
Tannin-Rich Plants as Natural Manipulators of Rumen Fermentation in the Livestock Industry

 , , and
, , and
Abstract
:1. Introduction
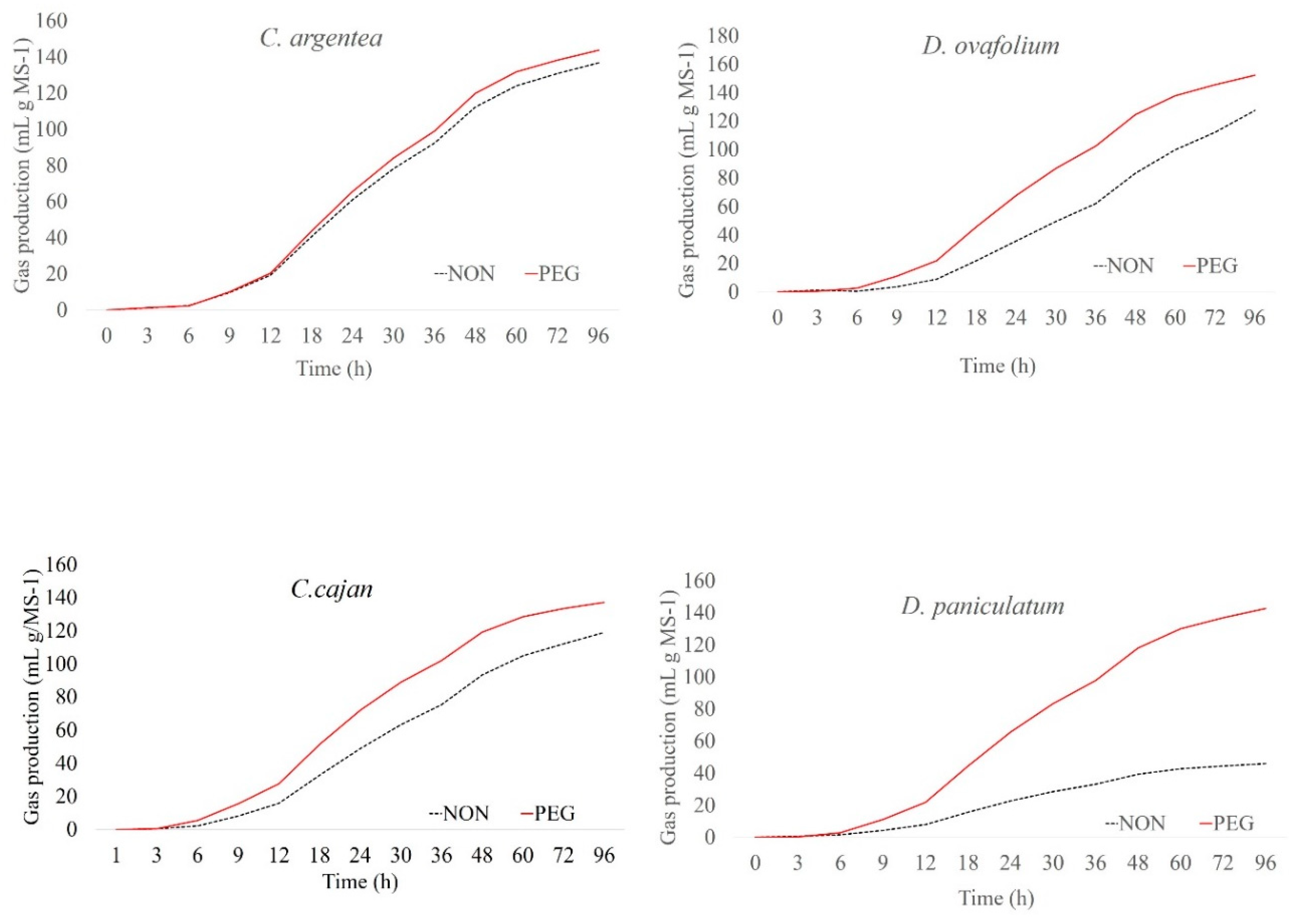
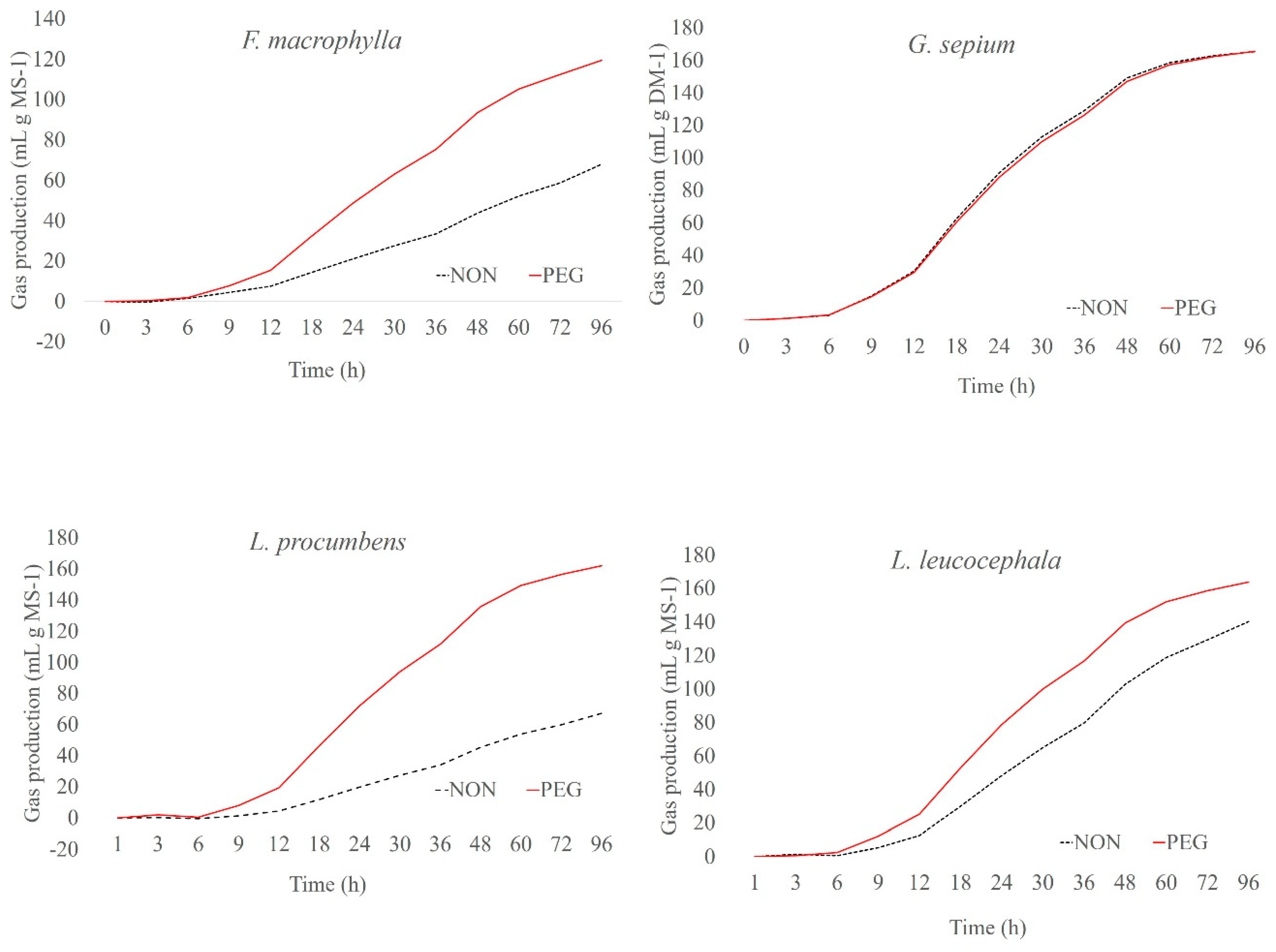
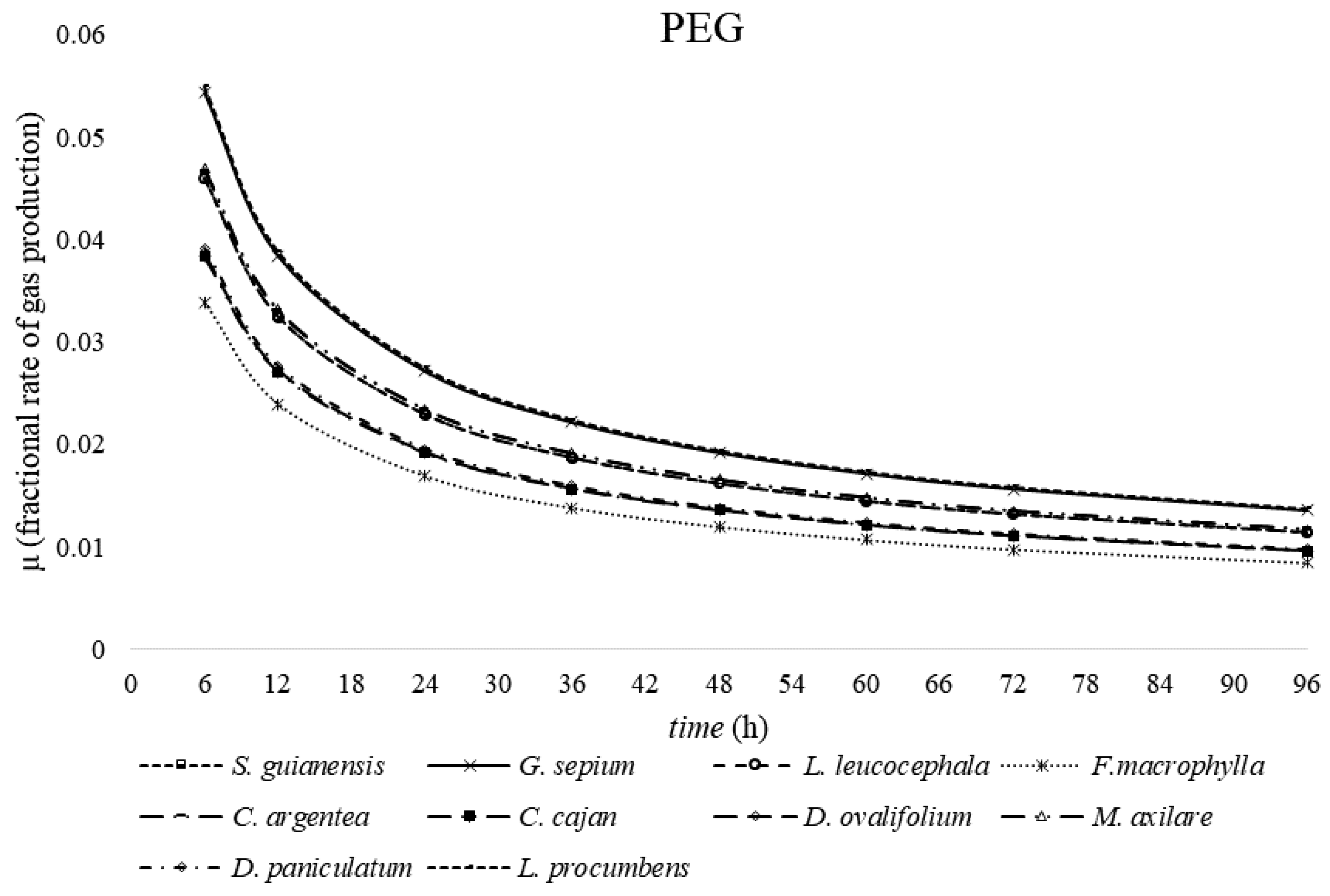
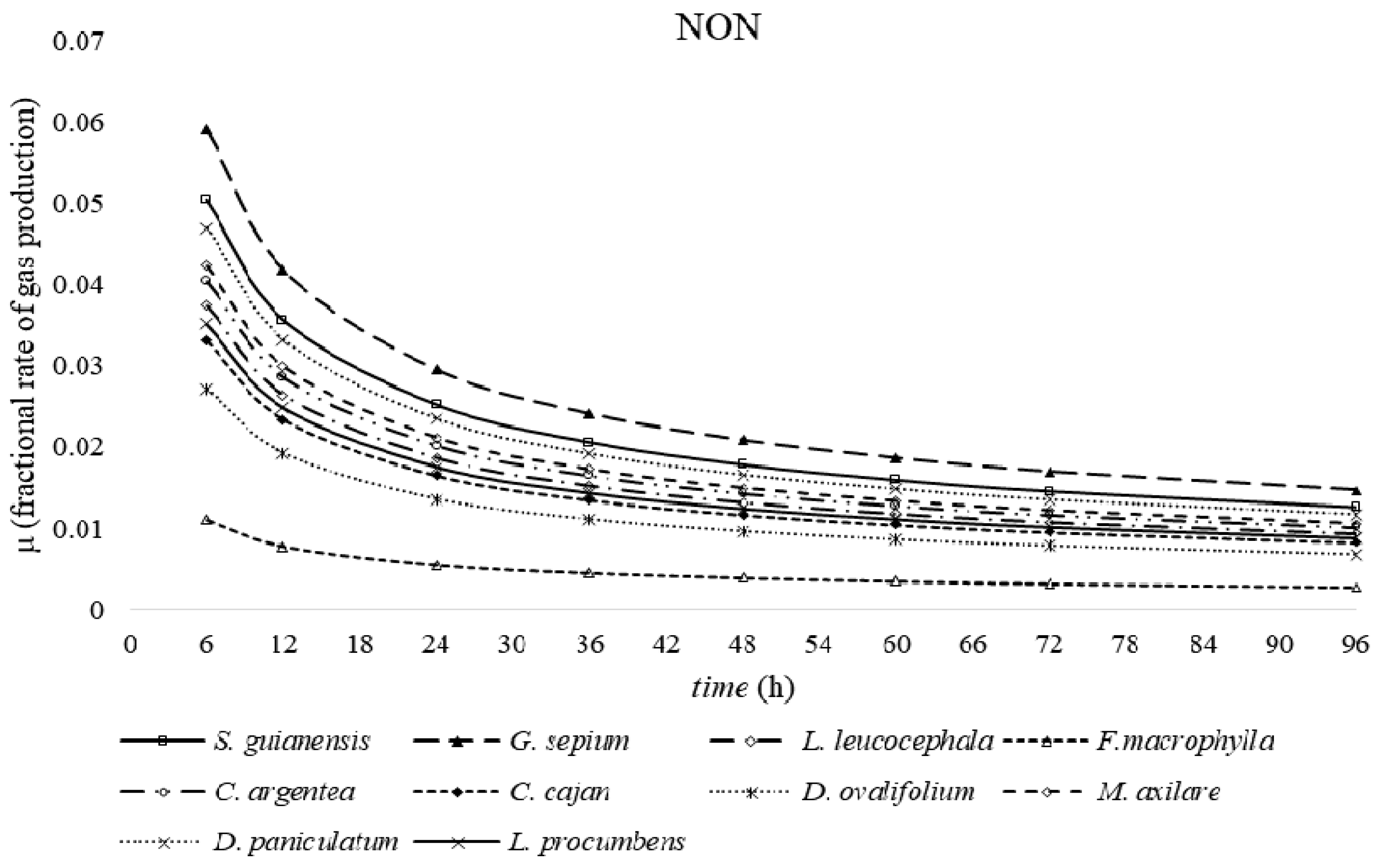
2. Results
3. Discussion
4. Materials and Methods
4.1. Tannin-Rich Plants
4.2. Chemical Analysis
4.3. Animals and Inoculum Preparation
4.4. In Vitro Gas Production Assay
4.5. Short-Chain Fatty Acid Determination
4.6. N-NH3 Determination
4.7. Experimental Design and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valle, C.B.; Jank, L.; Resende, R.M.S. O melhoramento de forrageiras tropicais no Brasil. Rev Ceres. 2009, 56, 460–472. [Google Scholar]
- Bueno, I.C.S.; Brandi, R.A.; Franzolin, R.; Fagundes, G.M.; Benetel, G.; Abdalla, A.L.; Louvandini, H.; Muir, J.P. Comparative in vitro methane production and antimethanogenic effect of tannins in five ruminant species. Anim. Feed Sci. Tech. 2015, 205, 1–9. [Google Scholar] [CrossRef]
- Fagundes, G.M.; Benetel, G.; Welter, K.C.; Melo, F.A.; Muir, J.P.; Carriero, M.M.; Souza, R.L.M.; Meo-Filho, P.; Frighetto, R.T.S.; Berndt, A.; et al. Tannin as a natural rumen modifier to control methanogenesis in beef cattle in tropical systems: Friend or foe to biogas energy production? Res. Vet. Sci. 2020, 132, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.O.C.A.; Ramírez-Restrepo, C.A.; Muir, J.P. Developing a conceptual model of possible benefits of condensed tannins for ruminant production. Anim. 2014, 8, 1095–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, A.B.; Crossland, W.L.; Tedeschi, L.O.; Foster, J.L.; Muir, J.P.; Pinchak, W.E.; Fonseca, M.A. Inclusion of quebracho tannin extract in a high-roughage cattle diet alters digestibility, nitrogen balance, and energy partitioning. J. Anim. Sci. 2020. (at press). [Google Scholar] [CrossRef]
- Fagundes, G.M.; Modesto, E.C.; Fonseca, C.E.M.; Lima, H.R.P.; Muir, J.P. Intake, digestibility and milk yield in goats fed Flemingia macrophylla with or without polyethylene glycol. Small Rumin. Res. 2014, 116, 88–93. [Google Scholar] [CrossRef]
- Muir, J.P.; Tedeschi, L.O.; Dubeux, J.C.B., Jr.; Peters, M.; Burkart, S. Enhancing food security in Latin America with forage legumes. Archivos Latinoamericanos Producción Anim. 2017, 25, 113–131. [Google Scholar]
