
Comparison of the Partial Structure and Antioxidant Activity of Polysaccharides from Two Species of Chinese Truffles


Abstract
:1. Introduction
2. Results and Discussion
2.1. The Carbohydrate, Phenol, and Protein Content of Tubers Extracts
2.2. Fractionation of Polysaccharides from Tubers
2.3. Molecular Weight of Polysaccharides
2.4. Chemical Composition of Polysaccharide Fractions
2.5. Glycosidic Linkages
2.6. Antioxidant Activity
2.6.1. Determination of Cell Viability
2.6.2. Antioxidant Enzyme Activity
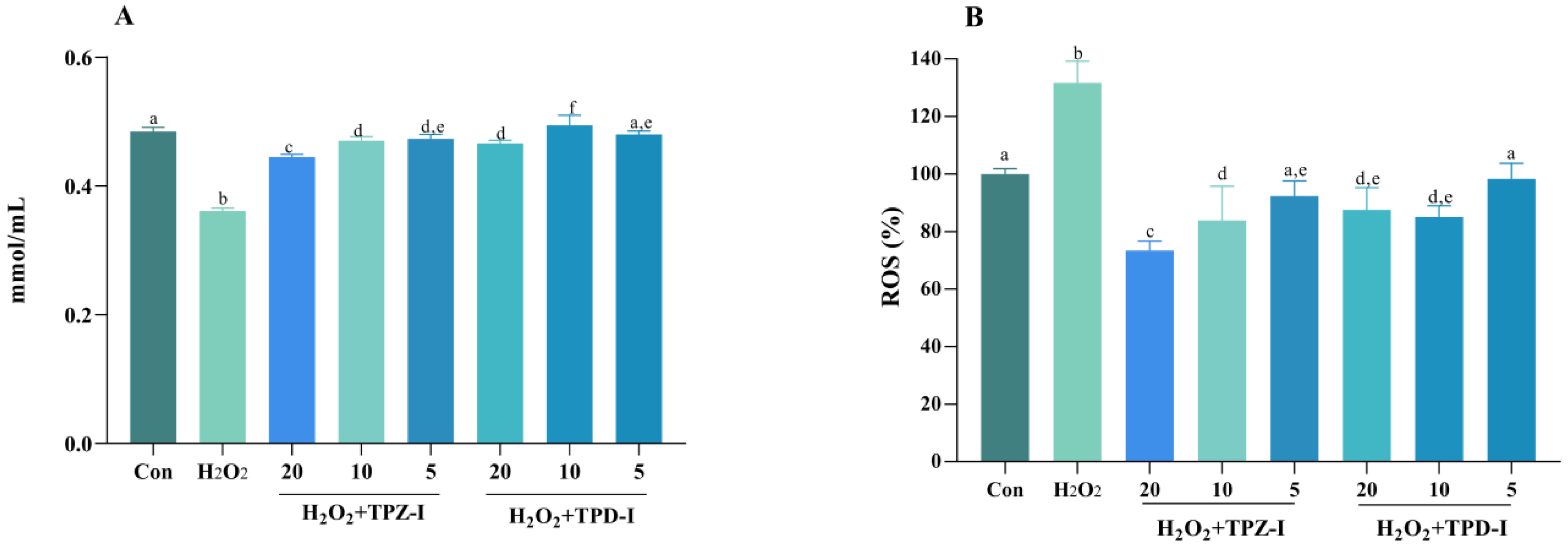
2.6.3. Effects of Acidic Polysaccharides on the Activities of T-AOC and ROS
3. Materials and Methods
3.1. Extraction of Polysaccharides
3.2. Purification of Polysaccharides
3.2.1. Ion-Exchange Chromatography on Diethylaminoethyl-Cellulose
3.2.2. Acidic Polysaccharide Purification
3.3. Determination of Molecular Weight
3.4. Chemical Compositions and Glycosidic Linkage Determination
3.5. Antioxidant Activity Assay
3.5.1. Cell Culture
3.5.2. Establishment of an Oxidative Stress Damage Model
3.5.3. Measurement of IPEC-J2 Cells Viability
3.5.4. Determination of Antioxidant Enzymes Activity
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Berch, S.M.; Bonito, G. Truffle diversity (Tuber, Tuberaceae) in British Columbia. Mycorrhiza 2016, 26, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Mello, A.; Murat, C.; Bonfante, P. Truffles: Much more than a prized and local fungal delicacy. FEMS Microbiol. Lett. 2006, 260, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Linde, C.C.; Selmes, H. Genetic diversity and mating type distribution of Tuber melanosporum and their significance to truffle cultivation in artificially planted truffieres in Australia. Appl. Environ. Microb. 2012, 78, 6534–6539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, M.; Annegret, K.; Claude, M.; Raffaella, B.; Pedro, M.C.; Olivier, J.; Barbara, M.; Emmanuelle, M.; Benjamin, N.; Riccardo, P.; et al. Peerigord black truffle genome uncovers evolutionary origins and mechanisms of symbiosis. Nature 2010, 464, 1033–1038. [Google Scholar]
- Gao, J.; Hu, L.; Liu, J. A novel sterol from Chinese truffles Tuber indicum. Steroids 2001, 66, 771–775. [Google Scholar]
- Luo, Q.; Zhang, J.; Yan, L.; Tang, Y.; Ding, X.; Yang, Z.; Sun, Q. Composition and antioxidant activity of water-soluble polysaccharides from Tuber Indicum. J. Med. Food 2011, 14, 1609–1616. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tan, Z.M.; Murat, C.; Jeandroz, S.; Le Tacon, F. Molecular taxonomy of Chinese truffles belonging to the Tuber rufum and Tuber puberulum groups. Fungal. Divers. 2007, 24, 301–328. [Google Scholar]
- Liu, R.S.; Tang, Y.J. Tuber melanosporum fermentation medium optimization by Plackett-Burman design coupled with Draper-Lin small composite design and desirability function. Bioresource Technol. 2010, 101, 3139–3146. [Google Scholar] [CrossRef]
- Francesco, P.; Andrea, R.; Bruno, G.; Sergio, A. Typing Tuber melanosporum and Chinese black truffle species by molecular markers. FEMS Microbiol. Lett. 1997, 153, 255–260. [Google Scholar]
