
Effects of Biosurfactants on Enzymatic Saccharification and Fermentation of Pretreated Softwood





Abstract
:1. Introduction
2. Results
2.1. Effects of Rhamnolipid and Escin on Saccharification of Avicel in the Presence of Lignin
2.2. Effects of Rhamnolipid and Saponins on Saccharification of Steam-Pretreated Spruce
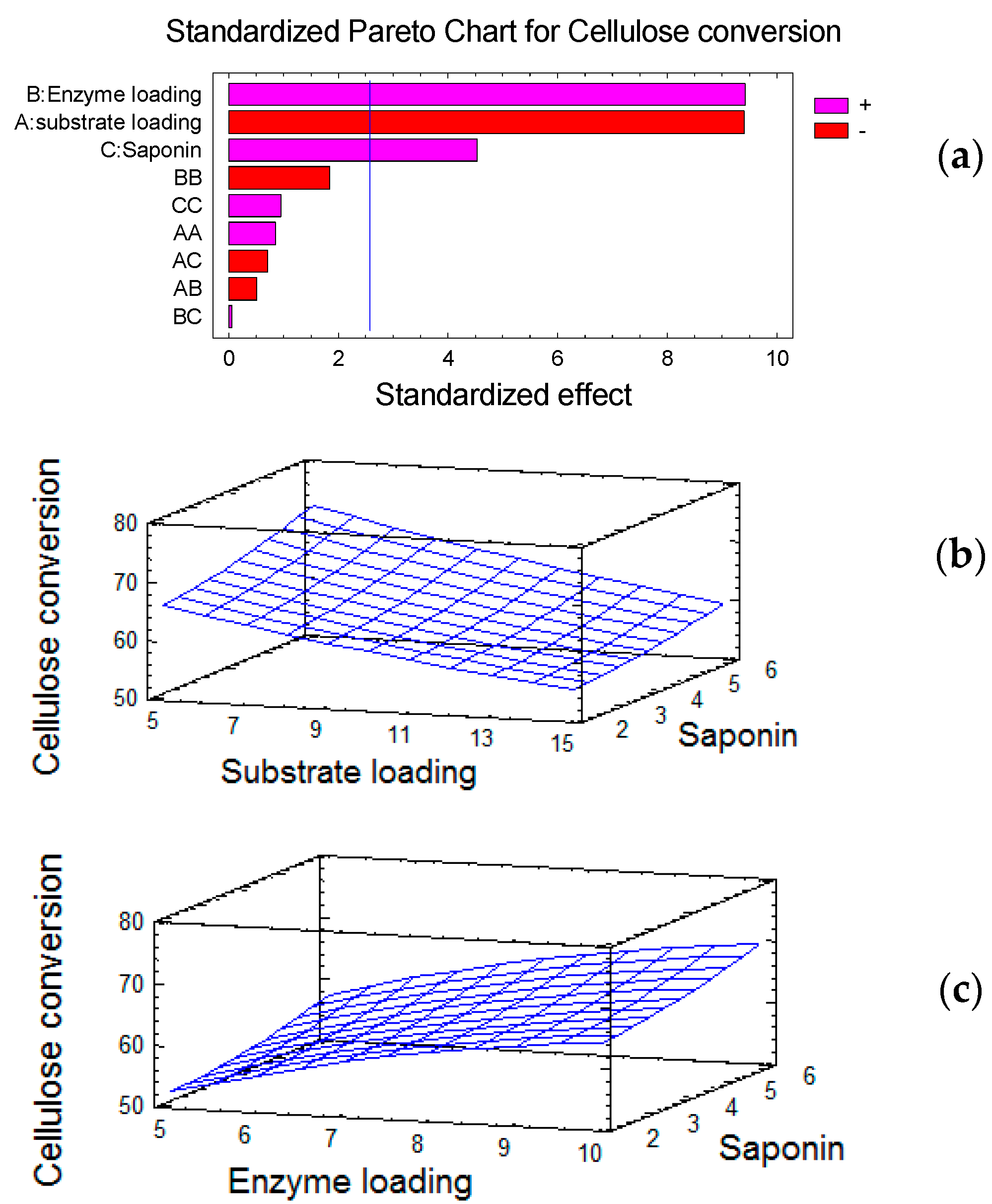
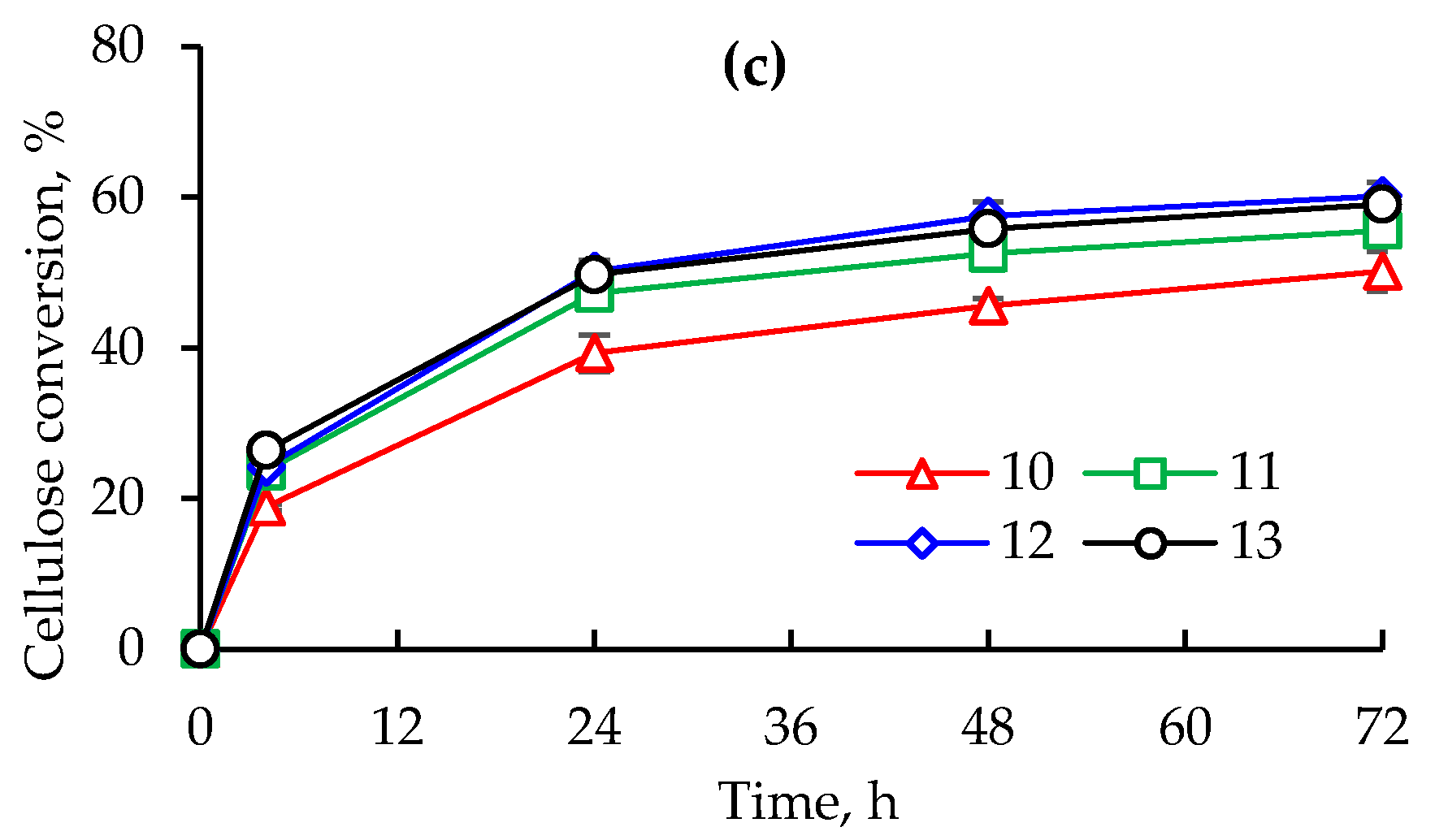
2.3. Evaluation of Red Quinoa Saponins (RQS) Dosage during Enzymatic Saccharification
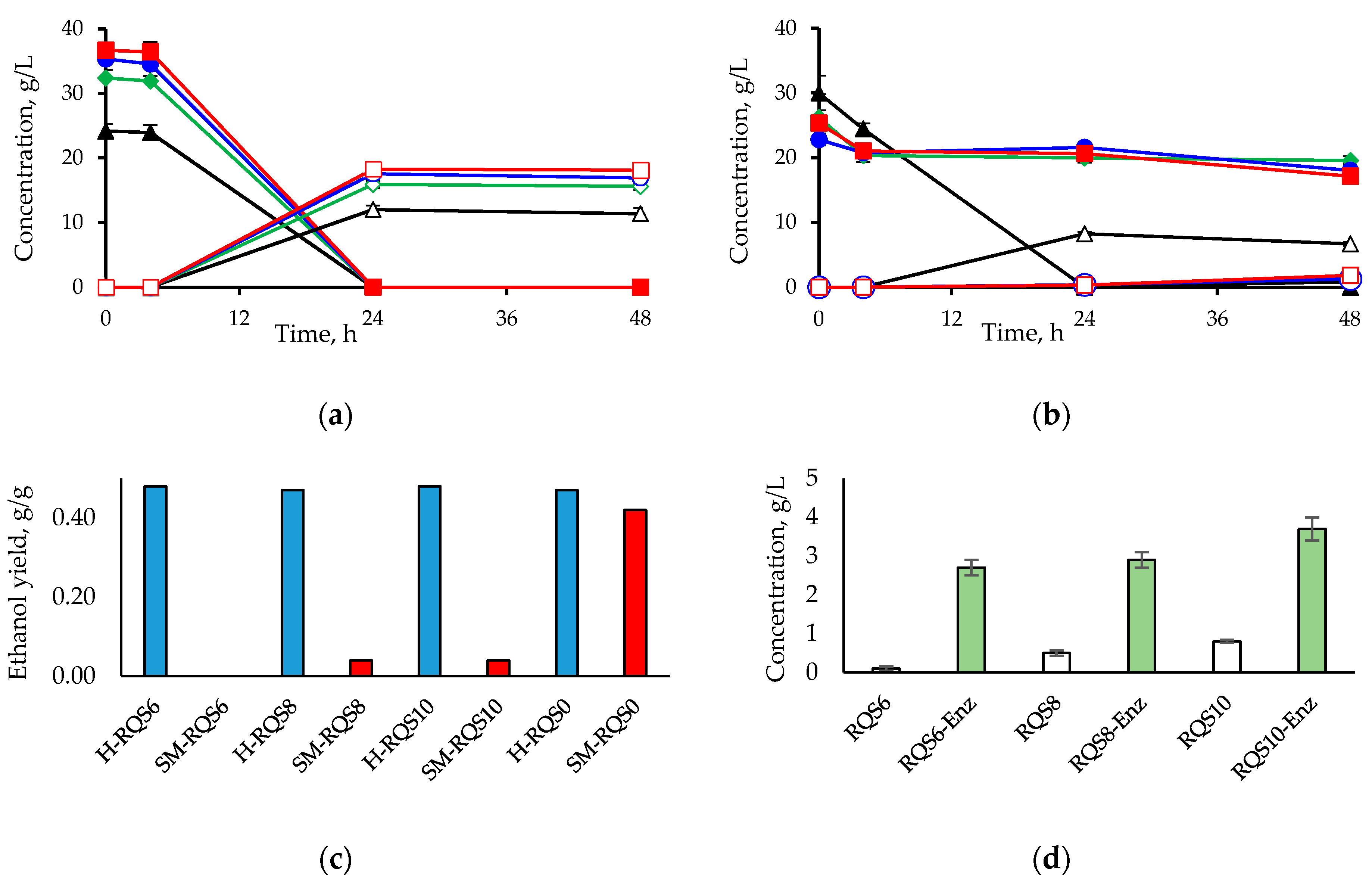
2.4. Effect of Red Quinoa Saponins (RQS) on Ethanol Production by Saccharomyces cerevisiae
3. Discussion
3.1. Effect of Biosurfactants on Enzymatic Hydrolysis of Avicel-Lignin Mixtures and Steam-Pretreated Spruce
3.2. Evaluation of Red Quinoa Saponins (RQS) Dosage during Enzymatic Saccharification
3.3. Effect of Red Quinoa Saponins (RQS) on Ethanol Production by Saccharomyces cerevisiae
4. Materials and Methods
4.1. Pretreatment of Material
4.2. Enzymes and Chemicals
4.3. Microorganism and Media
4.4. Enzymatic Saccharification of Avicel in Presence of Lignin and Biosurfactants
4.5. Enzymatic Saccharification of Steam-Pretreated Solids in Presence of Biosurfactants
4.6. Assessing the Red Quinoa Saponins (RQS) Dosage on Enzymatic Hydrolysis
4.7. Fermentation of Synthetic Medium and Hydrolysate in Presence of Red Quinoa Saponins (RQS)
4.8. Analytical Methods
4.9. Formatting of Chemical Structures
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schwanninger, M.; Rodrigues, J.C.; Gierlinger, N.; Hinterstoisser, B. Determination of lignin content in Norway spruce wood by Fourier transformed near infrared spectroscopy and partial least squares regression analysis. Part 2: Development and evaluation of the final model. J. Near. Infrared Spectrosc. 2011, 19, 331–341. [Google Scholar] [CrossRef]