- Santos, K.C.; Magalhães, A.L.R.; Silva, D.K.A.; Araújo, G.G.L.; Fagundes, G.M.; Ybarra, N.G.; Abdalla, A.L. Nutritional potential of forage species found in Brazilian Semiarid region. Livest. Sci. 2017, 195, 118–124. [Google Scholar] [CrossRef]
- Naumann, H.D.; Armstrong, S.A.; Lambert, B.D.; Muir, J.P.; Tedeschi, L.O.; Kothmann, M.M. Effect of molecular weight and concentration of legume condensed tannins on in vitro larval migration inhibition of Haemonchus contortus. J. Vet. Parasitol. 2014, 199, 93–98. [Google Scholar] [CrossRef]
- France, J.; Dhanoa, M.S.; Theodorou, M.K.; Lister, S.J.; Davies, D.R.; Isac, D. A model to interpret gas accumulation profiles associated with in vitro degradation of ruminant feeds. J. Theor. Biol. 1993, 163, 99–111. [Google Scholar] [CrossRef]
- Bueno, I.C.S.; Vitti, D.M.S.S.; Louvandini, H.; Abdalla, A.L. A new approach for in vitro bioassay to measure tannin biological effects based on a gas production technique. Anim. Feed Sci. Tech. 2008, 141, 153–170. [Google Scholar] [CrossRef]
- Baba, A.S.H.; Castro, F.B.; Ørskov, E.R. Partitioning of energy and degradability of browse plants in vitro and the implications of blocking the effects of tannin by the addition of polyethylene glycol. Anim. Feed Sci. Technol. 2002, 95, 93–104. [Google Scholar] [CrossRef]
- Gemeda, B.S.; Hassen, A. Effect of tannin and species variation on in vitro digestibility, gas, and methane production of tropical browse plants. Asian Australas. J. Anim. Sci. 2015, 28, 188–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatta, R.; Uyeno, Y.; Tajima, K.; Takenaka, A.; Yabumoto, Y.; Nonaka, I.; Kurihara, M. Difference in the nature of tannins on in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations. J. Dairy Sci. 2009, 92, 5512–5522. [Google Scholar] [CrossRef] [Green Version]
- Bhatta, R.; Baruah, L.; Saravanan, M.; Suresh, K.P.; Sampath, K.T. Effect of medicinal and aromatic plants on rumen fermentation, protozoa population and methanogenesis in vitro. J. Anim. Physiol. Anim. Nutr. 2013, 97, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef]
