1. Introduction
Hepatocellular carcinoma (HCC) is a primary malignancy of the liver. Most HCCs are derived from a chronic viral hepatitis B infection (HBV or HCV) or alcoholic liver cirrhosis [
1,
2]. HCC is one of the fatal cancers and the second leading cause of cancer death worldwide. Liver cancer death is about 22.8% out of 100,000 population in South Korea according to the WHO in 2018 [
3]. Among many treatment options for HCC, complete tumor resection is one of the best potentially curative treatments. However, this curative treatment is frequently limited due to metastasis, the multiplicity of tumors, and the lack of hepatic reservoirs derived from underlying liver cirrhosis. Moreover, few effective systemic treatments are available for the inoperable HCC. Several chemotherapeutic drugs like cisplatin, 5-fluorouracil, and paclitaxel have been used in the treatment of HCC, but the cancer cells develop resistance with the continuous treatment of these drugs. Multi signaling-targeted agents have developed, but their efficacies are still limited [
4]. Therefore, the development of chemotherapeutic or chemo-preventive options is urgently required for the management of HCC.
Natural polyphenols like flavonoids are known to safely modulate the physiological function and enhance anti-cancer activities like curcumin, reservatrol, silymarin, quercetin, fisetin and deguelin, which is a common dietary compound found in many fruits and vegetables which showed anti-cancer activities in various cancer cells [
5,
6,
7]. Some of the phytochemicals have been used as drugs for millennia. Egyptian medicines dating back to 2900 BC “Ebers Papyrus” consist of around 700 medicines, mostly plant derived. Traditional Chinese medicine, which has been documented over thousands of years, and Indian Ayurveda medicines dating back to the first millennium BC specifies that most of the medicines used are of plant origin. [
8]. With the growth of ecological movements, natural products have become more popular for the prevention or treatment of cancer. In recent decades, conventional chemotherapy has induced too many toxicities with few anti-cancer effects, so there is a quest for a different paradigm of therapy, mainly focusing on phytotherapy or natural medicines.
Vitis coignetiae Pulliat (Meoru in Korea) is a fruit that was used as a Korean folk medicine to treat various diseases like cancer and inflammatory disorders. The dark red colored fruits contain anthocyanins in abundance which belong to the class of flavonoids. Many studies have described the anti-cancer effects of anthocyanins regarding anti-angiogenesis and cancer invasion [
9,
10]. Anthocyanins isolated from
Vitis coignetiae Pulliat (AIMs) is a group of anthocyanins which consists of delphinidin, cyaniding, petunidin, and malvidin. We previously demonstrated that the AIMs induce apoptosis by modulating Bcl-2 family and IAP family members in hepatocellular carcinoma cells and leukemia cells. [
9,
10]. In addition, AIMs induced anti-invasive and anti-metastatic effects by suppressing the NF-κB pathway in HT-29 human colon cancer cells and MCF-7 human breast cancer cells [
11,
12]. Even though the efficacies and phenotypes of the anti-cancer effects of AIMs vary according to the cancer cell types, AIMs inhibited NF-κB activity and the downstream molecules involved in cancer proliferation, anti-apoptosis, invasion and metastasis [
13]. However, many researchers argued that the concentrations of AIMs were too high for in vivo experiments, and the toxicities to normal cells. Therefore, we here investigated the effects of AIMs in vitro at a lower concentration on Hep3B cells with or without TNF-α, as TNF-α treatments augmented indicates the advanced clinical stage of cancer. We also validated their effects in in vivo experiments on a Xenograft model of Hep3B human hepatocellular carcinoma cells to establish the relevance between the in vitro and in vivo results.