- Wang, Y.; Moreno, G.; Riousset, L.J.; Manjón, J.L.; Diéz, J. Tuber pseudoexcavatum sp. nov. A new species from China commercialised in Spain, France and Italy with additional comments on Chinese truffles. Cryptogamie Mycol. 1998, 19, 113–120. [Google Scholar]
- Chen, J.; Liu, P.G. Delimitation of Tuber pseudohimalayense and T. pseudoexcavatum Based on Morphological and Molecular Data. Cryptogamie Mycol. 2011, 32, 83–93. [Google Scholar] [CrossRef]
- Patel, S.; Rauf, A.; Khan, H.; Khalid, S.; Patel, S.; Mubarak, M.S. Potential health benefits of natural products derived from truffles: A review. Trends Food Sci. Tech. 2017, 70, 1–8. [Google Scholar] [CrossRef]
- Bhotmange, D.U.; Wallenius, J.H.; Singhal, R.S.; Shamekh, S.S. Enzymatic extraction and characterization of polysaccharide from Tuber aestivum. Bioact. Carbohyd Dietary Fibre 2017, 10, 1–9. [Google Scholar] [CrossRef]
- Luo, A.; He, X.; Zhou, S.; Fan, Y.; Luo, A.; Chun, Z. Purification, composition analysis and antioxidant activity of the polysaccharides from Dendrobium nobile Lindl. Carbohyd. Polym. 2010, 79, 1014–1019. [Google Scholar] [CrossRef]
- Wang, S.; Marcone, M.F. The biochemistry and biological properties of the world’s most expensive underground edible mushroom: Truffles. Food Res. Int. 2011, 44, 2567–2581. [Google Scholar] [CrossRef]
- Pattanayak, M.; Samanta, S.; Maity, P.; Manna, D.K.; Sen, I.K.; Nandi, A.K.; Panda, B.C.; Chattopadhyay, S.; Roy, S.; Sahoo, A.K.; et al. Polysaccharide of an edible truffle Tuber rufum: Structural studies and effects on human lymphocytes. Int. J. Biol. Macromol. 2017, 95, 1037–1048. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, X.H.; Li, H.M.; Wang, S.H.; Chen, T.; Yuan, Z.P.; Tang, Y.J. Isolation and characterization of polysaccharides with the antitumor activity from Tuber fruiting bodies and fermentation system. Appl. Microbiol. Biot. 2014, 98, 1991–2002. [Google Scholar] [CrossRef]
- Dubost, N.J.; Ou, B.; Beelman, R.B. Quantification of polyphenols and ergothioneine in cultivated mushrooms and correlation to total antioxidant capacity. Food Chem. 2007, 105, 727–735. [Google Scholar] [CrossRef]
- Surai, P.; Fisinin, V. Antioxidant-prooxidant balance in the intestine: Applications in chick placement and pig weaning. J. Vet. Sci. Med. 2015, 3, 1–16. [Google Scholar]
- Doan, H.H.; Aydin, S. Determination of antimicrobial effect, antioxidant activity and phenolic contents of desert truffle in Turkey. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Murcia, M.A.; MartÍNez-TomÉ, M.; JimÉNez, A.M.; Vera, A.M.; Honrubia, M.; Parras, P. Antioxidant Activity of Edible Fungi (Truffles and Mushrooms): Losses during Industrial Processing. J. Food Prot. 2002, 65, 1614–1622. [Google Scholar] [CrossRef] [PubMed]
- Jin-Mei, Y.; Biao, P.U.; An-Jun, C.; Xing-Yan, L. Antioxidant Activity in vitro of Polysaccharides with Different Molecular Weights from Truffles. Food Chem. 2013, 34, 127–131. [Google Scholar]