- Frutos, P.; Hervas, G.; Giraldez, F.J.; Mantecon, A.R. Review. Tannins and ruminant nutrition. Span. J. Agric. Res. 2004, 2, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Kamalak, A.D.E.M.; Canbolat, O.; Sahin, M.; Gurbuz, Y.; Ozkose, E.; Ozkan, C.O. The effect of polyethylene glycol (PEG 8000) supplementation on in vitro gas production kinetics of leaves from tannin containing trees. S. Afr. J. Anim. Sci. 2005, 35, 229. [Google Scholar] [CrossRef]
- Calabrò, S.; Guglielmelli, A.; Iannaccone, F.; Danieli, P.P.; Tudisco, R.; Ruggiero, C.; Piccolo, G.; Cutrignell, M.I.; Infascelli, F. Fermentation kinetics of sainfoin hay with and without PEG. J. Anim. Physiol. Anim. Nutr. 2012, 96, 842–848. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 16th ed.; AOAC: Arlington, VA, USA, 1995. [Google Scholar]
- Mertens, D.R. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beaker or crucibles: Collaborative study. J. AOAC Int. 2002, 85, 1217–1240. [Google Scholar]
- Robertson, J.B.; Van Soest, P.J. The detergent system of analysis. In The Analysis of Dietary Fibre in Food; James, W.P.T., Theander, O., Eds.; Marcel Dekker: New York, NY, USA, 1981; pp. 123–158. [Google Scholar]
- Makkar, H.P.S. Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Rumin. Res. 2003, 49, 241–256. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Blümmel, M.; Borowy, N.K.; Becker, K. Gravimetric determination of tannins and their correlations with chemical and protein precipitation methods. J. Sci. Food Agric. 1993, 61, 161–165. [Google Scholar] [CrossRef]
- Terrill, T.H.; Rowan, A.M.; Douglas, G.B.; Barry, T.N. Determination of extractable and bound condensed tannin concentrations in forage plants, protein concentrate meals and cereal grains. J. Sci. Food Agric. 1992, 58, 321–329. [Google Scholar] [CrossRef]
- Wolfe, R.M.; Terrill, T.H.; Muir, J.P. Drying method and origin of standard affect condensed tannin (CT) concentrations in perennial herbaceous legumes using simplified butanol-HCl CT analysis. J. Sci. Food Agric. 2008, 88, 1060–1067. [Google Scholar] [CrossRef]
- Bueno, I.C.S.; Cabral Filho, S.L.S.; Gobbo, S.P.; Louvandini, H.; Vitti, D.M.S.S.; Aballa, A.L. Influence of inoculum source in a gas production method. Anim. Feed Sci. Technol. 2005, 123–124, 96–105. [Google Scholar] [CrossRef]
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; Mcallan, A.B.; France, J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed Sci. Tech. 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Mauricio, R.M.; Mould, F.L.; Dhanoa, M.S.; Owen, E.; Channa, K.S.; Theodorou, M.K. A semi-automated in vitro gas production technnique for ruminant feedstuff evaluation. Anim. Feed Sci. Technol. 1999, 79, 321–330. [Google Scholar] [CrossRef]
