Exploring the Proteomic Alterations from Untreated and Cryoablation and Irradiation Treated Giant Cell Tumors of Bone Using Liquid-Chromatography Tandem Mass Spectrometry

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
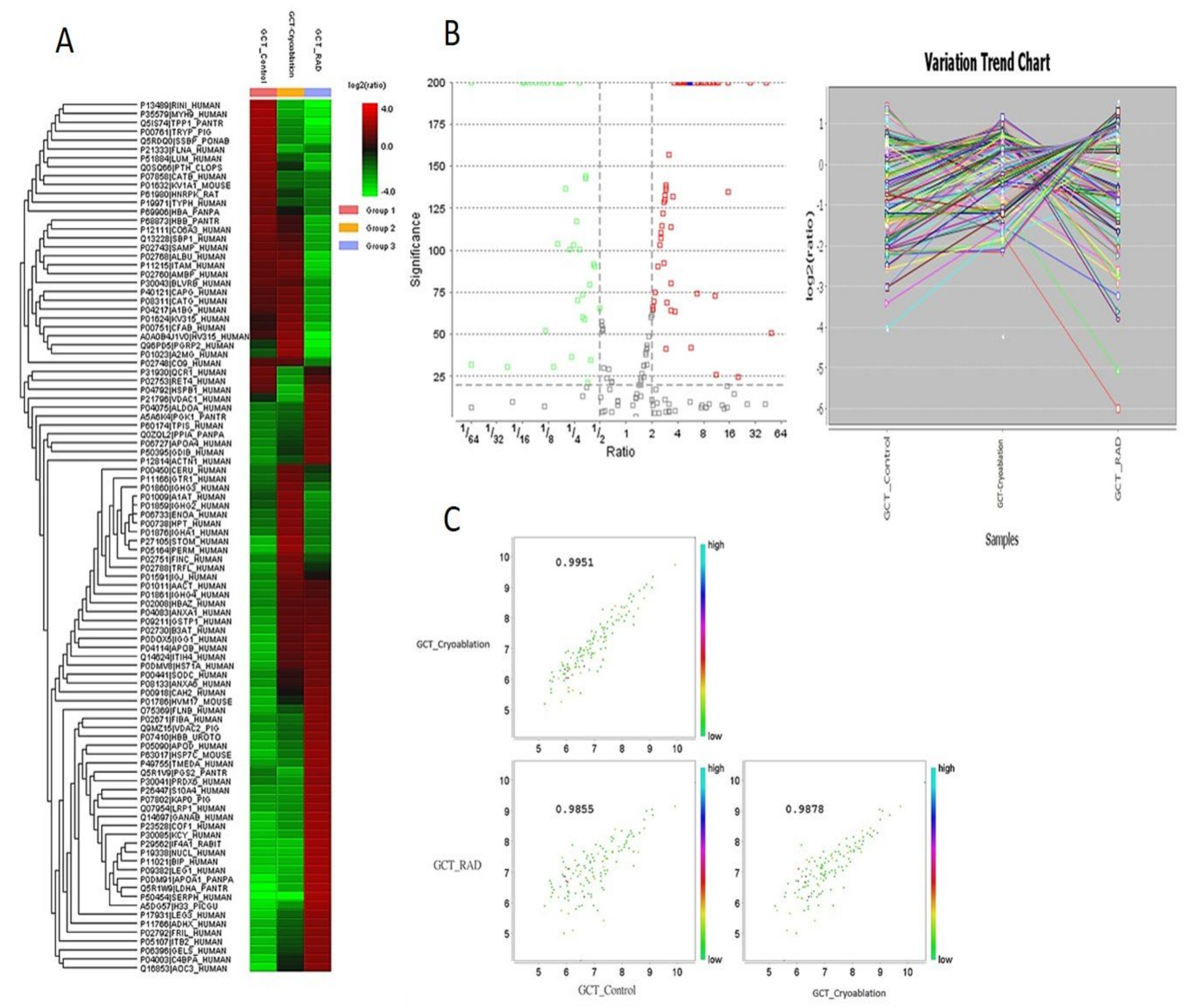
2.1. Protein Expression Profiles from GCT-Untreated/Control, and Cryoablation and Irradiation Treated Groups
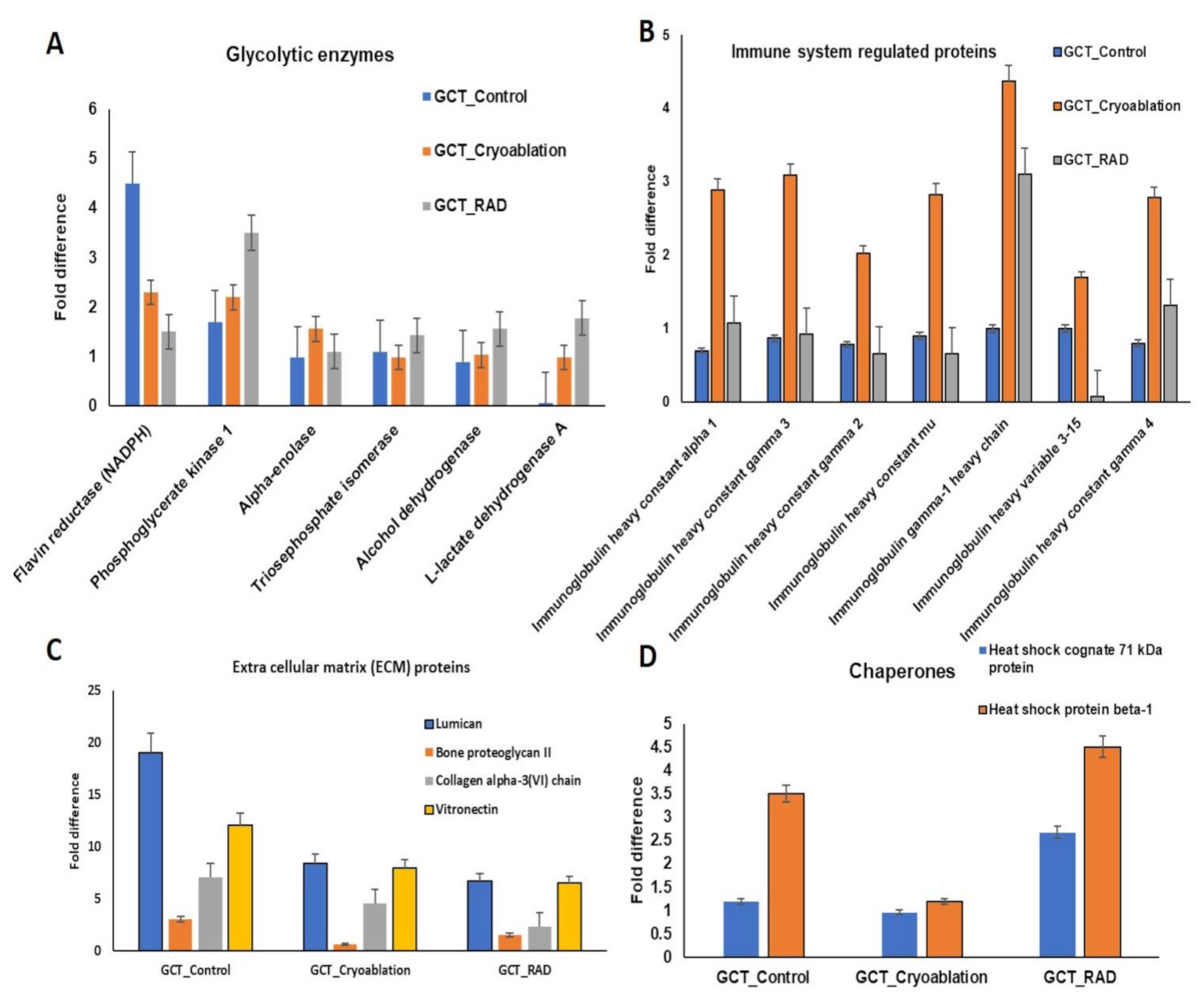
2.2. Exploring the Altered Protein Expressions in GCT-Treated and Untreated/Control Tumors
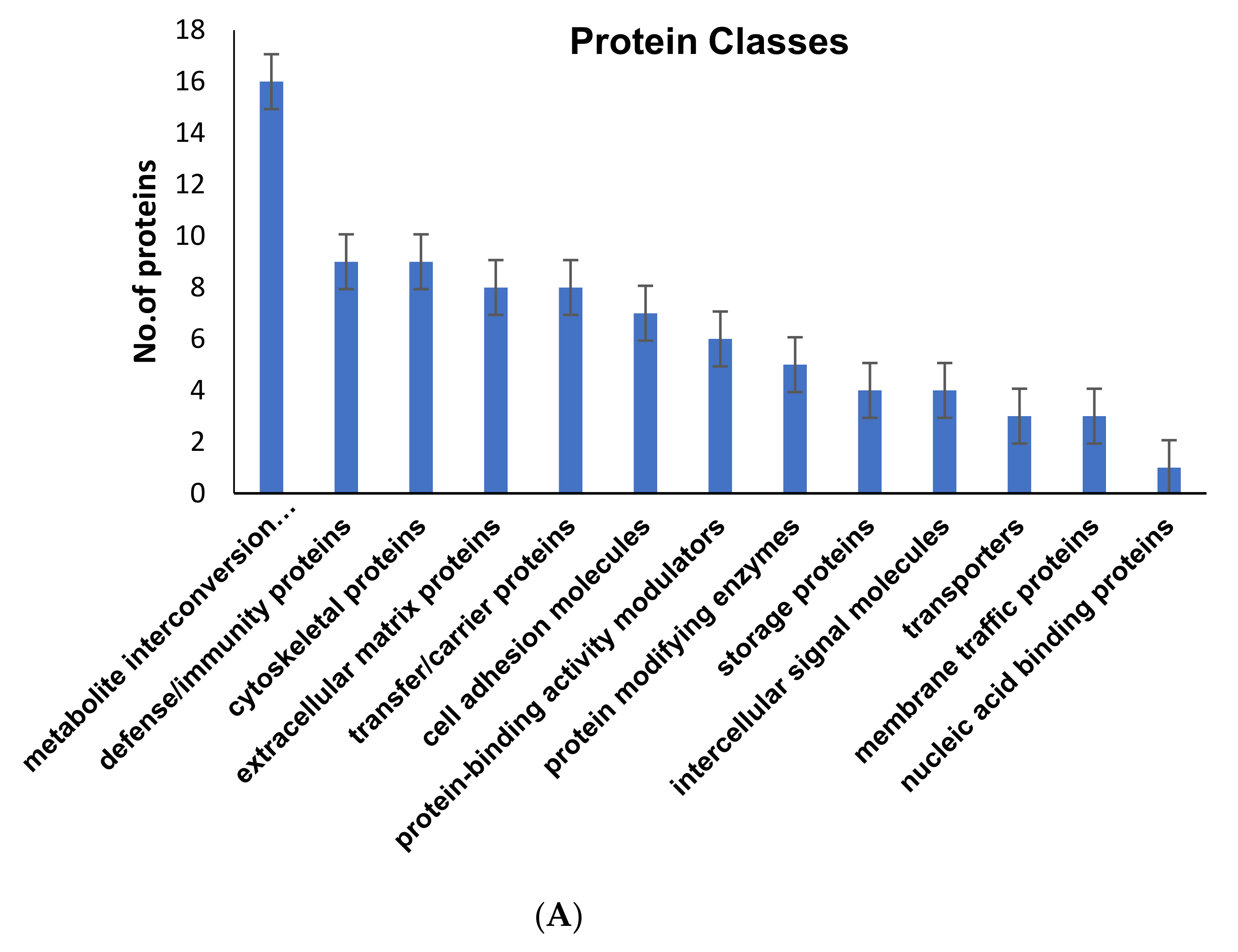
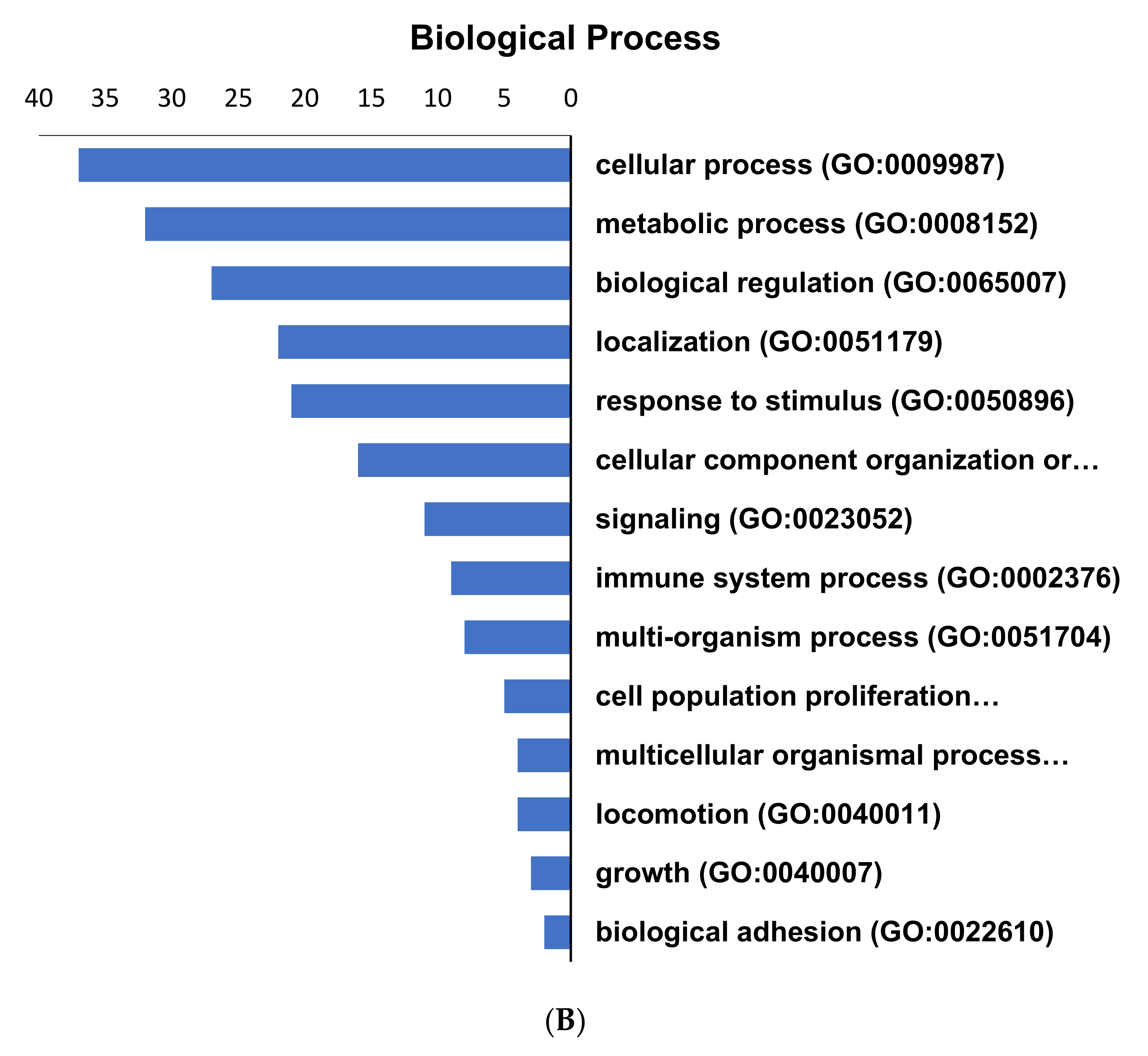
2.3. Pathways Regulated by the Altered Proteins from GCT
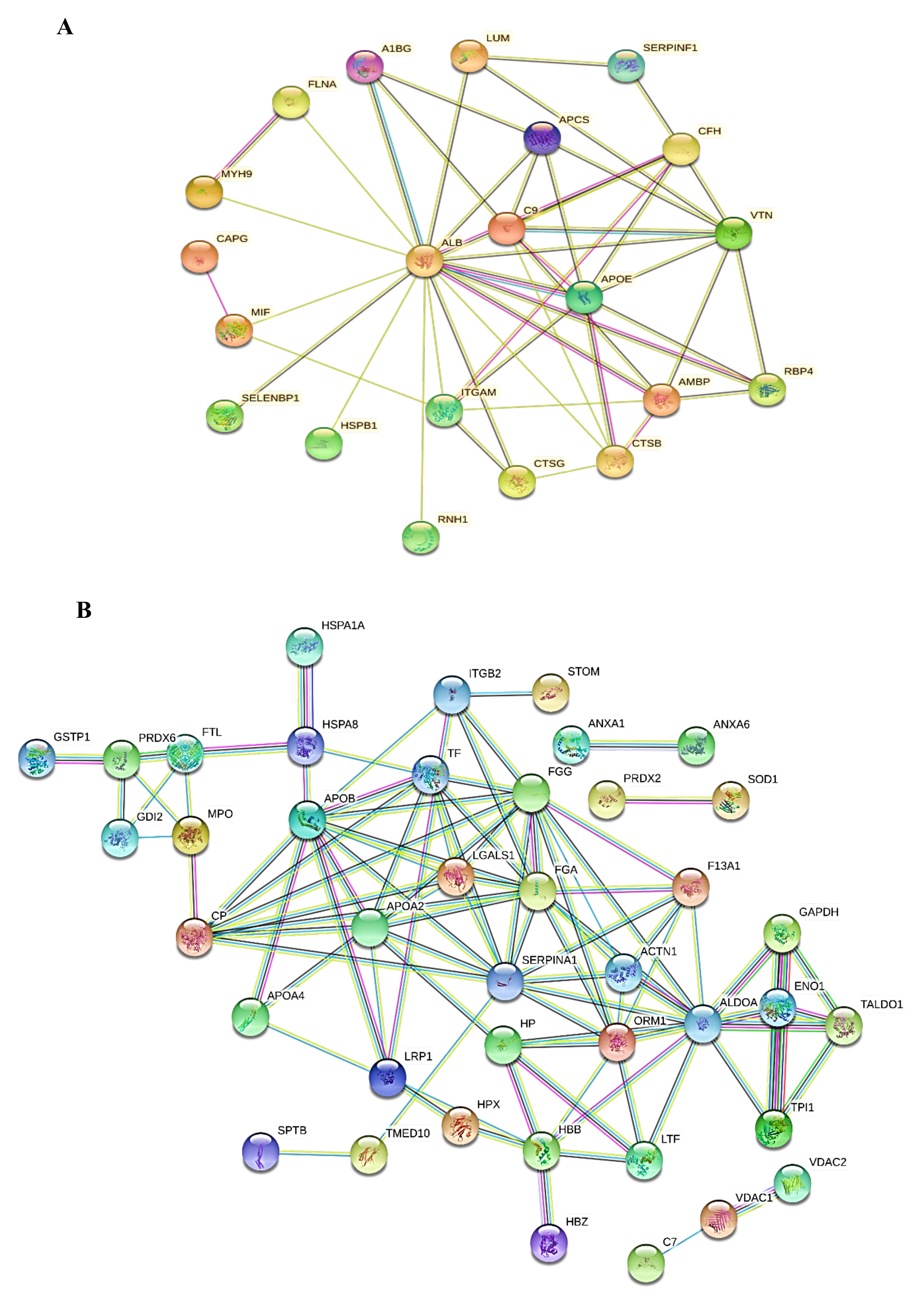
2.4. Protein-Protein Interaction Networks (PPI)
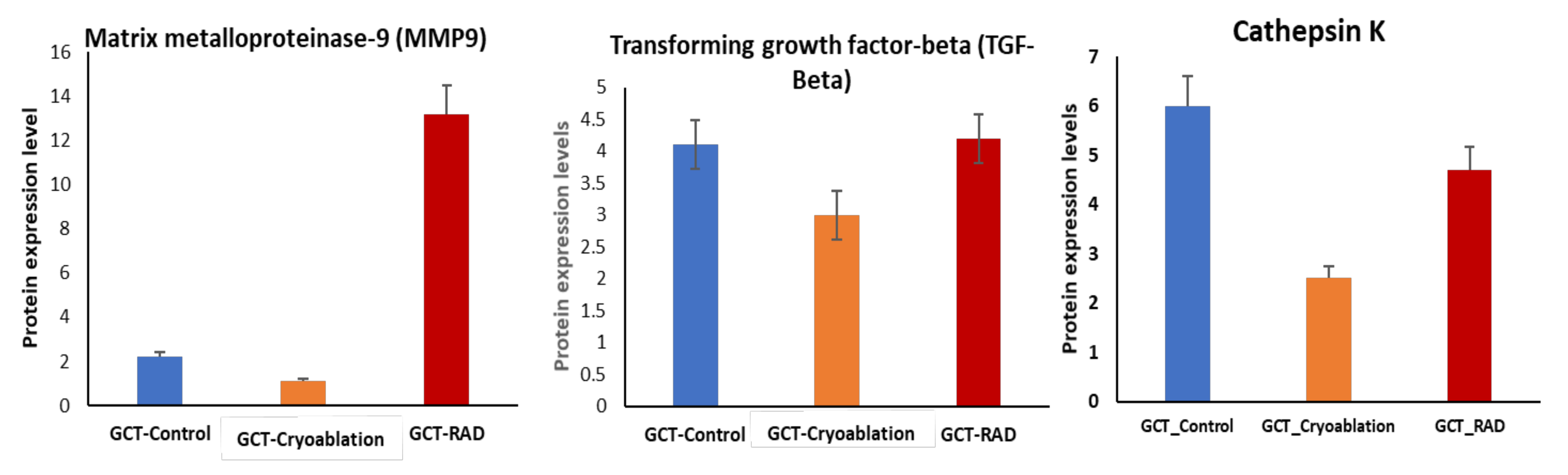
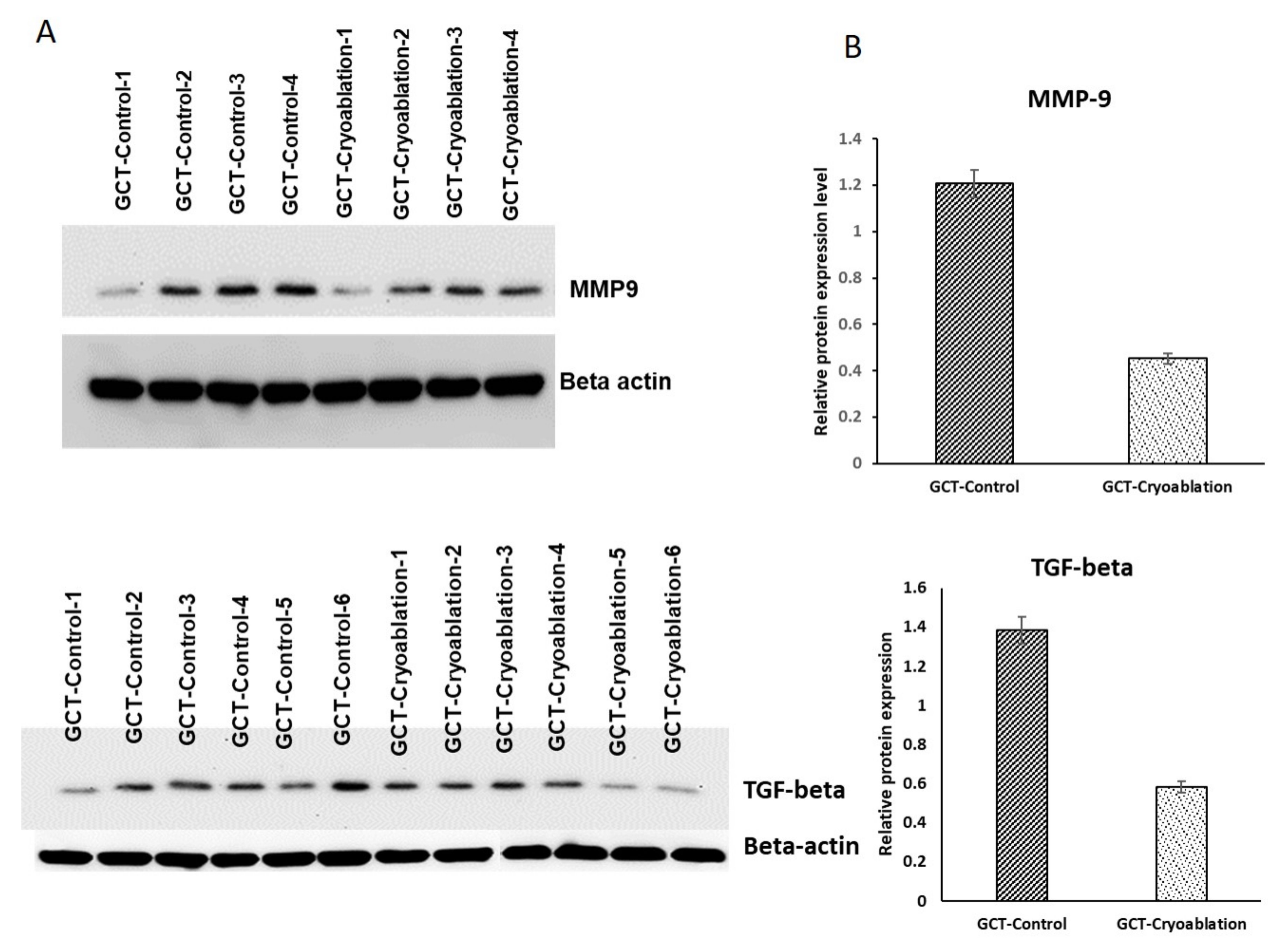
2.5. Validation of MMP-9 and TGF-Beta Protein Expression in GCT
3. Discussion
4. Materials and Methods
4.1. Patients and Clinical Information