- Rahikainen, J.; Mikander, S.; Marjamaa, K.; Tamminen, T.; Lappas, A.; Viikari, L.; Kruus, K. Inhibition of enzymatic hydrolysis by residual lignins from softwood—Study of enzyme binding and inactivation on lignin-rich surface. Biotechnol. Bioeng. 2011, 108, 2823–2834. [Google Scholar] [CrossRef] [PubMed]
- Vermaas, J.V.; Petridis, L.; Qi, X.; Schulz, R.; Lindner, B.; Smith, J.C. Mechanism of lignin inhibition of enzymatic biomass deconstruction. Biotechnol. Biofuels 2015, 8, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palonen, H.; Tjerneld, F.; Zacchi, G.; Tenkanen, M. Adsorption of Trichoderma reesei CBH I and EG II and their catalytic domains on steam pretreated softwood and isolated lignin. J. Biotechnol. 2004, 107, 65–72. [Google Scholar] [CrossRef]
- Xu, F.; Ding, H.; Osborn, D.; Tejirian, A.; Brown, K.; Albano, W.; Sheehy, N.; Langston, J. Partition of enzymes between the solvent and insoluble substrate during the hydrolysis of lignocellulose by cellulases. J. Mol. Catal. B-Enzym. 2008, 51, 42–48. [Google Scholar] [CrossRef]
- Eriksson, T.; Börjesson, J.; Tjerneld, F. Mechanism of surfactant effect in enzymatic hydrolysis of lignocellulose. Enzyme Microb. Technol. 2002, 31, 353–364. [Google Scholar] [CrossRef]
- Zhang, Q.; He, G.; Wang, J.; Cai, W.; Xu, Y. Mechanisms of the stimulatory effects of rhamnolipid biosurfactant on rice straw hydrolysis. Appl. Energy 2009, 86, 233–237. [Google Scholar] [CrossRef]
- Polarz, S.; Kunkel, M.; Donner, A.; Schlötter, M. Added-value surfactants. Chem. Eur. J. 2018, 24, 18842–18856. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Qian, C.; Esker, A.R.; Roman, M. Effect of nonionic surfactants on dispersion and polar interactions in the adsorption of cellulases onto lignin. J. Phys. Chem. B 2017, 121, 9607–9620. [Google Scholar] [CrossRef] [PubMed]
- Henkel, M.; Geissler, M.; Weggenmann, F.; Hausmann, R. Production of microbial biosurfactants: Status quo of rhamnolipid and surfactin towards large-scale production. Biotechnol. J. 2017, 12, 1600561. [Google Scholar] [CrossRef]