3. Discussion
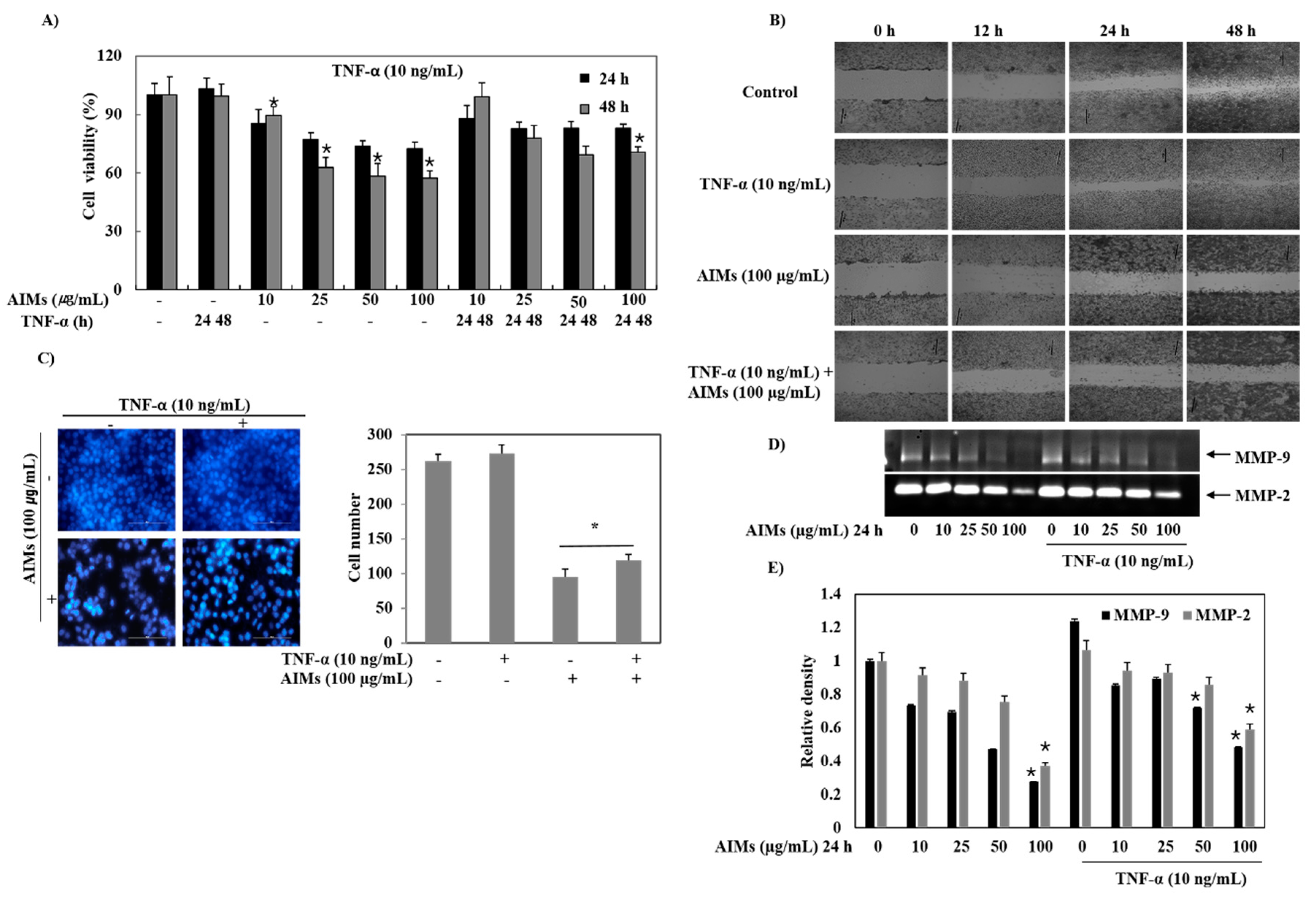
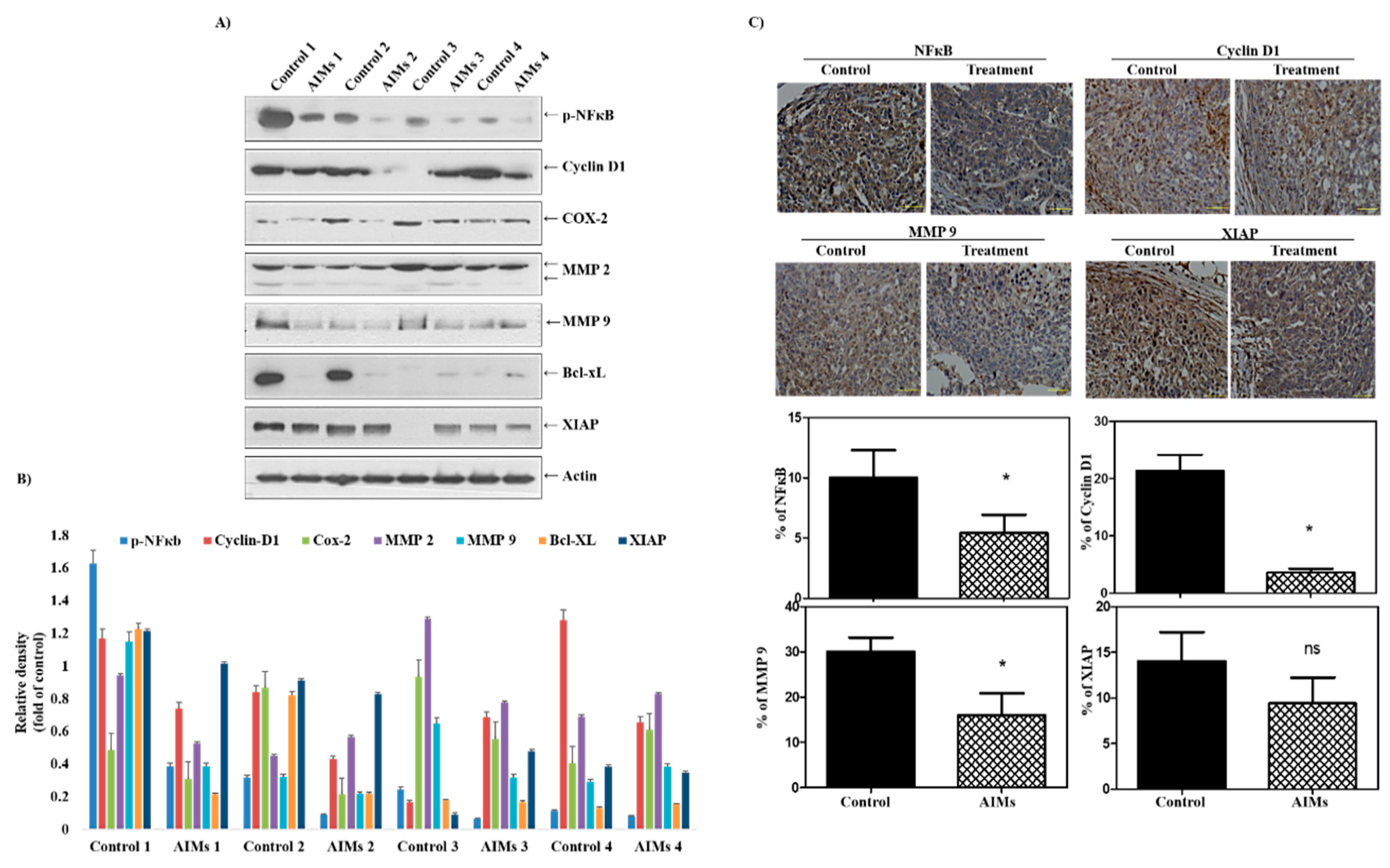
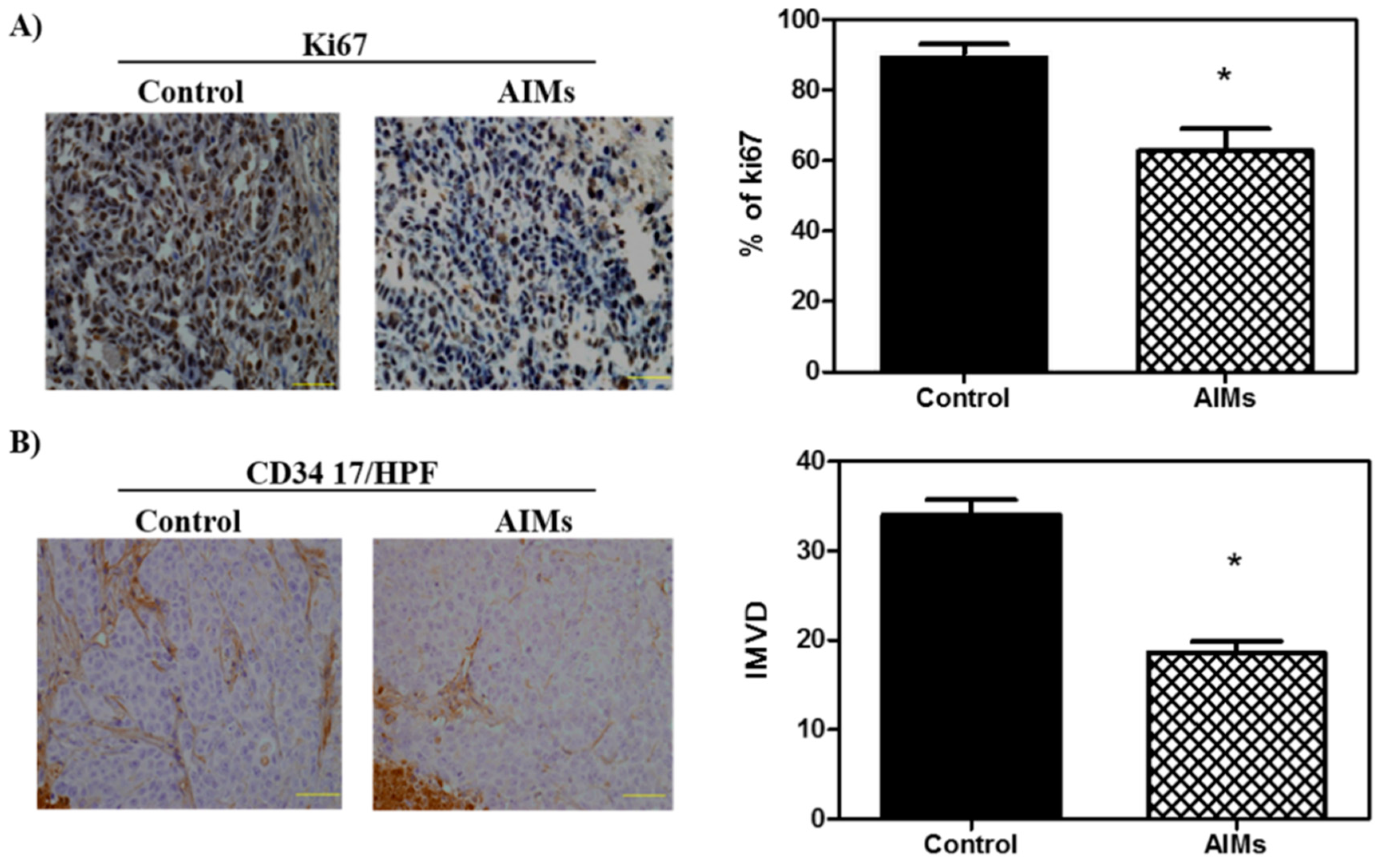
This study was designed to confirm that the anti-cancer effects of AIMs on cancer cells are valid in the in vivo system. We chose Hep3B human hepatocellular carcinoma cells in vitro and in vivo because regarding the effects of AIMs on Hep3B cells in vitro, reviewers doubted whether AIMs safely induced anti-cancer effects in vivo. In an in vitro study at lower doses (10–100 μg/mL), we found that AIMs still have anti-cancer effects in terms of proliferation, migration and invasion. However, the anti-cancer effects of AIMs were significantly diminished by TNF-α treatment in terms of proliferation. Before obtaining the in vivo study results, we were concerned that the results in the in vivo system would be different from those in in vitro system because the TNF-α-like cytokines may work in in vivo systems. Unlike our expectations, the results in vivo clearly demonstrated that AIMs harbor anti-proliferative, anti-invasive, and anti-angiogenic effects by inhibiting NF-κB activity, although the tumor sizes in control groups vary. The Western blot findings were especially well matched to the results of AIMs at high concentrations (100–400 μg/mL) [
11,