4.2. Extraction of Protein from GCT Untreated/Control and Treated Samples Preparation
4.3. Protein Precipitation and In-Solution Digestion
4.4. Nano UPLC and Mass Spectrometry Conditions
4.5. Protein Quantification
4.6. Protein Identification
4.7. Bioinformatics Analysis
4.8. Statistical Analysis
4.9. Western Blot Analysis:
5. Conclusions
Limitations
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mnaymneh, A.W.; Ghandur, M.L. Giant-cell tumor of bone. Prog. Clin. Cancer 1967, 3, 245–280. [Google Scholar] [CrossRef]
- Sobti, A.; Agrawal, P.; Agarwala, S.; Agarwal, M. Giant Cell Tumor of Bone–An Overview. Arch. Bone Jt. Surg. 2016, 4, 2–9. [Google Scholar]
- Gouin, F.; Dumaine, V. Local recurrence after curettage treatment of giant cell tumors in peripheral bones: Retrospective study by the GSF-GETO (French Sarcoma and Bone Tumor Study Groups). Orthop. Traumatol. Surg. Res. 2013, 99, S313–S318. [Google Scholar] [CrossRef] [Green Version]
- Palmerini, E.; Picci, P.; Reichardt, P.; Downey, G. Malignancy in Giant Cell Tumor of Bone: A Review of the Literature. Technol. Cancer Res. Treat. 2019, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skubitz, K.M. Giant Cell Tumor of Bone: Current Treatment Options. Curr. Treat. Options Oncol. 2014, 15, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Werner, M. Giant cell tumour of bone: Morphological, biological and histogenetical aspects. Int. Orthop. 2006, 30, 484–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbeitsgemeinschaft, K.; Becker, W.T.; Dohle, J.; Bernd, L.; Braun, A.; Cserhati, M.; Enderle, A.; Hovy, L.; Matejovsky, Z.; Szendroi, M.; et al. Local recurrence of giant cell tumor of bone after intralesional treatment with and without adjuvant therapy. J. Bone Jt. Surg. Am. 2008, 90, 1060–1067. [Google Scholar]
- Bickels, J.; Campanacci, D.A. Local Adjuvant Substances Following Curettage of Bone Tumors. J. Bone Jt. Surg.-Am. Vol. 2020, 102, 164–174. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Bassiony, A.A.; Shalaby, H.; Assal, M.K. Cryosurgery and Impaction Subchondral Bone Graft for the Treatment of Giant Cell Tumor Around the Knee. HSS J. 2009, 5, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Dabak, N.; Tomak, Y.; Piskin, A.; Gulman, B.; Ozcan, H. Early results of a modified technique of cryosurgery. Int. Orthop. 2003, 27, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Baust, J.M.A.; Gage, A.; Clarke, D.; Van Buskirk, R. Cryosurgery—A putative approach to molecular-based optimization. Cryobiology 2004, 48, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Cooper, S.M.; Dawber, R.P.R. The History of Cryosurgery. J. R. Soc. Med. 2001, 94, 196–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freezing out cancer. Cryoablation could be a potential new treatment for a wider variety of cancers. DukeMed Health 2010, 16, 3.