- Vatsa, P.; Sanchez, L.; Clement, C.; Baillieul, F.; Dorey, S. Rhamnolipid biosurfactants as new players in animal and plant defense against microbes. Int. J. Mol Sci. 2010, 11, 5095–5108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezerra, K.G.O.; Rufino, R.D.; Luna, J.M.; Sarubbo, L.A. Saponins and microbial biosurfactants: Potential raw materials for the formulation of cosmetics. Biotechnol. Prog. 2018, 34, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Geetha, S.J.; Banat, I.M.; Joshi, S.J. Biosurfactants: Production and potential applications in microbial enhanced oil recovery (MEOR). Biocatal. Agric. Biotechnol. 2018, 14, 23–32. [Google Scholar]
- Parthasarathi, R.; Sivakumaar, P.K. Enhancement effects of biosurfactant produced by Pseudomonas aeruginosa MTCC 2297 and Pseudomonas fluorescens on sugar cane bagasse composting. J. Ecobiotechnol. 2010, 3, 33–39. [Google Scholar]
- Feng, Y.; Jiang, J.; Zhu, L.; Yue, L.; Zhang, J.; Han, S. Effects of tea saponin on glucan conversion and bonding behavior of cellulolytic enzymes during enzymatic hydrolysis of corncob residue with high lignin content. Biotechnol. Biofuels 2013, 6, 161. [Google Scholar] [CrossRef] [Green Version]
- Chong, H.; Li, Q. Microbial production of rhamnolipids: Opportunities, challenges and strategies. Microb. Cell. Fact. 2017, 16, 137. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Y.; Fan, B.Q.; Li, C.H.; Liu, S.; Li, M. Effects of rhamnolipid on the cellulase and xylanase in hydrolysis of wheat straw. Bioresour. Technol. 2011, 102, 6515–6521. [Google Scholar] [CrossRef]
- Bezerra, K.G.O.; Gomes, U.V.R.; Silva, R.O.; Sarubbo, L.A.; Ribeiro, E. The potential application of biosurfactant produced by Pseudomonas aeruginosa TGC01 using crude glycerol on the enzymatic hydrolysis of lignocellulosic material. Biodegradation 2019, 30, 351–361. [Google Scholar] [CrossRef]
- Fiallos-Jurado, J.; Pollier, J.; Moses, T.; Arendt, P.; Barriga-Medina, N.; Morillo, E.; Arahana, V.; Torres, M.L.; Goossens, A.; Leon-Reyes, A. Saponin determination, expression analysis and functional characterization of saponin biosynthetic genes in Chenopodium quinoa leaves. Plant. Sci. 2016, 250, 188–197. [Google Scholar] [CrossRef]