12]. Even though the daily dose of the in vivo study was not sufficient to attain the plasma level of 100–400 μg/mL of AIMs, the exposed cumulation may produce anti-cancer effects that are attained at high dose in the in vitro study. Actually, we could not get the clear inhibition of NF-κB activity with 100 μg/mL of AIMs (data not shown). This is the first study that demonstrated that AIMs inhibited cancer proliferation by inhibiting NF-κB activity and its downstream molecules involved in cancer proliferation, invasion, and angiogenesis in in vivo systems [
13]. In this in vivo study, we used clinical biomarkers for proliferation and angiogenesis (Ki67 proliferative index and IMVD) that are frequently used in clinical tumor samples. Antigen Ki-67 is a nuclear protein that is associated with cellular proliferation and an excellent marker to determine the growth fraction of a given cell population [
14,
15]. IMVD has also been widely used as a surrogate marker for angiogenesis in cancer, counting the number of capillaries. The NF-κB expression has been implicated in the regulation of IMVD in human cancer [
16]. This study demonstrated that AIMs significantly inhibited Ki67 proliferation index and IMVD. This finding supports that AIMs harbor significant anti-cancer properties on hepatocellular carcinoma. In addition, at the molecular levels, we found that AIMs inhibited NF-κB activity in both Western blot analysis and immunohistochemical staining. The activation