
Neoglycosylated Collagen: Effect on Neuroblastoma F-11 Cell Lines


 , and
, and 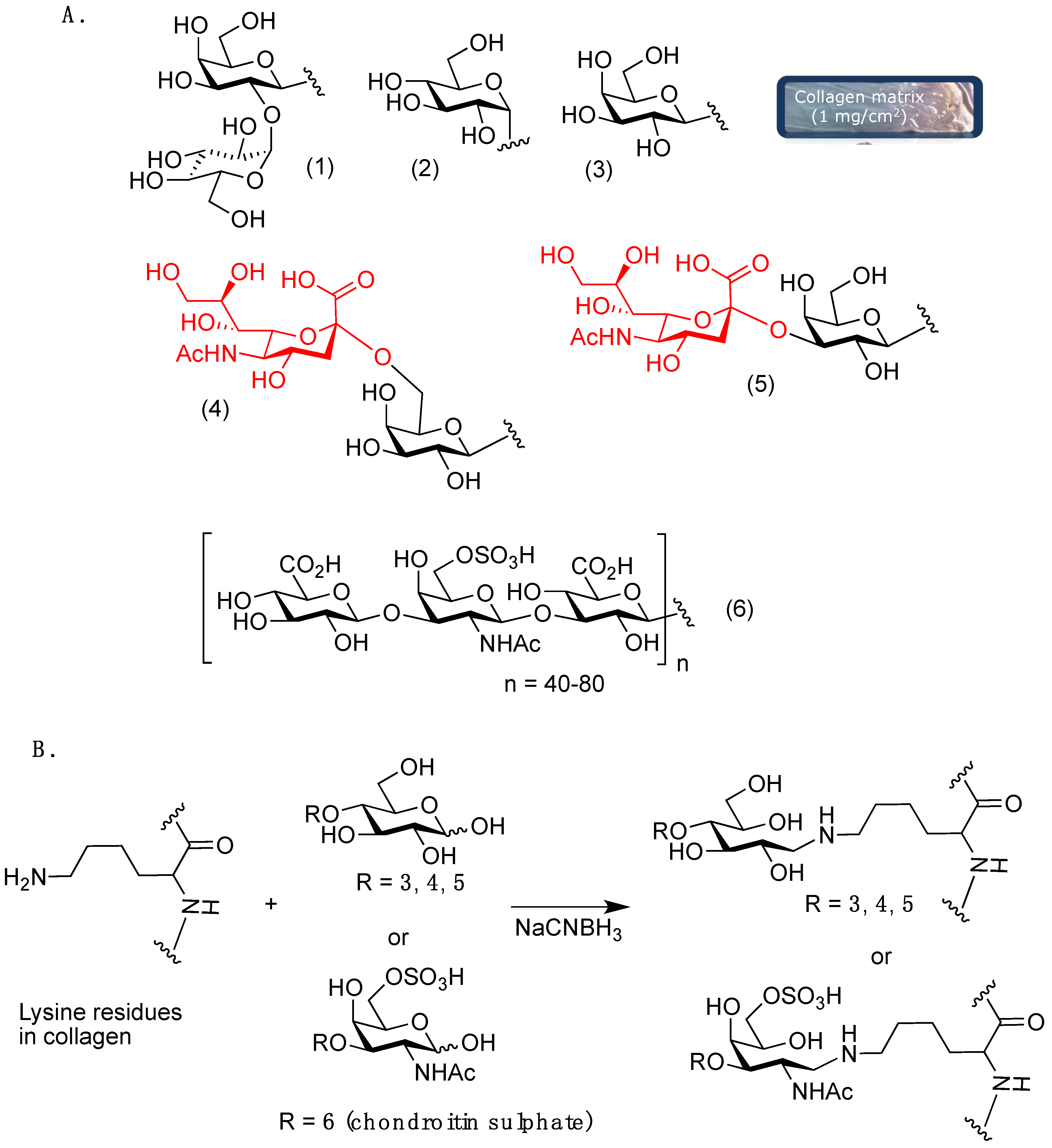{kind=link}
{kind=link}
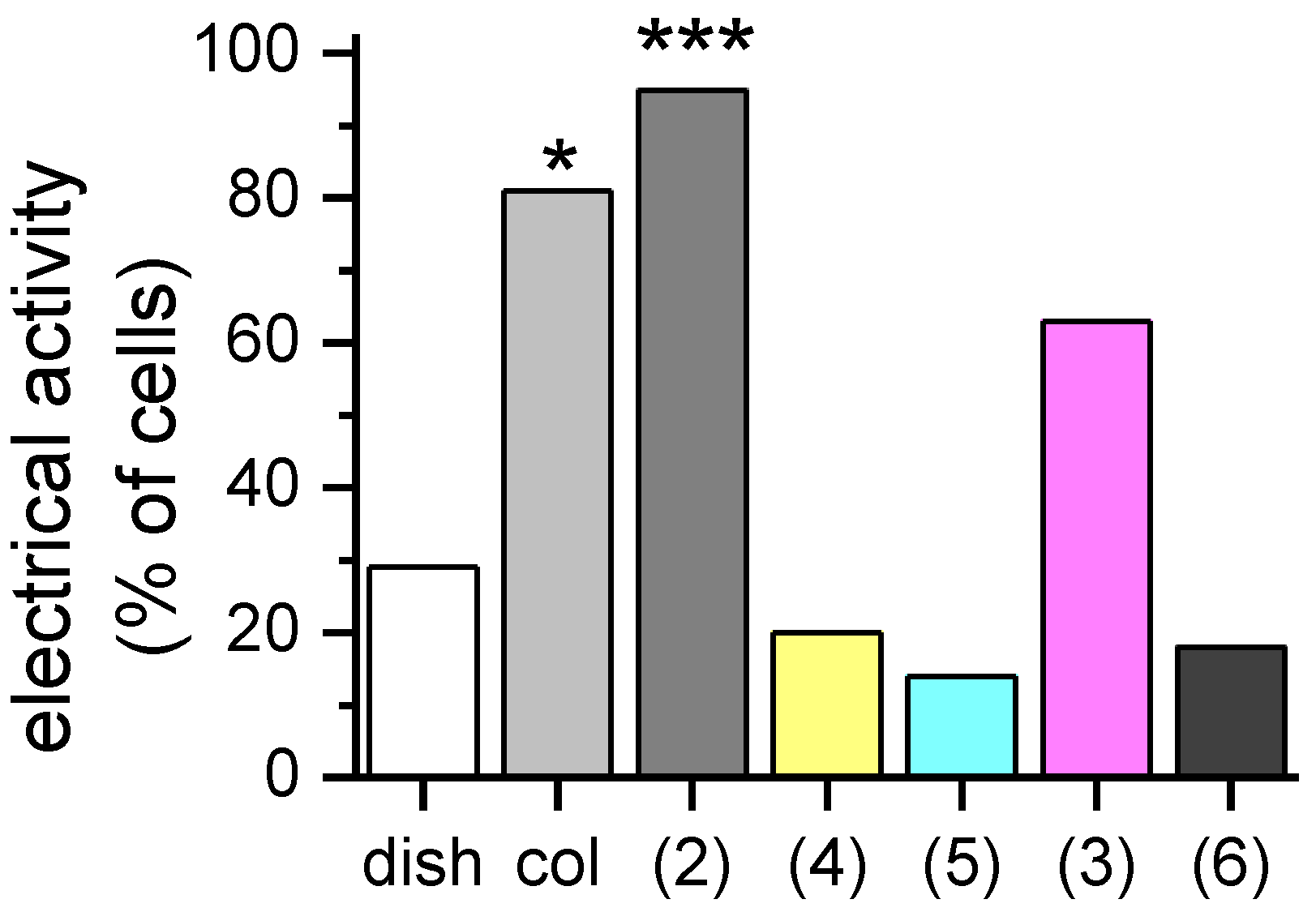{kind=link}
Abstract
:1. Introduction
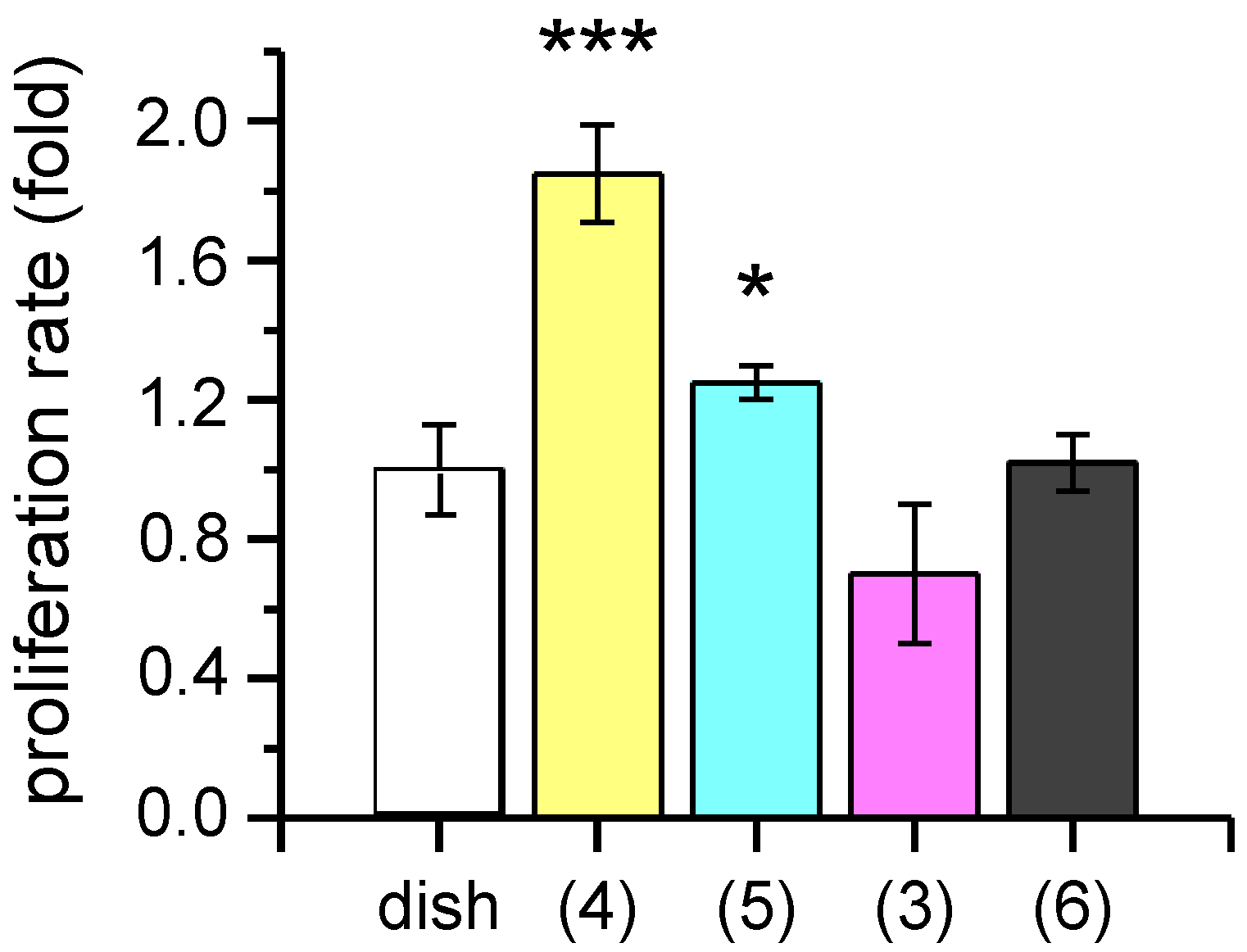
2. Results and Discussion
3. Materials and Methods
3.1. General Methods
3.2. Matrices Preparation
3.2.1. Pristine Collagen Matrices
3.2.2. Carbohydrate-Grafted Collagen Matrices
3.3. Cell Culture
3.3.1. Proliferation Assay and Functional Analysis by Patch-Clamp Recordings
3.3.2. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Naqvi, S.; Panghal, A.; Flora, S.J.S. Nanotechnology: A Promising Approach for Delivery of Neuroprotective Drugs. Front. Neurosci. 2020, 14, 494. [Google Scholar] [CrossRef] [PubMed]
- Hussain, R.; Zubair, H.; Pursell, S.; Shahab, M. Neurodegenerative Diseases: Regenerative Mechanisms and Novel Therapeutic Approaches. Brain Sci. 2018, 8, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, R.Y.; Fuehrmann, T.; Mitrousis, N.; Shoichet, M.S. Regenerative Therapies for Central Nervous System Diseases: A Biomaterials Approach. Neuropsychopharmacology 2014, 39, 169–188. [Google Scholar] [CrossRef] [Green Version]
- Orive, G.; Anitua, E.; Pedraz, J.L.; Emerich, D.F. Biomaterials for promoting brain protection, repair and regeneration. Nat. Rev. Neurosci. 2009, 10, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Friess, W. Collagen—Biomaterial for drug delivery. Eur. J. Pharm. Biopharm. 1998, 45, 113–136. [Google Scholar] [CrossRef]
- Abou Neel, E.A.; Bozec, L.; Knowles, J.C.; Syed, O.; Mudera, V.; Day, R.; Hyun, J.K. Collagen—Emerging collagen based therapies hit the patient. Adv. Drug Deliv. Rev. 2013, 65, 429–456. [Google Scholar] [CrossRef]