- Xing, Y.; Bu, L.; Sun, D.; Liu, Z.; Liu, S.; Jiang, J. Enhancement of high-solids enzymatic hydrolysis and fermentation of furfural residues by addition of Gleditsia saponin. Fuel 2016, 177, 142–147. [Google Scholar] [CrossRef]
- Xing, Y.; Jia, L.; Liu, Z.; Zhang, W.; Jiang, J. Effects of Gleditsia saponin on high-solids enzymatic hydrolysis of furfural residues. Ind. Crop. Prod. 2015, 64, 209–214. [Google Scholar] [CrossRef]
- Ahumada, A.; Ortega, A.; Chito, D.; Benitez, R. Saponinas de quinua (Chenopodium quinoa Willd.): Un subproducto con alto potencial biológico. Rev. Colomb. Cienc. Quim. Farm. 2016, 45, 438–469. [Google Scholar] [CrossRef]
- Flores, Y.; Díaz, C.; Garay, F.; Colque, O.; Sterner, O.; Almanza, G.R. Oleanane-type triterpenes and derivatives from seed coat of Bolivian Chenopodium quinoa genotype “salar”. Rev. Boliv. Quím. 2005, 22, 71–77. [Google Scholar]
- Kuljanabhagavad, T.; Thongphasuk, P.; Chamulitrat, W.; Wink, M. Triterpene saponins from Chenopodium quinoa Willd. Phytochemistry 2008, 69, 1919–1926. [Google Scholar] [CrossRef]
- Berlowska, J.; Dudkiewicz-Kolodziejska, M.; Pawlikowska, E.; Pielech-Przybylska, K.; Balcerek, M.; Czysowska, A.; Kregiel, D. Utilization of post-fermentation yeasts for yeast extract production by autolysis: The effect of yeast strain and saponin from Quillaja saponaria. J. Inst. Brew. 2017, 123, 396–401. [Google Scholar]
- Park, J.H.; Lee, Y.J.; Kim, Y.H.; Yoon, K.S. Antioxidant and antimicrobial activities of quinoa (Chenopodium quinoa Willd.) seeds cultivated in Korea. Prev. Nutr. Food. Sci. 2017, 22, 195–202. [Google Scholar]
- Fischer, M.J.C.; Pensec, F.; Demangeat, G.; Farine, S.; Chong, J.; Ramirez-Suero, M.; Mazet, F.; Bertsch, C. Impact of Quillaja saponaria saponins on grapevine ecosystem organisms. Anton. Leeuw. 2011, 100, 197–206. [Google Scholar] [CrossRef]