- Onodera, R.; Henderson, C. Growth factors of bacterial origin for the culture of the rumen oligotrich protozoon, Entodinium caudatum. J. Appl. Bacteriol. 1980, 48, 125–134. [Google Scholar] [CrossRef]
- Blümmel, M.; Makkar, H.P.S.; Becker, K. In vitro gas production: A technique revisited. J. Anim. Physiol. Anim. Nutr. 1997, 77, 24–34. [Google Scholar] [CrossRef]
- Bueno, I.C.S.; Brandi, R.A.; Fagundes, G.M.; Benetel, G.; Muir, J.P. The role of condensed tannins in the in vitro rumen fermentation kinetics in ruminant species: Feeding type involved? Animals 2020, 10, 635. [Google Scholar] [CrossRef] [Green Version]
- Preston, T.R. Biological and chemical analytical methods. In Tropical Animal Feeding: A Manual for Research Workers; FAO: Rome, Italy, 1995; pp. 191–264. [Google Scholar]
- Sas Institute. The SAS System for Windows. Release 8.01; SAS Institute: Cary, NC, USA, 2000. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables/Plant | |||
|---|---|---|---|
| IVDMD (g/kg DM) | Without PEG | With PEG | Plant × PEG (2) |
| Cajanus cajan | 403 §D | 435 DE | ns |
| Cynodon spp (Tifton 85) | 751 A | 739 A | ns |
| Cratylia argentea | 553 aC | 458 bD | *** |
| Desmodium ovalifolium | 432 D | 474 D | ns |
| Desmodium paniculatum | 278 bF | 336 aF | * |
| Flemingia macrophylla | 300 bF | 362 aF | * |
| Gliricidia sepium | 676 B | 633 C | ns |
| Lespedeza procumbens | 352 bE | 406 aE | * |
| Leucaena leucocephala | 567 bC | 633 aC | * |
| Medicago sativa | 697 B | 703 BC | ns |
| Macrotiloma axilare | 670 B | 671 BC | ns |
| Stylosanthes guianensis | 694 B | 695 AB | ns |
| SEM(1) | 2.42 | 2.10 | 1.81 |
| p-values (2) | *** | *** | |
| IVOMD (g/kg DM) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 406 §D | 438 CDE | ns |
| Cynodon spp (Tifton 85) | 753 A | 748 A | ns |
| Cratylia argentea | 554 C | 502 C | ns |
| Desmodium ovalifolium | 429 D | 471CD | ns |
| Desmodium paniculatum | 273 bF | 409 aDE | *** |
| Flemingia macrophylla | 298 F | 357 E | ns |
| Gliricidia sepium | 685 B | 637 B | ns |
| Lespedeza procumbens | 357 E | 409 DE | ns |
| Leucaena leucocephala | 557 bC | 628 aB | * |