- Ucar, B.; Humpel, C. Collagen for brain repair: Therapeutic perspectives. Neural Regen. Res. 2018, 13, 595–598. [Google Scholar]
- Kourgiantaki, A.; Tzeranis, D.S.; Karali, K.; Georgelou, K.; Bampoula, E.; Psilodimitrakopoulos, S.; Yannas, I.V.; Stratakis, E.; Sidiropoulou, K.; Charalampopoulos, I.; et al. Neural stem cell delivery via porous collagen scaffolds promotes neuronal differentiation and locomotion recovery in spinal cord injury. NPJ Regen. Med. 2020, 5, 12. [Google Scholar] [CrossRef]
- Khaing, Z.Z.; Schmidt, C.E. Advances in natural biomaterials for nerve tissue repair. Neurosci. Lett. 2012, 519, 103–114. [Google Scholar] [CrossRef]
- Khaing, Z.Z.; Thomas, R.C.; Geissler, S.A.; Schmidt, C.E. Advanced biomaterials for repairing the nervous system: What can hydrogels do for the brain? Mater. Today 2014, 17, 332–340. [Google Scholar] [CrossRef]
- Kucharz, E.J. Collagen in the Nervous System. In The Collagens: Biochemistry and Pathophysiology; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Gregorio, I.; Braghetta, P.; Bonaldo, P.; Cescon, M. Disease Models & Mechanisms Collagen VI in healthy and diseased nervous system. Dis. Models Mech. 2018, 11, dmm032946. [Google Scholar]
- Russo, L.; Cipolla, L. Glycomics: New challenges and opportunities in regenerative medicine. Chem. Eur. J. 2016, 22, 13380–13388. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; Sgambato, A.; Guizzardi, R.; Vesentini, S.; Cipolla, L.; Nicotra, F. Glyco-Functionalysed Biomaterials in Neuroregeneration. In Drug and Gene Delivery to the Central Nervous System for Neuroprotection; Sharma, H., Muresanu, D., Sharma, A., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 179–198. [Google Scholar]