of the NF-κB pathways is mainly involved in the inflammation, cancer cell proliferation, invasion, adhesion, and angiogenesis. Thus, AIMs showed anti-cancer activity through the inhibition of NF-κB activity in hepatocellular carcinoma. We also confirmed that AIMs inhibited NF-κB-regulated proteins involved in cell proliferation and invasion (cyclin D1, COX-2, MMP-2, and MMP-9). Cyclin D1 and COX-2 are overexpressed in a variety of cancers and mediate cancer cell proliferation [
17,
18]. MMP-2 and MMP-9 are key molecules in cancer invasion by the proteolytic digestion of the extracellular matrix (ECM) which is the first step to metastasis [
19,
20] to help cancer cells to migrate into or out of the vessel walls to metastasize to the distant organs.
The limitations of the study are as follows. First, TNF-α is a known NF-κB stimulant. We use it to determine the linkage between NF-κB activation and the anti-cancer effects of AIMs. The pathophysiological relevance of TNF-induced NF-κB activation is highlighted by the reports that TNF-α is very frequently increased in patients with advanced cancers which have high chances to metastasize [
15,
16]. One may question the use of TNF-α, because there was no significant difference between the cells treated with and without TNF-α, especially in cancer cell proliferation. Regarding TNF-α effects on cell proliferation, it should not be overlooked that TNF-α has its own anti-cancer effect, so some cancer cell such as MCF-7 cells are susceptible to TNF-α treatment. However, in many cancer cells, immediately after treatment with TNF-α, NF-κB is activated to prevent cell death, grow better, become more invasive, and show better metastasis. In addition, in the case of advanced or metastatic cancer, TNF-α was increased, so we thought that TNF-α treatment would be more suitable in the in vivo model. As observed in the above results, TNF-α not only increased the growth, invasion, and migration of Hep3B cells, but also reduced the AIM effect, so we wondered how AIMs would work in in vivo systems. Fortunately, however, AIMs showed anti-cancer effects in in vivo systems, by inhibiting NF-κB and its downstream molecules that are involved in cancer proliferation, invasion and metastasis, so it was possible to conclude that AIM also showed anti-cancer effects in vivo. However, it is unfortunate that it was not possible to draw a conclusion about how much the anti-cancer effect of AIMs is reduced in the situation where TNF-α is highly observed in the in vitro study, and whether AIMs will exert the anti-cancer effect in the situation where the actual TNF-α is high, such as highly metastatic cancer or far advanced cancers. Further investigation is warranted to validate the in vitro findings in in vivo systems.
