chaves López, C. Colletotrichumgloesporioides inhibition in situ by chitosan-Ruta graveolens essential oil coatings: Effect on microbiological, physicochemical, and organoleptic properties of Guava (Psidium guajava L.) during room temperature storage. Biomolecules 2019, 9, 399. [Google Scholar] [CrossRef] [Green Version]
- Semerdjieva, I.B.; Burducea, M.; Astatkie, T.; Zheljazkov, V.D.; Dincheva, I. Essential oil composition of Ruta graveolens L. Fruits and Hyssopus officinalis subsp. aristatus (Godr.) Nyman biomass as a function of hydrodistillation time. Molecules 2019, 24, 4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande Tovar, C.D.; Castro, J.I.; Valencia Llano, C.H.; Navia Porras, D.P.; Delgado Ospina, J.; Valencia Zapata, M.E.; Herminsul Mina Hernandez, J.; Chaur, M.N. Synthesis, characterization, and histological evaluation of chitosan-Ruta graveolens essential oil films. Molecules 2020, 25, 1688. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, O.M.; Vieira, L.C.C.; Bellete, B.S.; King-Diaz, B.; Lotina-Hennsen, B.; Da Silva, M.F.; Veiga, T.A.M. Evaluation of alkaloids isolated from Ruta graveolens as photosynthesis inhibitors. Molecules 2018, 23, 2693. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.S.M.; Sethi, G.; Lee, G.S.; Malek, S.N.A. Chalepin: Isolated from Ruta angustifolia L. Pers induces mitochondrial mediated apoptosis in lung carcinoma cells. BMC Complem. Altern. Med. 2016, 16, 389. [Google Scholar] [CrossRef] [Green Version]
- Ulubelen, A.; Terem, B.; Tuzlaci, E.; Cheng, K.F.; Kong, Y.C. Alkaloids and coumarins from Ruta chalepensis. Phytochemistry 1986, 25, 2692–2693. [Google Scholar] [CrossRef]
- Rollinger, J.M.; Schuster, D.; Danzl, B.; Schwaiger, S.; Markt, P.; Schmidtke, M.; Gertsch, J.; Raduner, S.; Wolber, G.; Langer, T. In silico target fishing for rationalized ligand discovery exemplified on constituents of Ruta graveolens. Planta Med. 2009, 75, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Luca, S.V.; Miron, A.; Ignatova, S.; Skalicka-Woźniak, K. An overview of the two-phase solvent systems used in the countercurrent separation of phenylethanoid glycosides and iridoids and their biological relevance. Phytochem. Rev. 2019, 18, 377–403. [Google Scholar] [CrossRef] [Green Version]
- Roehrer, S.; Behr, R.; Stork, V.; Ramires, M.; Médard, G.; Frank, O.; Kleigrewe, K.; Hofmann, T.; Minceva, M. Xanthohumol C, a minor bioactive hop compound: Production, purification strategies and antimicrobial test. J. Chromotgr. B. 2018, 1095, 39–49. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Zhang, S.L.; Ma, L.; Zhang, Y.; Shen, X.; Wang, W.; Hu, L.H. Total synthesis of (+)-rutamarin. Adv. Synth. Cat. 2008, 350, 2373–2379. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, Q.; Gu, Q.; Zhang, H.; Jiang, C.; Hu, J.; Wang, Y.; Yan, Y.; Xu, J. Semisynthesis of (-)-rutamarin derivatives and their inhibitory activity on Epstein–Barr Virus lytic replication. J. Nat. Prod. 2017, 80, 53–60. [Google Scholar] [CrossRef]
- Suhaimi, S.A.; Hong, S.L.; Malek, S.N.A. Rutamarin, an active constituent from Ruta angustifolia Pers., induced apoptotic cell death in the HT29 colon adenocarcinoma cell line. Pharmacogn. Mag. 2017, 13, S179. [Google Scholar]
- Skalicka-Woźniak, K.; Orhan, I.A.; Cordell, G.A.; Nabavi, S.M.; Budzyńska, B. Implication of coumarins towards central nervous system disorders. Pharmacol. Res. 2016, 103, 188–203. [Google Scholar] [CrossRef]
- Catto, M.; Nicolotti, O.; Leonetti, F.; Carotti, A.; Favia, A.D.; Soto-Otero, R.; Méndez-Álvarez, E.; Carotti, A. Structural insights into monoamine oxidase inhibitory potency and selectivity of 7-substituted coumarins from ligand-and target-based approaches. J. Med. Chem. 2006, 49, 4912–4925. [Google Scholar] [CrossRef]
- Binda, C.; Wang, J.; Pisani, L.; Caccia, C.; Carotti, A.; Salvati, P.; Edmondson, D.E.; Mattevi, A. Structures of human monoamine oxidase B complexes with selective noncovalent inhibitors: Safinamide and coumarin analogs. J. Med. Chem. 2017, 50, 5848–5852. [Google Scholar] [CrossRef]