- Salyers, A.A.; Palmer, J.K.; Wilkins, T.D. Degradation of polysaccharides by intestinal bacterial enzymes. Am. J. Clin. Nutr. 1978, 31, S128–S130. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chai, D.-D.; Li, H.-M.; Chen, T.; Tang, Y.-J. Significance of metal ion supplementation in the fermentation medium on the structure and anti-tumor activity of Tuber polysaccharides produced by submerged culture of Tuber melanosporum. Process. Biochem. 2014, 49, 2030–2038. [Google Scholar] [CrossRef]
- Dolfi, S.; Sveronis, A.; Silipo, A.; Rizzo, R.; Cescutti, P. A novel rhamno-mannan exopolysaccharide isolated from biofilms of Burkholderia multivorans C1576. Carbohyd. Res. 2015, 411, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, B.; Li, J.; Zhou, J.; He, X.; Ye, L.; Zou, J.; Wu, C.; Zhang, X.; Peng, W. Purification, characterization, and complement fixation activity of acidic polysaccharides from Tuber sinoaestivum. LWT-Food Sci. Technol. 2017, 85, 82–88. [Google Scholar] [CrossRef]
- Lo, T.C.-T.; Chang, C.A.; Chiu, K.-H.; Tsay, P.-K.; Jen, J.-F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohyd. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Zhang, C.-H.; Yu, Y.; Liang, Y.-Z.; Chen, X.-Q. Purification, partial characterization and antioxidant activity of polysaccharides from Glycyrrhiza uralensis. Int. J. Biol. Macromol. 2015, 79, 681–686. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, S.; Ran, C.; Wang, L.; Kan, J. Extraction, characterization and antioxidant activity of water-soluble polysaccharides from Tuber huidongense. Int. J. Biol. Macromol. 2016, 91, 431–442. [Google Scholar] [CrossRef]
- Liu, Y.; Qiang, M.; Sun, Z.; Du, Y. Optimization of ultrasonic extraction of polysaccharides from Hovenia dulcis peduncles and their antioxidant potential. Int. J. Biol. Macromol. 2015, 80, 350–357. [Google Scholar] [CrossRef]
- Qi, H.; Zhao, T.; Zhang, Q.; Li, Z.; Zhao, Z.; Xing, R. Antioxidant activity of different molecular weight sulfated polysaccharides from Ulva pertusa Kjellm (Chlorophyta). J. Appl. Phycol. 2005, 17, 527–534. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, Q.; Qi, H.; Zhang, H.; Niu, X.; Xu, Z.; Li, Z. Degradation of porphyran from Porphyra haitanensis and the antioxidant activities of the degraded porphyrans with different molecular weight. Int. J. Biol. Macromol. 2006, 38, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, X.; Mo, X.; Qi, H. Degradation and the antioxidant activity of polysaccharide from Enteromorpha linza. Carbohyd. Polym. 2013, 92, 2084–2087. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Huang, C.; Cao, X.; Chen, X.; Fu, Y.; Zhu, Y.; Chen, Z.; Luo, Q.; Li, L.; Song, X.; Jia, R.; et al. A pectic polysaccharide from Ligusticum chuanxiong promotes intestine antioxidant defense in aged mice. Carbohyd. Polym. 2017, 174, 915–922. [Google Scholar] [CrossRef]
- Zhu, Z.Y.; Liu, R.Q.; Si, C.L.; Zhou, F.; Wang, Y.X.; Ding, L.N.; Jing, C.; Liu, A.J.; Zhang, Y.M. Structural analysis and anti-tumor activity comparison of polysaccharides from Astragalus. Carbohyd. Polym. 2011, 85, 895–902. [Google Scholar] [CrossRef]
- Austarheim, I.; Christensen, B.E.; Hegna, I.K.; Petersen, B.O.; Duus, J.O.; Bye, R.; Michaelsen, T.E.; Diallo, D.; Inngjerdingen, M.; Paulsen, B.S. Chemical and biological characterization of pectin-like polysaccharides from the bark of the Malian medicinal tree Cola cordifolia. Carbohyd. Polym. 2012, 89, 259–268. [Google Scholar] [CrossRef]