- Wittig, J.C.; Simpson, B.M.; Bickels, J.; Kellar-Graney, K.L.; Malawer, M.M. Giant cell tumor of the hand: Superior results with curettage, cryosurgery, and cementation. J. Hand Surg. 2001, 26, 546–555. [Google Scholar] [CrossRef] [Green Version]
- Madda, R.; Chen, C.-M.; Wang, J.-Y.; Chen, C.-F.; Chao, K.-Y.; Yang, Y.-M.; Wu, H.-Y.; Chen, W.-M.; Wu, P.-K. Proteomic profiling and identification of significant markers from high-grade osteosarcoma after cryotherapy and irradiation. Sci. Rep. 2020, 10, 2105–2118. [Google Scholar] [CrossRef]
- Mukaihara, K.; Suehara, Y.; Kohsaka, S.; Akaike, K.; Tanabe, Y.; Kubota, D.; Ishii, M.; Fujimura, T.; Kazuno, S.; Okubo, T.; et al. Protein Expression Profiling of Giant Cell Tumors of Bone Treated with Denosumab. PLoS ONE 2016, 11, e0148401. [Google Scholar] [CrossRef] [Green Version]
- Conti, A.; Rodriguez, G.C.; Chiechi, A.; Blazquez, R.M.D.; Barbado, V.; Krènacs, T.; Novello, C.; Pazzaglia, L.; Quattrini, I.; Zanella, L.; et al. Identification of Potential Biomarkers for Giant Cell Tumor of Bone Using Comparative Proteomics Analysis. Am. J. Pathol. 2011, 178, 88–97. [Google Scholar] [CrossRef]
- Schenker, T.; Trueb, B. Down-Regulated proteins of mesenchymal tumor cells. Exp. Cell Res. 1998, 239, 161–168. [Google Scholar] [CrossRef]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular Matrix Degradation and Remodeling in Development and Disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Ueda, Y.; Imai, K.; Tsuchiya, H.; Fujimoto, N.; Nakanishi, I.; Katsuda, S.; Seiki, M.; Okada, Y. Matrix metalloproteinase 9 (gelatinase B) is expressed in multinucleated giant cells of human giant cell tumor of bone and is associated with vascular invasion. Am. J. Pathol. 1996, 148, 611–622. [Google Scholar] [PubMed]
- Shingleton, W.D.; Cawston, T.E.; Hodges, D.J.; Brick, P. Collagenase: A key enzyme in collagen turnover. Biochem. Cell Biol. 1996, 74, 759–775. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, K.; Nishimura, R.; Senn, J.; Youssef, R.F.; London, S.D.; Reddy, S.V. RANK ligand signaling modulates the matrix metalloproteinase-9 gene expression during osteoclast differentiation. Exp. Cell Res. 2007, 313, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiao, J.; Zhang, H.; Zhou, W.; Li, Z.; Han, S.; Wang, J.; Yang, X.; Huang, Q.; Wu, Z.; et al. TGF-β induced PAR-1 expression promotes tumor progression and osteoclast differentiation in giant cell tumor of bone. Int. J. Cancer 2017, 141, 1630–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindeman, J.H.; Hanemaaijer, R.; Mulder, A.; Dijkstra, P.S.; Szuhai, K.; Brömme, D.; Verheijen, J.H.; Hogendoorn, P.C.W. Cathepsin K Is the Principal Protease in Giant Cell Tumor of Bone. Am. J. Pathol. 2004, 165, 593–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.; Petrow, P.K.; Gaumann, A.; Keyszer, G.M.; Otto, M.; Kirkpatrick, C.J.; Bräuer, R. Expression of cysteine proteinases cathepsins b and k and of cysteine proteinase inhibitor cystatin c in giant cell tumor of tendon sheath. Mod. Pathol. 2001, 14, 318–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Younusi, A.; Cao, L.; Tian, Z.; Zhou, Y.; Song, X. Potential role of heat-shock proteins in giant cell tumors. Genet. Mol. Res. 2015, 14, 19144–19154. [Google Scholar] [CrossRef]
- Elamin, Y.Y.; Rafee, S.; Osman, N.; O′byrne, K.J.; Gately, K. Thymidine Phosphorylase in Cancer; Enemy or Friend? Cancer Microenviron. 2016, 9, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Haigis, M.C. Ribonuclease inhibitor as an intracellular sentry. Nucleic Acids Res. 2003, 31, 1024–1032. [Google Scholar] [CrossRef] [Green Version]
- Becerra, S.P.; Notario, V. The effects of PEDF on cancer biology: Mechanisms of action and therapeutic potential. Nat. Rev. Cancer 2013, 13, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Kornberg, M.D. The immunologic Warburg effect: Evidence and therapeutic opportunities in autoimmunity. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganapathy-Kanniappan, S.; Geschwind, J.F. Tumor glycolysis as a target for cancer therapy: Progress and prospects. Mol. Cancer 2013, 12, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacruz-López, D.; Griselda, K.; Castro-Muñoz, L.J.; Reyes-Hernández, D.O.; García-Carrancá, A.; Manzo Merino, J. Lactate in the regulation of tumor microenvironment and therapeutic approaches. Front. Oncol. 2019, 9, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.-Y.; Yang, Y.-C.; Ting, K.-L.; Wen, S.-Y.; Padma, V.V.; Huang, C.-Y.; Kuo, W.-W. Lactate dehydrogenase downregulation mediates the inhibitory effect of diallyl trisulfide on proliferation, metastasis, and invasion in triple-negative breast cancer. Environ. Toxicol. 2016, 32, 1390–1398. [Google Scholar] [CrossRef]
- Avnet, S.; Lemma, S.; Errani, C.; Falzetti, L.; Panza, E.; Columbaro, M.; Nanni, C.; Baldini, N. Benign albeit glycolytic: MCT4 expression and lactate release in giant cell tumour of bone. Bone 2020, 134, 115302. [Google Scholar] [CrossRef]
- Shih, N.-Y.; Lai, H.-L.; Chang, G.-C.; Lin, H.-C.; Wu, Y.-C.; Liu, J.M.; Liu, K.-J.; Tseng, S.-W. Anti-enolase Autoantibodies Are Down-regulated in Advanced Cancer Patients. Jpn. J. Clin. Oncol. 2010, 40, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Aarts, B.M.; Klompenhouwer, E.G.; Rice, S.L.; Imani, F.; Baetens, T.R.; Bex, A.; Horenblas, S.; Kok, M.; Haanen, J.B.A.G.; Beets-Tan, R.G.H.; et al. Cryoablation and immunotherapy: An overview of evidence on its synergy. Insights Imaging 2019, 10, 53. [Google Scholar] [CrossRef]