| Medicago sativa | 689 B | 697 AB | ns |
| Macrotiloma axilare | 672 B | 673 AB | ns |
| Stylosanthes guianensis | 700 B | 702 AB | ns |
| SEM(1) | 2.44 | 2.05 | 2.35 |
| p-values (2) | *** | *** | |
| Partitioning factor (mg DMD/mL) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 3.09 §BC | 2.84 C | ns |
| Cynodon spp (Tifton 85) | 2.73 C | 2.64 C | ns |
| Cratylia argentea | 3.82 aB | 2.87 bC | ** |
| Desmodium ovalifolium | 3.11 BC | 2.81 C | ns |
| Desmodium paniculatum | 5.50 aA | 2.14 bD | *** |
| Flemingia macrophylla | 3.86 aB | 2.73 bC | *** |
| Gliricidia sepium | 3.61 BC | 3.39 AB | ns |
| Lespedeza procumbens | 4.74 aA | 2.28 bD | *** |
| Leucaena leucocephala | 3.73 B | 3.49 A | ns |
| Medicago sativa | 3.27 BC | 3.32 AB | ns |
| Macrotiloma axilare | 3.46 BC | 3.36 AB | ns |
| Stylosanthes guianensis | 3.22 BC | 3.18 B | ns |
| SEM(1) | 0.13 | 0.07 | 0.24 |
| p-values (2) | *** | *** | |
| NNH3 (mg/100 mL) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 22.70 §CD | 27.87 AB | ns |
| Cynodon spp (Tifton 85) | 22.12 CD | 19.37 D | ns |
| Cratylia argentea | 25.87 BCD | 29.87 AB | ns |
| Desmodium ovalifolium | 12.75 bF | 20.50 aCD | * |
| Desmodium paniculatum | 15.50 bF | 30.25 aAB | *** |
| Flemingia macrophylla | 17.25 bEF | 27.87 aAB | *** |
| Gliricidia sepium | 30.50 AB | 34.00 A | ns |
| Lespedeza procumbens | 12.37 bF | 24.75 aBCD | *** |
| Leucaena leucocephala | 21.25 bDE | 29.12 aAB | * |
| Medicago sativa | 32.12 A | 26.62 BC | ns |
| Macrotiloma axilare | 25.62 BCD | 23.37 BCD | ns |
| Stylosanthes guianensis | 26.87 BC | 27.75 AB | ns |
| SEM(1) | 0.82 | 1.07 | 2.49 |
| p-values (2) | *** | *** |
| Short-Chain Fatty Acid/Plant | |||
|---|---|---|---|
| Acetic Acid (mmol/g OMD) | Without PEG | With PEG | Plant × PEG (2) |
| Cajanus cajan | 8.42 §AB | 8.43 BC | ns |
| Cynodon spp (Tifton 85) | 6.71 B | 6.96 C | ns |
| Cratylia argentea | 6.74 B | 7.55 C | ns |
| Desmodium ovalifolium | 8.44 AB | 8.03 BC | ns |
| Desmodium paniculatum | 8.56 bAB | 10.50 aA | ** |
| Flemingia macrophylla | 9.45 A | 9.70 AB | ns |
| Gliricidia sepium | 7.20 B | 7.52 C | ns |
| Lespedeza procumbens | 7.48 bAB | 10.48 aA | *** |
| Leucaena leucocephala | 7.61 AB | 6.90 C | ns |
| Medicago sativa | 7.52 AB | 7.15 C | ns |
| Macrotiloma axilare | 6.59 B | 6.77 C | ns |
| Stylosanthes guianensis | 7.45 AB | 7.41 C | ns |