- Kim, J.-B.; Carpita, N.C. Changes in Esterification of the Uronic Acid Groups of Cell Wall Polysaccharides during Elongation of Maize Coleoptiles. Plant. Physiol. 1992, 98, 646. [Google Scholar] [CrossRef] [Green Version]
- Sweet, D.P.; Shapiro, R.H.; Albersheim, P. Quantitative analysis by various g.l.c. response-factor theories for partially methylated and partially ethylated alditol acetates. Carbohyd. Res. 1975, 40, 217–225. [Google Scholar] [CrossRef]
- Zou, Y.-F.; Zhang, Y.-Y.; Paulsen, B.S.; Rise, F.; Chen, Z.-L.; Jia, R.-Y.; Li, L.-X.; Song, X.; Feng, B.; Tang, H.-Q.; et al. Structural features of pectic polysaccharides from stems of two species of Radix Codonopsis and their antioxidant activities. Int. J. Biol. Macromol. 2020, 159, 704–713. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds TPZ-NP, TPZ-I, TPZ-II, TPD-NP, and TPD-I are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPD | TPZ | |
|---|---|---|
| Protein | 3.79% | 4.15% |
| Phenol | 10.11% | 15.48% |
| Carbohydrates | 57.90% | 67.40% |
| Fraction | Molecular Weight (kDa) |
|---|---|
| TPZ-NP | 5.13 |
| TPZ-I | 30.84 |
| TPZ-II | 17.43 |
| TPD-NP | 8.37 |
| TPD-I | 18.91 |
| TPZ-NP | TPZ-I | TPZ-II | TPD-NP | TPD-I | |
|---|---|---|---|---|---|
| Rha a | 23.9 | 23.0 | 22.2 | 19.8 | 19.7 |
| Man b | 62.3 | 67.8 | 58.8 | 63.4 | 56.0 |
| Gal c | 4.7 | 3.6 | 5.6 | 2.4 | 4.7 |
| Glc d | 9.2 | 4.8 | 9.4 | 14.4 | 17.7 |
| ManA e | n.d. f | 0.9 | 4.0 | n.d. f | 1.9 |
| TPZ-I | TPD-I | ||
|---|---|---|---|
| Rha | T | 12.6 | 9.7 |
| 1→2 | 5.4 | 5.6 | |
| 1→3 | 5.0 | 4.4 | |
| Man | T | 5.2 | 8.0 |
| 1→2 | 23.3 | 15.6 | |
| 1→3 | 18.7 | 12.4 | |
| 1→6 | 2.3 | 1.8 | |
| 1→2, 3 | 3.0 | 3.6 | |
| 1→3, 6 | 1.9 | n.d. | |
| 1→2, 6 | 13.4 | 14.6 | |
| ManA | 1→4 | 0.9 | 1.9 |
| Gal | T | 1.3 | 1.4 |
| 1→3 | 2.3 | 3.3 | |
| Glc | 1→4 | 4.8 | 17.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhu, Z.; Ye, L.; Kang, Z.; Zhang, X.; Huang, Y.; Zhang, B.; Zou, Y. Comparison of the Partial Structure and Antioxidant Activity of Polysaccharides from Two Species of Chinese Truffles. Molecules 2020, 25, 4345. https://doi.org/10.3390/molecules25184345
Li X, Zhu Z, Ye L, Kang Z, Zhang X, Huang Y, Zhang B, Zou Y. Comparison of the Partial Structure and Antioxidant Activity of Polysaccharides from Two Species of Chinese Truffles. Molecules. 2020; 25(18):4345. https://doi.org/10.3390/molecules25184345
Chicago/Turabian StyleLi, Xiaolin, Zhongkai Zhu, Lei Ye, Zongjing Kang, Xiaoping Zhang, Yue Huang, Bo Zhang, and Yuanfeng Zou. 2020. "Comparison of the Partial Structure and Antioxidant Activity of Polysaccharides from Two Species of Chinese Truffles" Molecules 25, no. 18: 4345. https://doi.org/10.3390/molecules25184345
APA StyleLi, X., Zhu, Z., Ye, L., Kang, Z., Zhang, X., Huang, Y., Zhang, B., & Zou, Y. (2020). Comparison of the Partial Structure and Antioxidant Activity of Polysaccharides from Two Species of Chinese Truffles. Molecules, 25(18), 4345. https://doi.org/10.3390/molecules25184345