- Rebelo, A.L.J.; Bizeau, J.; Russo, L.; Pandit, A. Glycan-Functionalized Collagen Hydrogels Modulate the Glycoenvironment of a Neuronal Primary Culture. Biomacromolecules 2020, 21, 2681–2694. [Google Scholar] [CrossRef]
- Russo, L.; Sgambato, A.; Lecchi, M.; Pastori, V.; Raspanti, M.; Natalello, A.; Doglia, S.M.; Nicotra, F.; Cipolla, L. Neoglucosylated Collagen Matrices Drive Neuronal Cells to Differentiate. ACS Chem. Neurosci. 2014, 5, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennet, T. Collagen glycosylation. Curr. Opin. Struct. Biol. 2019, 56, 131–138. [Google Scholar] [CrossRef]
- Sgambato, A.; Russo, L.; Montesi, M.; Panseri, S.; Marcacci, M.; Caravà, E.; Raspanti, M.; Cipolla, L. Different Sialoside Epitopes on Collagen Film Surfaces Direct Mesenchymal Stem Cell Fate. ACS Appl. Mater. Interfaces 2016, 8, 14952–14957. [Google Scholar] [CrossRef]
- Russo, L.; Battocchio, C.; Secchi, V.; Magnano, E.; Nappini, S.; Taraballi, F.; Gabrielli, L.; Comelli, F.; Papagni, A.; Costa, B.; et al. Thiol-ene mediated neoglycosylation of collagen patches: A preliminary study. Langmuir 2014, 30, 1336–1342. [Google Scholar] [CrossRef]
- Russo, L.; Sgambato, A.; Giannoni, P.; Quarto, R.; Vesentini, S.; Gautieri, A.; Cipolla, L. Response of osteoblast-like MG63 on neoglycosylated collagen matrices. MedChemCommun 2014, 5, 1208–1212. [Google Scholar] [CrossRef] [Green Version]