| SEM(1) | 0.25 | 0.17 | 0.56 |
| p-values (2) | *** | *** | |
| Propionic Acid (mmol/g OMD) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 2.35 §ABC | 2.33 BC | ns |
| Cynodon spp (Tifton 85) | 2.29 ABC | 2.54 AB | ns |
| Cratylia argentea | 2.23 ABC | 2.78 AB | ns |
| Desmodium ovalifolium | 1.75 BC | 2.29 BC | ns |
| Desmodium paniculatum | 1.98 BC | 2.63 AB | ns |
| Flemingia macrophylla | 2.55 AB | 2.85 A | ns |
| Gliricidia sepium | 2.95 A | 3.01 A | ns |
| Lespedeza procumbens | 1.56 bC | 3.02 aA | *** |
| Leucaena leucocephala | 2.62 AB | 2.30 BC | ns |
| Medicago sativa | 1.99 BC | 1.93 C | ns |
| Macrotiloma axilare | 2.33 ABC | 2.31 BC | ns |
| Stylosanthes guianensis | 1.96 BC | 1.95 C | ns |
| SEM(1) | 0.09 | 0.07 | 0.24 |
| p-values (2) | * | *** | |
| Butyric acid (mmol/g OMD) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 0.92 §ABC | 1.09 AB | ns |
| Cynodon spp (Tifton 85) | 1.05 A | 1.11 AB | ns |
| Cratylia argentea | 0.77 DE | 0.89 CDE | ns |
| Desmodium ovalifolium | 0.73 bDEF | 0.94 aBCD | * |
| Desmodium paniculatum | 0.57 bFG | 1.14 aA | *** |
| Flemingia macrophylla | 0.70 bDEF | 1.08 aAB | *** |
| Gliricidia sepium | 0.73 DEF | 0.77 DE | ns |
| Lespedeza procumbens | 0.45 bG | 1.20 aA | *** |
| Leucaena leucocephala | 0.64 EF | 0.74 E | ns |
| Medicago sativa | 1.00 AB | 0.96 BC | ns |
| Macrotiloma axilare | 0.86 BCD | 0.87 CDE | ns |
| Stylosanthes guianensis | 0.82 DC | 0.79 CDE | ns |
| SEM(1) | 0.03 | 0.03 | 0.06 |
| p-values (2) | *** | *** | |
| Isobutyric acid (mmol/g OMD) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 0.19 §bABC | 0.26 aCDE | * |
| Cynodon spp (Tifton 85) | 0.14 BCD | 0.14 F | ns |
| Cratylia argentea | 0.21 ABC | 0.25 CDE | ns |
| Desmodium ovalifolium | 0.17 ABCD | 0.19 EF | ns |
| Desmodium paniculatum | n.d b | 0.37 aA | *** |
| Flemingia macrophylla | 0.12 bCD | 0.35 aAB | *** |
| Gliricidia sepium | 0.26 A | 0.26 CDE | ns |
| Lespedeza procumbens | 0.06 bD | 0.33 aABC | *** |
| Leucaena leucocephala | 0.16 bABCD | 0.29 aBCD | * |
| Medicago sativa | 0.25 AB | 0.24 DE | ns |
| Macrotiloma axilare | 0.19 ABC | 0.21 DEF | ns |
| Stylosanthes guianensis | 0.19 ABC | 0.19 EF | ns |
| SEM(1) | 0.01 | 0.01 | 0.04 |
| p-values (2) | * | *** | |
| Isovaleric acid (mmol/g OMD) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 0.22§bCD | 0.36 aBCDE | *** |