- Gage, A.; Baust, J. Experimental cryosurgery investigations in vivo. Cryobiology 2009, 59, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Cortés-Ríos, J.; Zárate, A.M.; Figueroa, J.D.; Medina, J.; Fuentes-Lemus, E.; Rodríguez-Fernández, M.; Aliaga, M.E.; López-Alarcón, C. Protein quantification by bicinchoninic acid (BCA) assay follows complex kinetics and can be performed at short incubation times. Anal. Biochem. 2020, 113904. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, H.; Zhang, J.; Boldogh, I.; Zhao, Y. Quantitative Assessment of the Effects of Trypsin Digestion Methods on Affinity Purification–Mass Spectrometry-based Protein–Protein Interaction Analysis. J. Proteome Res. 2017, 16, 3068–3082. [Google Scholar] [CrossRef]
- Erve, J.C.L.; DeMaio, W.; Talaat, R.E. Rapid metabolite identification with sub parts-per-million mass accuracy from biological matrices by direct infusion nanoelectrospray ionization after clean-up on a ZipTip and LTQ/Orbitrap mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 3015–3026. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G.A. PEAKS: Powerful software for peptidede novo sequencing by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef] [PubMed]
- Megger, D.A.; Bracht, T.; Meyer, H.E.; Sitek, B. Label-free quantification in clinical proteomics. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2013, 1834, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Yadav, A.K. False Discovery Rate Estimation in Proteomics. Adv. Struct. Saf. Stud. 2016, 1362, 119–128. [Google Scholar]
- Tran, N.H.; Qiao, R.; Xin, L.; Chen, X.; Liu, C.; Zhang, X.; Shan, B.; Ghodsi, A.; Li, M. Deep learning enables de novo peptide sequencing from data-independent-acquisition mass spectrometry. Nat. Methods 2019, 16, 63–66. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, J.; Wu, S.; Li, D.; Zhu, Y.; He, F. Improving the sensitivity of MASCOT search results validation by combining new features with Bayesian nonparametric model. Proteomics 2010, 10, 4293–4300. [Google Scholar] [CrossRef]
- Soudy, M.; Anwar, A.M.; Ahmed, E.A.; Osama, A.; Ezzeldin, S.; Mahgoub, S.; Magdeldin, S. UniprotR: Retrieving and visualizing protein sequence and functional information from Universal Protein Resource (UniProt knowledgebase). J. Proteom. 2020, 213, 103613. [Google Scholar] [CrossRef]
- The UniProt Consortium UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef] [Green Version]
- Sharma, S.; Ciufo, S.; Starchenko, E.; Darji, D.; Chlumsky, L.; Karsch-Mizrachi, I.; Schoch, C.L. The NCBI BioCollections Database. Database 2019, 2019. [Google Scholar] [CrossRef]
- Zhou, X.; Su, Z. EasyGO: Gene Ontology-based annotation and functional enrichment analysis tool for agronomical species. BMC Genom. 2007, 8, 246. [Google Scholar] [CrossRef]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [PubMed]
- Algina, J.; Olejnik, S. Conducting Power Analyses for Anova and Ancova in between-Subjects Designs. Eval. Health Prof. 2003, 26, 288–314. [Google Scholar] [CrossRef]
- TBST for Western Blotting. Cold Spring Harb. Protoc. 2013, 2013, 074104. [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Protein Name a | Significance | Coverage (%) | #Peptides | #Unique | GCT_Untreated//Control b | GCT_Cryoablation b | GCT_RAD | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| FLNA_HUMAN | Filamin | 200 | 12 | 7 | 6 | 1.763093 | −1.42652 | −3.1814 | 5.24 × 10−6 |
| PDL1_HUMAN | Programmed cell death ligand 1 | 200 | 91 | 81 | 18 | 1.15715 | −0.92406 | −2.84759 | 0.0005 |
| LUM_HUMAN | Lumican | 200 | 21 | 2 | 1 | 1.060151 | 0.151582 | −1.45149 | 0.001 |
| ALBU_HUMAN | ALBUMIN | 200 | 17 | 15 | 15 | 1.040816 | 0.673168 | −1.83586 | 0.001 |
| THRB_HUMAN | Prothrombin | 200 | 11 | 6 | 4 | 0.911502 | 0.542668 | −3.46715 | 0.04 |
| CO6A3_HUMAN | Collagen alpha-3(VI) chain | 200 | 61 | 20 | 1 | 0.789581 | 0.609516 | −1.85642 | 0.01 |
| BLVRB_HUMAN | Flavin reductase (NADPH) | 200 | 14 | 8 | 5 | 0.796578 | 0.568552 | −0.76413 | 0.04 |
| SBP1_HUMAN | Methanethiol oxidase | 200 | 15 | 9 | 3 | 0.691594 | 0.360094 | −1.34739 | 0.00012 |
| HBA_HUMAN | Hemoglobin subunit alpha | 200 | 18 | 5 | 1 | 0.650356 | 0.159428 | −0.585 | 0.04 |
| HBB_HUMAN | Hemoglobin subunit beta | 200 | 8 | 5 | 1 | 0.712071 | 0.219155 | −0.34253 | 0.0023 |
| VTNC_HUMAN | Vitronectin | 200 | 18 | 5 | 1 | 0.659152 | 0.04037 | −0.24675 | 0.004 |
| MMP9_HUMAN | Matrix metalloproteinase 9 | 200 | 8 | 5 | 1 | 0.63959 | −0.1886 | −0.58489 | 0.01 |
| CATB_HUMAN | Cathepsin B | 200 | 15 | 5 | 1 | 0.626248 | −0.6555 | −0.93563 | 0.0007 |
| SAMP_HUMAN | Serum amyloid P-component | 200 | 8 | 5 | 1 | 0.454084 | 0.310969 | −1.43804 | 0.02 |
| ITAM_HUMAN | Integrin alpha-M | 200 | 5 | 3 | 1 | 0.388519 | 0.57785 | −2.65341 | 0.04 |
| CSF1_HUMAN | Colony stimulating factor 1 | 200 | 11 | 4 | 1 | 0.014655 | 0.751954 | −2.10563 | 0.04 |
| A1BG_HUMAN | Alpha-1B-glycoprotein | 200 | 11 | 4 | 1 | 0.381938 | 0.742658 | −1.48566 | 0.01 |
| KV315_HUMAN | Immunoglobulin kappa variable 3–15 | 200 | 11 | 4 | 1 | 0.175186 | 0.796431 | −1.21047 | 0.005 |
| FETUA_HUMAN | Alpha-2-HS-glycoprotein | 200 | 97 | 24 | 1 | 0.46504 | 0.201584 | 0.03433 | 0.0004 |