- Boggs, J.M. Role of Galactosylceramide and Sulfatide in Oligodendrocytes and CNS Myelin: Formation of a Glycosynapse. In Glycobiology of the Nervous System. Advances in Neurobiology; Yu, R., Schengrund, C.L., Eds.; Springer: New York, NY, USA, 2014; Volume 9. [Google Scholar]
- Kalovidouris, S.A.; Gama, C.I.; Lee, L.W.; Hsieh-Wilson, L.C. A role for fucose α(1-2) galactose carbohydrates in neuronal growth. J. Am. Chem. Soc. 2005, 127, 1340–1341. [Google Scholar] [CrossRef] [Green Version]
- Bornhöfft, K.F.; Goldammer, T.; Rebl, A.; Galuska, S.P. Siglecs: A journey through the evolution of sialic acid-binding immunoglobulin-type lectins. Dev. Comp. Immunol. 2018, 86, 219–231. [Google Scholar] [CrossRef]
- Snow, D.M.; Brown, E.M.; Letourneau, P.C. Growth cone behavior in the presence of soluble chondroitin sulfate proteoglycan (cspg), compared to behavior on cspg bound to laminin or fibronectin. Int. J. Dev. Neurosci. 1996, 14, 331–349. [Google Scholar] [CrossRef]
- Kwon, H.J.; Han, Y. Chondroitin sulfate-based biomaterials for tissue engineering Turk. J. Biol. 2016, 40, 290–299. [Google Scholar]
- Tomac, A.; Lindqvist, E.; Lin, L.F.; Ogren, S.O.; Young, D.; Hoffer, B.J.; Olson, L. Protection and repair of the nigrostriatal dopaminergic system by GDNF In Vivo. Nature 1995, 373, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Raspanti, M.; Caravà, E.; Sgambato, A.; Natalello, A.; Russo, L.; Cipolla, L. The collaggrecan: Synthesis and visualization of an artificial proteoglycan. Int. J. Biol. Macromol. 2016, 86, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Pastori, V.; D’Aloia, A.; Blasa, S.; Lecchi, M. Serum-deprived differentiated neuroblastoma F-11 cells express functional dorsal root ganglion neuron properties. PeerJ 2019, 7, e7951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schengrund, C.-L. Gangliosides and neuroblastomas. Int. J. Mol. Sci. 2020, 21, 5313. [Google Scholar] [CrossRef]