| Cynodon spp (Tifton 85) | 0.17 DE | 0.18 H | ns |
| Cratylia argentea | 0.31 B | 0.34 CDEF | ns |
| Desmodium ovalifolium | 0.14 bDEF | 0.24 bGH | *** |
| Desmodium paniculatum | 0.08 bF | 0.52 aA | *** |
| Flemingia macrophylla | 0.12 bEF | 0.44 aB | *** |
| Gliricidia sepium | 0.29 BC | 0.30 DEFG | ns |
| Lespedeza procumbens | 0.08 bF | 0.41 aBC | *** |
| Leucaena leucocephala | 0.16 bDE | 0.30 aDEFG | *** |
| Medicago sativa | 0.39 A | 0.37 BCD | ns |
| Macrotiloma axilare | 0.26 BC | 0.28 EFG | ns |
| Stylosanthes guianensis | 0.28 BC | 0.27 FG | ns |
| SEM(1) | 0.02 | 0.01 | 0.03 |
| p-values (2) | *** | *** | |
| Valeric acid (mmol/g OMD) | Without PEG | With PEG | Plant × PEG |
| Cajanus cajan | 0.22 §bAB | 0.28 aB | * |
| Cynodon spp (Tifton 85) | 0.18 BC | 0.19 DE | ns |
| Cratylia argentea | 0.17 CD | 0.20 DE | ns |
| Desmodium ovalifolium | 0.14 bD | 0.19 aE | * |
| Desmodium paniculatum | 0.08 bE | 0.23 aBCD | *** |
| Flemingia macrophylla | 0.19 bBC | 0.31 aA | *** |
| Gliricidia sepium | 0.20 BC | 0.21 CDE | ns |
| Lespedeza procumbens | 0.08 bE | 0.23 aBCDE | *** |
| Leucaena leucocephala | 0.13 bD | 0.21 aCDE | *** |
| Medicago sativa | 0.21 ABC | 0.21 CDE | ns |
| Macrotiloma axilare | 0.22 AB | 0.22 BCDE | ns |
| Stylosanthes guianensis | 0.24 A | 0.25 BC | ns |
| SEM(1) | 0.01 | 0.01 | 0.01 |
| p-values (2) | *** | *** |
| Model Parameters | A (mL) | L (h/min) | T/2 (h) | |||
|---|---|---|---|---|---|---|
| NON | PEG (1) | NON | PEG | NON | PEG | |
| Cajanus cajan | 122.87 §bF | 138.50 aF | 3.66 B | 3.30 A | 29.20 CD | 23.10 D |
| Cynodon spp | 254.52 bA | 258.82 aA | 4.10 AB | 4.00 A | 29.56 CD | 29.81 A |
| Cratylia argentea | 139.45 bE | 146.15 aF | 3.55 B | 4.74 A | 27.17 DE | 26.30 BC |
| Desmodium ovalifolium | 143.12 bE | 155.60 aE | 4.84 AB | 4.18 A | 41.45 aAB | 27.16 bB |
| Desmodium paniculatum | 46.43 bH | 145.40 aF | 4.47 AB | 4.53 A | 24.53 DEF | 26.37 BC |
| Flemingia macrophylla | 81.99 bG | 123.27 aG | 3.87 B | 4.25 A | 44.92 aA | 29.42 bA |
| Gliricidia sepium | 165.92 D | 166.42 D | 3.92 B | 4.47 A | 22.25 EF | 22.90 D |
| Lespedeza procumbens | 75.60 bG | 163.85 aD | 6.21 bA | 3.86 aA | 39.42 aB | 26.62 bBC |
| Leucaena leucocephala | 147.32 bE | 165.47 aD | 4.72 AB | 4.25 A | 33.49 C | 25.11 C |