- Delogu, G.; Picciau, C.; Ferino, G.; Quezada, E.; Podda, G.; Uriarte, E.; Viña, D. Synthesis, human monoamine oxidase inhibitory activity and molecular docking studies of 3-heteroarylcoumarin derivatives. Eur. J. Med. Chem. 2011, 46, 1147–1152. [Google Scholar] [CrossRef]
- Huang, M.; Xie, S.S.; Jiang, N.; Lan, J.S.; Kong, L.Y.; & Wang, X.B. Multifunctional coumarin derivatives: Monoamine oxidase B (MAO-B) inhibition, anti-β-amyloid (Aβ) aggregation and metal chelation properties against Alzheimer’s disease. Bioorg. Med. Chem. Lett. 2015, 25, 508–513. [Google Scholar] [CrossRef]
- Fonseca, A.; Reis, J.; Silva, T.; Matos, M.J.; Bagetta, D.; Ortuso, F.; Alcaro, S.; Uriarte, E.; Borges, F. Coumarin versus chromone monoamine oxidase B inhibitors: Quo vadis? J. Med. Chem. 2017, 60, 7206–7212. [Google Scholar] [CrossRef]
- Kang, S.Y.; Kim, Y.C. Neuroprotective coumarins from the root of Angelica gigas: Structure-activity relationships. Arch. Pharm. Res. 2007, 30, 1368–1373. [Google Scholar] [CrossRef]
- Kang, S.Y.; Lee, K.Y.; Sung, S.H.; Park, M.J.; Kim, Y.C. Coumarins isolated from Angelica gigas inhibit acetylcholinesterase: Structure-activity relationships. J. Nat. Prod. 2001, 64, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, D.Y.; Kim, Y.C.; Jung, J.W.; Lee, S.; Yoon, B.H.; Cheong, J.H.; Kim, Y.S.; Kang, S.S.; Ko, K.H.; et al. Nodakenin, a coumarin compound, ameliorates scopolamine-induced memory disruption in mice. Life Sci. 2007, 80, 1944–1950. [Google Scholar] [CrossRef]
- Luca, S.V.; Czerwinska, M.E.; Miron, A.; Aprotosoaie, A.C.; Marcourt, L.; Wolfender, J.L.; Granica, S.; Skalicka-Wozniak, K. High-performance countercurrent chromatographic isolation of acylated iridoid diglycosides from Verbascum ovalifolium Donn ex Sims and evaluation of their inhibitory potential on IL-8 and TNF-α production. J. Pharm. Biomed. Anal. 2019, 166, 295–303. [Google Scholar] [CrossRef]
Sample Availability: Samples of the rutamarin and R.graveolens dichloromethane extract are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Concentration | %INH(hMAO-A) | %INH(hMAO-B) | SI 1 |
|---|---|---|---|---|
| Rutamarin | 6.17 μM | 25.15 | 95.26 | 3.788 |
| R. graveolens extract | 9.78 mg/mL | 88.22 | 89.98 | 1.020 |
| Reference selective MAO-A or MAO-B inhibitors | ||||
| Selegiline | 5.34 μM | – | 99.07 | – |
| Clorgiline | 3.67 μM | 99.29 | – | – |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozioł, E.; Luca, S.V.; Ağalar, H.G.; Sağlık, B.N.; Demirci, F.; Marcourt, L.; Wolfender, J.-L.; Jóźwiak, K.; Skalicka-Woźniak, K. Rutamarin: Efficient Liquid–Liquid Chromatographic Isolation from Ruta graveolens L. and Evaluation of Its In Vitro and In Silico MAO-B Inhibitory Activity. Molecules 2020, 25, 2678. https://doi.org/10.3390/molecules25112678
Kozioł E, Luca SV, Ağalar HG, Sağlık BN, Demirci F, Marcourt L, Wolfender J-L, Jóźwiak K, Skalicka-Woźniak K. Rutamarin: Efficient Liquid–Liquid Chromatographic Isolation from Ruta graveolens L. and Evaluation of Its In Vitro and In Silico MAO-B Inhibitory Activity. Molecules. 2020; 25(11):2678. https://doi.org/10.3390/molecules25112678
Chicago/Turabian StyleKozioł, Ewelina, Simon Vlad Luca, Hale Gamze Ağalar, Begüm Nurpelin Sağlık, Fatih Demirci, Laurence Marcourt, Jean-Luc Wolfender, Krzysztof Jóźwiak, and Krystyna Skalicka-Woźniak. 2020. "Rutamarin: Efficient Liquid–Liquid Chromatographic Isolation from Ruta graveolens L. and Evaluation of Its In Vitro and In Silico MAO-B Inhibitory Activity" Molecules 25, no. 11: 2678. https://doi.org/10.3390/molecules25112678
APA StyleKozioł, E., Luca, S. V., Ağalar, H. G., Sağlık, B. N., Demirci, F., Marcourt, L., Wolfender, J. -L., Jóźwiak, K., & Skalicka-Woźniak, K. (2020). Rutamarin: Efficient Liquid–Liquid Chromatographic Isolation from Ruta graveolens L. and Evaluation of Its In Vitro and In Silico MAO-B Inhibitory Activity. Molecules, 25(11), 2678. https://doi.org/10.3390/molecules25112678