- Gruza, M.M.; Jatczak, K.; Zagrodzki, B.; Łaszcz, M.; Koziak, K.; Malińska, M.; Cmoch, P.; Giller, T.; Zegrocka-Stendel, O.; Woźniak, K.; et al. Preparation, purification and regioselective functionalization of protoescigenin-the main aglycone of escin complex. Molecules 2013, 18, 4389–4402. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, K.B.; Khakimov, B.; Engelsen, S.B.; Bak, S.; Biondi, S.; Jacobsen, S.E. Quinoa seed coats as an expanding and sustainable resource of bioactive compounds: An investigation of genotypic diversity in saponin profiles. Ind. Crops. Prod. 2017, 104, 156–163. [Google Scholar] [CrossRef]
- Wallace-Salinas, V.; Gorwa-Grauslund, M.F. Adaptative evolution of an industrial strain of Saccharomyces cerevisiae for combined tolerance to inhibitors and temperature. Biotechnol. Biofuels 2013, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, K.; Piotrowski, M.; Popielarz, W.; Sosnowski, T.R. Short- and mid-term adsorption behavior of Quillaja bark saponin and its mixtures with lysozyme. Food Hydrocoll. 2011, 25, 687. [Google Scholar] [CrossRef]
- Kiefer, J.; Radzuan, M.N.; Winterburn, J. Infrared spectroscopy for studying structure and aging effects in rhamnolipid biosurfactants. Appl. Sci. 2017, 7, 533. [Google Scholar] [CrossRef]
- Sreij, R.; Dargel, C.; Geisler, P.; Hertle, Y.; Radulescu, A.; Pasini, S.; Pérez, J.; Moleiroa, L.H.; Hellweg, T. DMPC vesicle structure and dynamics in the presence of low amounts of the saponin aescin. Phys. Chem. Chem. Phys. 2018, 20, 9070–9083. [Google Scholar] [CrossRef] [PubMed]
- Escribano, J.; Cabanes, J.; Jiménez-Atiénzar, M.; Ibañez-Tremolada, M.; Gómez-Pando, L.R.; García-Carmona, F.; Gandía-Herrero, F. Characterization of betalains, saponins and antioxidant power in differently colored quinoa (Chenopodium quinoa) varieties. Food Chem. 2017, 234, 285–294. [Google Scholar] [CrossRef]