- Taraballi, F.; Zanini, S.; Lupo, C.; Panseri, S.; Cunha, C.; Riccardi, C.; Marcacci, M.; Campione, M.; Cipolla, L. Amino and carboxyl plasma functionalization of collagen films for tissue engineering applications. J. Colloid Interface Sci. 2013, 394, 590–597. [Google Scholar] [CrossRef]
- Russo, L.; Gautieri, A.; Raspanti, M.; Taraballi, F.; Nicotra, F.; Vesentini, S.; Cipolla, L. Carbohydrate-functionalized collagen matrices: Design and characterization of a novel neoglycosylated biomaterial. Carbohydr. Res. 2014, 389, 12–17. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sgambato, A.; Pastori, V.; Russo, L.; Vesentini, S.; Lecchi, M.; Cipolla, L. Neoglycosylated Collagen: Effect on Neuroblastoma F-11 Cell Lines. Molecules 2020, 25, 4361. https://doi.org/10.3390/molecules25194361
Sgambato A, Pastori V, Russo L, Vesentini S, Lecchi M, Cipolla L. Neoglycosylated Collagen: Effect on Neuroblastoma F-11 Cell Lines. Molecules. 2020; 25(19):4361. https://doi.org/10.3390/molecules25194361
Chicago/Turabian StyleSgambato, Antonella, Valentina Pastori, Laura Russo, Simone Vesentini, Marzia Lecchi, and Laura Cipolla. 2020. "Neoglycosylated Collagen: Effect on Neuroblastoma F-11 Cell Lines" Molecules 25, no. 19: 4361. https://doi.org/10.3390/molecules25194361
APA StyleSgambato, A., Pastori, V., Russo, L., Vesentini, S., Lecchi, M., & Cipolla, L. (2020). Neoglycosylated Collagen: Effect on Neuroblastoma F-11 Cell Lines. Molecules, 25(19), 4361. https://doi.org/10.3390/molecules25194361