| CSF1R_HUMAN | Colony stimulating factor receptor | 200 | 97 | 24 | 1 | 0.107399 | 0.678371 | 0.000169 | 0.00061 |
| A1AT_HUMAN | Alpha-1-antitrypsin | 200 | 97 | 24 | 1 | 0.50398 | 0.972984 | −1.73774 | 0.0004 |
| HV315_HUMAN | Immunoglobulin heavy variable 3–15 | 200 | 0 | 1 | 1 | 0.441371 | 1.29497 | −4.54668 | 0.00014 |
| A2MG_HUMAN | Alpha-2-macroglobulin | 200 | 37 | 15 | 5 | 0.067335 | 1.38741 | −2.57057 | 0.00058 |
| TRFE_HUMAN | Serotransferrin | 200 | 50 | 15 | 5 | 0.050994 | 1.028623 | −0.85167 | 0.00025 |
| Accession | Protein Name a | Significance | Coverage (%) | #Peptides | #Unique | GCT_Untreated/Control b | GCT_Cryoablation b | GCT_RAD b | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| ENOA_HUMAN | Alpha-enolase | 200 | 50 | 15 | 5 | −0.18695 | 1.127769 | −0.52723 | 0.01 |
| FHL1_HUMAN | Four and a half LIM domains protein 1 | 200 | 32 | 12 | 2 | −0.11059 | 0.930268 | −0.54024 | 0.04 |
| IGHG3_HUMAN | Immunoglobulin heavy constant gamma 3 | 200 | 18 | 3 | 1 | −0.10647 | 0.956033 | −0.22101 | 0.001 |
| IGHA1_HUMAN | Immunoglobulin heavy constant alpha 1 | 200 | 28 | 13 | 1 | −0.50502 | 1.027274 | −0.39062 | 0.001 |
| HPT_HUMAN | Haptoglobin | 200 | 29 | 6 | 5 | −0.58019 | 0.927425 | −0.23422 | 0.002 |
| PRDX2_HUMAN | Peroxiredoxin-2 | 200 | 21 | 6 | 6 | −0.24551 | 0.767434 | −0.03834 | 0.01 |
| IGHG2_HUMAN | Immunoglobulin heavy constant gamma 2 | 200 | 1 | 1 | 1 | −0.42138 | 0.601606 | −0.50063 | 0.0000065 |
| DPYL2_HUMAN | Dihydropyrimidinase-related protein 2 | 200 | 1 | 1 | 1 | −0.53341 | 0.671534 | −0.41069 | 0.04 |
| FINC_HUMAN | Fibronectin | 200 | 3 | 2 | 2 | −0.81512 | 0.7081 | −0.23971 | 0.02 |
| TRFL_HUMAN | Lactotransferrin/Growth-inhibiting protein 12 | 200 | 1 | 1 | 1 | −1.32309 | 0.707061 | −0.04577 | 0.04 |
| IGJ_HUMAN | Immunoglobulin J chain | 200 | 28 | 9 | 5 | −0.7484 | 0.69038 | 0.149366 | 0.00032 |
| FLNB_HUMAN | Filamin-B | 200 | 3 | 2 | 1 | −0.79679 | 0.708131 | 0.103967 | 0.00000561 |
| GDIB_HUMAN | Rab GDP dissociation inhibitor beta | 200 | 11 | 16 | 6 | −0.63366 | 0.523302 | 0.435171 | 0.01 |
| CATD_HUMAN | Cathepsin D | 200 | 18 | 2 | 2 | 0.137456 | 0.13756 | 0.412715 | 0.0005 |
| KAP0_HUMAN | cAMP-dependent protein kinase type I-alpha regulatory subunit | 200 | 8 | 2 | 2 | −0.19286 | 0.249868 | 0.616559 | 0.0007 |
| CLIC1_HUMAN | Chloride intracellular channel protein 1 | 200 | 13 | 11 | 1 | −0.38777 | −0.32961 | 0.719213 | 0.0000024 |
| ALDOA_HUMAN | Fructose-bisphosphate aldolase A/Lung cancer antigen NY-LU-1 | 200 | 3 | 12 | 1 | −0.65764 | −0.40178 | 0.848804 | 0.003 |
| TPIS_HUMAN | Triosephosphate isomerase | 200 | 7 | 14 | 1 | −0.82482 | −0.31639 | 0.655647 | 0.04 |
| PGK1_HUMAN | Phosphoglycerate kinase 1 | 200 | 2 | 16 | 1 | −0.85269 | −0.46433 | 0.613938 | 0.00012 |
| FIBA_HUMAN | Fibrinogen alpha chain | 200 | 4 | 1 | 8 | −1.10845 | −0.66497 | 0.757142 | 0.04 |
| HBAZ_HUMAN | Hemoglobin subunit zeta | 200 | 3 | 4 | 2 | −0.96093 | 0.237888 | 0.116009 | 0.0023 |
| IGHG4_HUMAN | Immunoglobulin heavy constant gamma 4 | 200 | 13 | 1 | 1 | −1.11525 | 0.2758 | 0.094595 | 0.004 |
| ANXA1_HUMAN | Annexin A1/p35 | 200 | 3 | 1 | 1 | −1.25595 | 0.194197 | 0.232402 | 0.01 |
| AMBP_HUMAN | Protein AMBP | 200 | 3 | 1 | 1 | −0.00093 | 0.172501 | −2.48943 | 0.0007 |
| A1AG1_HUMAN | Alpha-1-acid glycoprotein 1 | 200 | 23 | 14 | 5 | −0.59995 | 0.077871 | −0.81155 | 0.02 |
| S10A9_HUMAN | Protein S100-A9/Migration inhibitory factor-related protein 14 | 200 | 3 | 7 | 1 | −0.73456 | 0.257952 | −0.74316 | 0.04 |
| PERM_HUMAN | Myeloperoxidase | 200 | 14 | 1 | 1 | −2.42652 | 0.801777 | −1.24608 | 0.04 |
| GTR1_HUMAN | Solute carrier family 2, facilitated glucose transporter member 1 | 200 | 4 | 5 | 1 | −1.02121 | −0.12839 | −0.27789 | 0.01 |
| TPM3_HUMAN | 200 | 5 | 16 | 1 | −0.91326 | −0.40327 | −0.41174 | 0.005 | |
| CAPG_HUMAN | Macrophage-capping protein | 200 | 15 | 3 | 1 | −1.29189 | −1.04556 | −3.08074 | 0.0004 |
| ITIH4_HUMAN | Inter-alpha-trypsin inhibitor heavy chain H4 | 200 | 13 | 12 | 6 | −2.61728 | −0.57445 | −0.26668 | 0.00061 |
| FRIL_HUMAN | Ferritin light chain | 200 | 3 | 7 | 1 | −2.54766 | −0.96978 | 0.245989 | 0.0004 |
| B3AT_HUMAN | Band 3 anion transport protein | 200 | 3 | 14 | 1 | −2.00353 | 0.106701 | −0.41339 | 0.00014 |
| STOM_HUMAN | Erythrocyte band 7 integral membrane protein | 200 | 3 | 1 | 4 | −2.4425 | 0.156164 | −0.67122 | 0.00058 |
| AACT_HUMAN | Alpha-1-antichymotrypsin | 200 | 32 | 6 | 1 | −1.34745 | 0.192083 | −0.0366 | 0.00025 |
| IGG1_HUMAN | Immunoglobulin gamma-1 heavy chain | 200 | 22 | 4 | 1 | −1.99553 | 0.112038 | 0.194619 | 0.00014 |
| APOB_HUMAN | Apolipoprotein B-100 | 200 | 4 | 2 | 2 | −1.88271 | 0.42193 | 0.588524 | 0.000056 |
| PEBP1_HUMAN | Phosphatidylethanolamine-binding protein 1 | 200 | 8 | 1 | 1 | −1.92032 | 0.549155 | 0.384127 | 0.00047 |
| CAH2_HUMAN | Carbonic anhydrase 2 | 200 | 7 | 1 | 1 | −1.91215 | −0.23354 | 0.693737 | 0.00014 |
| GSTP1_HUMAN | Glutathione S-transferase P | 200 | 7 | 1 | 1 | −4.03432 | −0.08147 | 0.859662 | 0.0056 |
| PPIA_HUMAN | Peptidyl-prolyl cis-trans isomerase A | 200 | 16 | 2 | 1 | −0.6593 | 0.020047 | 0.843733 | 0.00014 |
| F13A_HUMAN | Coagulation factor XIII A chain | 200 | 16 | 3 | 3 | −0.77574 | 0.216645 | 0.714859 | 0.00061 |
| SODC_HUMAN | Superoxide dismutase [Cu-Zn] | 200 | 10 | 2 | 2 | −0.85644 | 0.197895 | 0.714553 | 0.0004 |