- Rinderle, S.J.; Goldstein, I.J.; Remsen, E.E. Physicochemical properties of amaranthin, the lectin from Amaranthus caudatus seeds. Biochemistry 1990, 29, 10555–10561. [Google Scholar] [CrossRef]
- Tu, M.B.; Saddler, J.N. Potential enzyme cost reduction with the addition of surfactant during the hydrolysis of pretreated softwood. Appl. Biochem. Biotechnol. 2010, 161, 274–287. [Google Scholar] [CrossRef]
- Kumar, L.; Arantes, V.; Chandra, R.; Saddler, J.N. The lignin present in steam pretreated softwood binds enzymes and limits cellulose accessibility. Bioresour. Technol. 2012, 103, 201–208. [Google Scholar] [CrossRef]
- Nakagame, S.; Chandra, R.P.; Saddler, J.N. The effect of isolated lignins, obtained from a range of pretreated lignocellulosic substrates, on enzymatic hydrolysis. Biotechnol. Bioeng. 2010, 105, 871–879. [Google Scholar] [CrossRef]
- Siqueira, G.; Arantes, V.; Saddler, J.N.; Ferraz, A.; Milagres, A.M.F. Limitation of cellulose accessibility and unproductive binding of cellulases by pretreated sugarcane bagasse lignin. Biotechnol. Biofuels 2017, 10, 176. [Google Scholar] [CrossRef]
- Pekdemir, T.; Opur, M.C.; Urum, K. Emulsification of crude oil–water systems using biosurfactants. Process Saf. Environ. Prot. 2005, 83, 38–46. [Google Scholar] [CrossRef]
- Stanimirova, R.; Marinova, K.; Tcholakova, S.; Denkov, N.D.; Stoyanov, S.; Pelan, E. Surface rheology of saponin adsorption layers. Langmuir 2011, 27, 12486–12498. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Yang, C.; Zhao, Y.; Jia, Y.; Chen, L.W.; Zhou, C.F.; Yong, Q. Promoting enzymatic saccharification of organosolv-pretreated poplar sawdust by saponin-rich tea seed waste. Bioprocess Biosyst. Eng. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cira, L.A.; González, G.A.; Torres, J.C.; Pelayo, C.; Gutiérrez, M.; Ramírez, J. Heterologous expression of Fusarium oxysporum tomatinase in Saccharomyces cerevisiae increases its resistance to saponins and improves ethanol production during the fermentation of Agave tequilana Weber var. azul and Agave salmiana must. A. van Leeuw. J. Microb. 