| Medicago sativa | 188.01 aBC | 187.15 bC | 3.04 B | 3.41 A | 20.85 F | 21.34 E |
| Macrotiloma axilare | 175.25 bCD | 180.95 aC | 3.53 aB | 3.86 A | 23.39 EF | 23.61 D |
| Stylosanthes guianensis | 192.92 bB | 197.45 aB | 3.38 aB | 3.61 A | 20.21 F | 20.72 E |
| p-values (2) | *** | *** | *** | *** | *** | *** |
| Plant | CP | EE | NDF | ADF | ADL | TP (1) | TT (1) | CT (2) |
|---|---|---|---|---|---|---|---|---|
| Cajanus cajan | 200 | 38 | 552 | 438 | 191 | 50 | 39 | 34 |
| Cynodon spp (Tifton 85) | 138 | 13 | 743 | 306 | 133 | n.d §. | n.d | n.d |
| Cratylia argentea | 195 | 18 | 577 | 418 | 164 | 36 | 21 | 1.0 |
| Desmodium ovalifolium | 112 | 12 | 614 | 490 | 176 | 237 | 223 | 164 |
| Desmodium paniculatum | 162 | 20 | 515 | 414 | 216 | 171 | 154 | 70 |
| Flemingia macrophylla | 189 | 16 | 577 | 465 | 229 | 181 | 168 | 109 |
| Gliricidia sepium | 281 | 23 | 441 | 277 | 130 | 45 | 28 | 0.3 |
| Lespedeza procumbens | 141 | 20 | 475 | 349 | 161 | 108 | 87 | 198 |
| Leucaena leucocephala | 263 | 22 | 444 | 271 | 136 | 258 | 229 | 48 |
| Medicago sativa | 271 | 84 | 448 | 245 | 127 | 14 | 8 | 0.2 |
| Macrotiloma axilare | 208 | 22 | 523 | 377 | 105 | 30 | 24 | 1.2 |
| Stylosanthes guianensis | 212 | 31 | 509 | 350 | 95 | 63 | 41 | 6.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fagundes, G.M.; Benetel, G.; Santos, K.C.; Welter, K.C.; Melo, F.A.; Muir, J.P.; Bueno, I.C.S. Tannin-Rich Plants as Natural Manipulators of Rumen Fermentation in the Livestock Industry. Molecules 2020, 25, 2943. https://doi.org/10.3390/molecules25122943
Fagundes GM, Benetel G, Santos KC, Welter KC, Melo FA, Muir JP, Bueno ICS. Tannin-Rich Plants as Natural Manipulators of Rumen Fermentation in the Livestock Industry. Molecules. 2020; 25(12):2943. https://doi.org/10.3390/molecules25122943
Chicago/Turabian StyleFagundes, G. M., G. Benetel, K. C. Santos, K. C. Welter, F. A. Melo, J. P. Muir, and I. C. S. Bueno. 2020. "Tannin-Rich Plants as Natural Manipulators of Rumen Fermentation in the Livestock Industry" Molecules 25, no. 12: 2943. https://doi.org/10.3390/molecules25122943
APA StyleFagundes, G. M., Benetel, G., Santos, K. C., Welter, K. C., Melo, F. A., Muir, J. P., & Bueno, I. C. S. (2020). Tannin-Rich Plants as Natural Manipulators of Rumen Fermentation in the Livestock Industry. Molecules, 25(12), 2943. https://doi.org/10.3390/molecules25122943