| SPTB1_HUMAN | Spectrin beta chain, erythrocytic | 200 | 28 | 9 | 5 | −0.71009 | −0.04636 | 1.052198 | 0.00014 |
| HVM17_HUMAN | Ig heavy chain V region MOPC 47A | 200 | 3 | 2 | 1 | −1.46988 | 0.246253 | 1.037756 | 0.00058 |
| VDAC2_HUMAN | Voltage-dependent anion-selective channel protein 2 | 200 | 1 | 1 | 1 | −1.07957 | −0.27273 | 1.016574 | 0.00025 |
| HSP7C_HUMAN | Heat shock cognate 71 kDa protein | 200 | 18 | 2 | 2 | −1.87161 | −0.55349 | 1.177799 | 0.04 |
| PGS2_HUMAN | Decorin/Bone proteoglycan II | 200 | 18 | 2 | 2 | −0.37748 | −1.37542 | 1.357619 | 0.04 |
| VAT1_HUMAN | Synaptic vesicle membrane protein VAT-1 homolog | 200 | 3 | 1 | 1 | −1.69165 | −0.99373 | 1.390167 | 0.01 |
| COF1_HUMAN | Cofilin-1 | 200 | 3 | 1 | 3 | −2.07774 | −0.98897 | 1.379402 | 0.002 |
| IF4A1_HUMAN | Eukaryotic initiation factor 4A-I/ATP-dependent RNA helicase eIF4A-1 | 200 | 7 | 1 | 1 | −1.75171 | −1.79621 | 1.711834 | 0.0005 |
| NUCL_HUMAN | Nucleolin | 200 | 2 | 1 | 1 | −2.86071 | −2.53729 | 1.807236 | 0.003 |
| BIP_HUMAN | Endoplasmic reticulum chaperone BiP/Heat shock protein 70 family protein 5 | 200 | 4 | 1 | 5 | −2.00732 | −1.73829 | 1.556309 | 0.00061 |
| GANAB_HUMAN | Neutral alpha-glucosidase AB | 200 | 3 | 1 | 1 | −2.49319 | −2.45966 | 1.608849 | 0.0004 |
| APOA1_HUMAN | Apolipoprotein A-I | 200 | 13 | 5 | 8 | −3.31579 | −1.46294 | 1.466519 | 0.00014 |
| RL18_HUMAN | 60S ribosomal protein L18 | 200 | 3 | 1 | 1 | −3.85556 | −2.22158 | 1.512808 | 0.00058 |
| APOD_HUMAN | Apolipoprotein D | 200 | 28 | 9 | 5 | −1.7026 | −0.60243 | 1.034735 | 0.00025 |
| S10A4_HUMAN | Protein S100-A4 | 200 | 13 | 3 | 1 | −1.15342 | −1.16094 | 1.143946 | 0.04 |
| PRDX6_HUMAN | Peroxiredoxin-6 | 200 | 21 | 4 | 1 | −0.7977 | −1.20379 | 0.928545 | 0.04 |
| IDHP_HUMAN | Isocitrate dehydrogenase [NADP], mitochondrial | 200 | 18 | 2 | 2 | −1.7431 | −1.2689 | 1.137871 | 0.01 |
| RS21_BOVIN | 40S ribosomal protein S21 | 200 | 8 | 2 | 2 | −2.49295 | −1.47642 | 1.202521 | 0.0000006 |
| RS19_HUMAN | 40S ribosomal protein S19 | 200 | 3 | 1 | 1 | −2.55879 | −1.83901 | 1.212821 | 0.04 |
| CO4A_HUMAN | Complement C4-A | 200 | 3 | 1 | 1 | −2.79494 | −1.27031 | 1.102623 | 0.04 |
| LEG1_HUMAN | Galectin-1 | 200 | 7 | 1 | 1 | −2.88241 | −2.12528 | 1.046419 | 0.01 |
| PDIA1_HUMAN | Protein disulfide-isomerase/p55 | 200 | 22 | 4 | 1 | −3.6809 | −1.07449 | 1.012252 | 0.00061 |
| GELS_HUMAN | Gelsolin | 200 | 4 | 1 | 6 | −2.06544 | −0.18597 | 1.095377 | 0.0004 |
| ITB2_HUMAN | Integrin beta-2 | 200 | 3 | 1 | 1 | −1.86435 | −0.05573 | 1.100121 | 0.00014 |
| C4BPA_HUMAN | C4b-binding protein alpha chain | 200 | 3 | 1 | 1 | −2.41077 | 0.128627 | 1.05217 | 0.00058 |
| AOC3_HUMAN | Membrane primary amine oxidase | 200 | 3 | 1 | 1 | −2.95289 | 0.051086 | 0.980408 | 0.00025 |
| HBA_HUMAN | Hemoglobin subunit alpha | 200 | 3 | 1 | 2 | −3.05928 | −0.02117 | 1.105323 | 0.001 |
| Patient | Sex | Age | Location | Size of Tumor | Treatment | Reconstruction | Local Recurrence | Chemo & Radiation |
|---|---|---|---|---|---|---|---|---|
| 1 | F | 33 | Distal femur | 8 | Wide excision | recycle autograft | Yes | No |
| 2 | M | 36 | Proximal humerus | 5 | Curettage + cryoablation | recycle autograft | No | No |
| 3 | M | 45 | Proximal humerus | 7 | Wide excision | recycle autograft | Yes | No |
| 4 | F | 55 | Distal femur | 18 | Curettage + cryoablation | recycle autograft | No | No |
| 5 | F | 46 | Humeral shaft | 4 | Wide excision | recycle autograft | Yes | No |
| 6 | F | 54 | Distal femur | 6 | Curettage + cryoablation | recycle autograft | No | No |
| 7 | M | 47 | Acetabulum | 5.6 | Wide excision | bone grafting | Yes | No |
| 8 | M | 58 | Humeral Shaft | 7 | Wide excision | bone grafting | Yes | No |
| 9 | F | 38 | Acetabulum | 6 | Curettage + cryoablation | bone grafting | No | No |
| 10 | M | 63 | Distal femur | 18 | Curettage + cryoablation | total hip arthroplasty | No | No |
| 11 | F | 64 | Proximal humerus | 10 | Wide excision | bone grafting | Yes | No |
| 12 | M | 56 | Distal femur | 12 | Curettage + cryoablation | bone grafting | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madda, R.; Chen, C.-M.; Chen, C.-F.; Wang, J.-Y.; Wu, P.-K.; Chen, W.-M. Exploring the Proteomic Alterations from Untreated and Cryoablation and Irradiation Treated Giant Cell Tumors of Bone Using Liquid-Chromatography Tandem Mass Spectrometry. Molecules 2020, 25, 5355. https://doi.org/10.3390/molecules25225355
Madda R, Chen C-M, Chen C-F, Wang J-Y, Wu P-K, Chen W-M. Exploring the Proteomic Alterations from Untreated and Cryoablation and Irradiation Treated Giant Cell Tumors of Bone Using Liquid-Chromatography Tandem Mass Spectrometry. Molecules. 2020; 25(22):5355. https://doi.org/10.3390/molecules25225355
Chicago/Turabian StyleMadda, Rashmi, Chao-Ming Chen, Cheng-Fong Chen, Jir-You Wang, Po-Kuei Wu, and Wei-Ming Chen. 2020. "Exploring the Proteomic Alterations from Untreated and Cryoablation and Irradiation Treated Giant Cell Tumors of Bone Using Liquid-Chromatography Tandem Mass Spectrometry" Molecules 25, no. 22: 5355. https://doi.org/10.3390/molecules25225355
APA StyleMadda, R., Chen, C. -M., Chen, C. -F., Wang, J. -Y., Wu, P. -K., & Chen, W. -M. (2020). Exploring the Proteomic Alterations from Untreated and Cryoablation and Irradiation Treated Giant Cell Tumors of Bone Using Liquid-Chromatography Tandem Mass Spectrometry. Molecules, 25(22), 5355. https://doi.org/10.3390/molecules25225355