2008, 93, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, J.M.; Contreras, M.F.; Faller, R.; Longo, M.L. Role of unsaturated lipid and ergosterol in ethanol tolerance of model yeast biomembranes. Biophys. J. 2012, 102, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Ghose, T.K. Measurement of cellulose activities (Recommendations of Commission on Biotechnology IUPAC). Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Puls, J.; Schreiber, A.; Saake, B. Conversion of beech wood into platform chemicals after organosolv treatment. In Proceedings of the 15th International Symposium on Wood, Fiber and Pulping Chemistry (ISWFPC 2009), Oslo, Norway, 15–18 June 2009; p. O-036. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proced. 2008, 1617, 1–16. [Google Scholar]
- Koziol, M.J. Afrosimetric estimation of threshold saponin concentration for bitterness in quinoa (Chenopodium quinoa Willd). J. Sci. Food. Agric. 1991, 54, 211–219. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Surfactant | Cellulose Conversion, % (w/w) 1 | ||
|---|---|---|---|---|
| D2: 0 | D2: 2 | D2: 4 | ||
| A | ||||
| Avicel + lignin | None | 76.9 (0.4) | - | - |
| Avicel + lignin | Rhamnolipid | - | 80.2 (3.3) | - |
| Avicel + lignin | Escin | - | 92.4 (3.1) | - |
| B | ||||
| Pretreated solids | None | 70.7 (5.2) | - | - |
| Pretreated solids | Rhamnolipid | - | 75.7 (1.1) | 79.6 (1.2) |
| Pretreated solids | Escin | - | 87.2 (0.4) | 87.4 (0.8) |
| Pretreated solids | RQ saponin3 | - | 75.3 (2.0) | 81.9 (0.5) |
| Pretreated solids | WQ saponin4 | - | 73.0 (0.2) | 76.8 (0.7) |
| Experimental Reaction | RQS Dosage, g/100 g | Biomass Loading, % (w/w) | Enzyme Loading, FPU and CBU/g dry Weight Substrate |
|---|---|---|---|
| 1 | 4 | 5 | 5 |
| 2 | 2 | 5 | 7.5 |
| 3 | 6 | 5 | 7.5 |
| 4 | 4 | 5 | 10 |
| 5 | 2 | 10 | 5 |
| 6 | 6 | 10 | 5 |
| 7 | 4 | 10 | 7.5 |
| 8 | 2 | 10 | 10 |
| 9 | 6 | 10 | 10 |
| 10 | 4 | 15 | 5 |
| 11 | 2 | 15 | 7.5 |
| 12 | 6 | 15 | 7.5 |
| 13 | 4 | 15 | 10 |
| Control | 0 | 10 | 7.5 |
| Enzyme Loading, FPU/g | Saponins Dosage, g/100 g | Protein Concentration, mg/mL | Free Protein, % | |
|---|---|---|---|---|
| Initial | Final | |||
| 5 | 2 | 1.24 (0.01) | 0.48 (0.01) | 38.7 |
| 5 | 6 | 1.24 (0.01) | 0.58 (0.01) | 47.7 |
| 7.5 | 0 | 1.39 (0.10) | 0.48 (0.01) | 34.6 |
| 7.5 | 4 | 1.39 (0.10) | 0.53 (0.01) | 38.1 |
| 10 | 2 | 1.69 (0.10) | 0.56 (0.01) | 33.3 |
| 10 | 6 | 1.69 (0.10) | 0.76(0.04) | 43.3 |
| RQSinitial, g/L | RQSfinal, g/L | RQS Recovery, % |
|---|---|---|
| 2.0 | 1.0 (±0.1) | 50.0 |
| 6.0 | 5.0 (±0.0) | 83.3 |
| 8.0 | 7.5 (±0.1) | 93.8 |
| 10.0 | 9.5 (±0.1) | 95.0 |
| 12.0 | 11.6 (±0.1) | 96.6 |
| Source of Variation | Sum of Squares | Degree of Freedom | Mean Squares | F-Ratio | P-Value | F Critical Value |
|---|---|---|---|---|---|---|
| Between groups | 1136.787 | 3 | 378.929 | 587.721 | 1.16 × 10−19 | 3.090 |
| Within groups | 12.895 | 20 | 0.645 | |||
| Total | 1149.681 | 23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliva-Taravilla, A.; Carrasco, C.; Jönsson, L.J.; Martín, C. Effects of Biosurfactants on Enzymatic Saccharification and Fermentation of Pretreated Softwood. Molecules 2020, 25, 3559. https://doi.org/10.3390/molecules25163559
Oliva-Taravilla A, Carrasco C, Jönsson LJ, Martín C. Effects of Biosurfactants on Enzymatic Saccharification and Fermentation of Pretreated Softwood. Molecules. 2020; 25(16):3559. https://doi.org/10.3390/molecules25163559
Chicago/Turabian StyleOliva-Taravilla, Alfredo, Cristhian Carrasco, Leif J. Jönsson, and Carlos Martín. 2020. "Effects of Biosurfactants on Enzymatic Saccharification and Fermentation of Pretreated Softwood" Molecules 25, no. 16: 3559. https://doi.org/10.3390/molecules25163559
APA StyleOliva-Taravilla, A., Carrasco, C., Jönsson, L. J., & Martín, C. (2020). Effects of Biosurfactants on Enzymatic Saccharification and Fermentation of Pretreated Softwood. Molecules, 25(16), 3559. https://doi.org/10.3390/molecules25163559