Secondly, AIMs are composed of more various anthocyanins than anthocyanins from other sources [
21,
22]. We could not fully understand the meaning that Meoru has more various anthocyanins than other vegetables or fruit, and which compound is more effective in inhibiting cancer invasion, since few results on the effect of each anthocyanin are available for cancer cells. A previous study reported that the anthocyanidins, mainly cyanidin, delphinidin, and malvidin, caused strong growth inhibition in a hepatoma cell line, whereas the aglycoside anthocyanins, including cyanidin 3-glucoside, peonidin 3-glucoside, pelargonidin 3-glucoside, and malvidin 3-glucoside, had lower inhibitory activities [
23]. AIMs consist of mainly aglycoside anthocyanins. Meoru has been used orally in cancer patients even though it has been used topically for the treatment of wounds. These glycoside chains, when we take these glycosylated anthocyanins orally, would be eliminated to be absorbed in the body. Therefore, more studies are needed regarding the effect of the glycosylation of anthocyanins on anti-cancer activities.
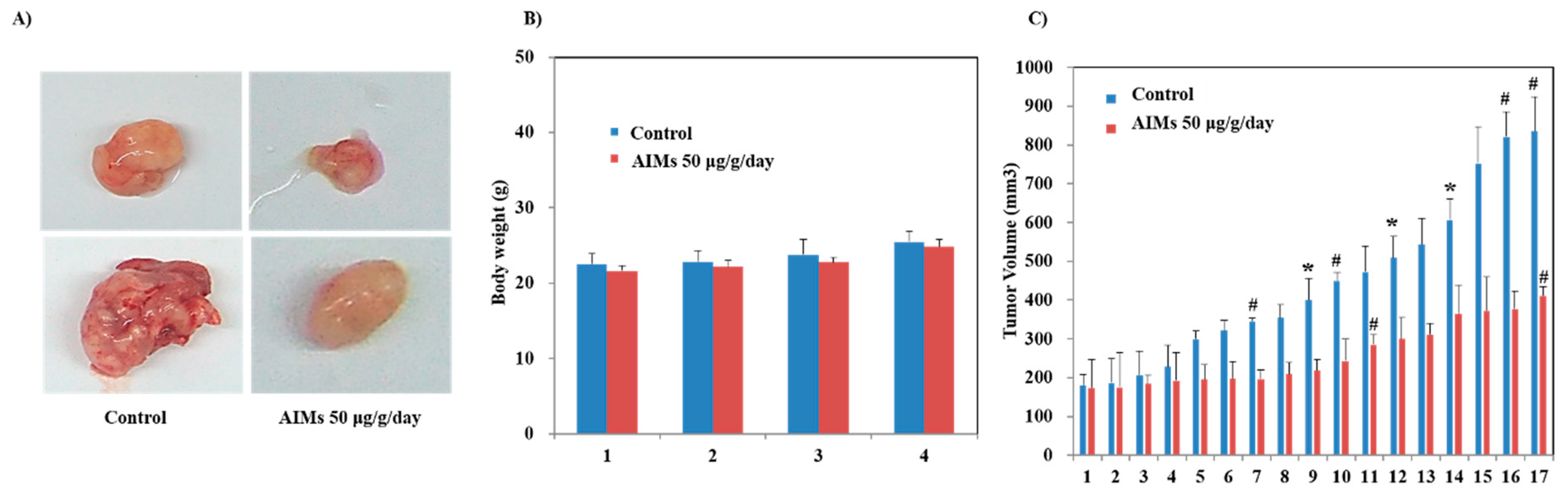
Third, regarding the heterogeneity of the in vivo results, the tumor sizes were various; the largest one in the AIM-treated group was bigger than the smallest one in the control group. Even though the serial measurements of tumors revealed that there were a significant changes in tumor size between the AIM-treated and control group, we were concerned about the bias regarding this experiment. The meticulous examination of tumors by matching the serial exam of the tumor size showed that the AIM effects seemed to appear after 1 month. The tumor size heterogeneity may be explained by seed and soil theory—in the process of settling in different mice, appropriate tumor cells were selected and appeared as tumors of various sizes because the surrounding tumor environment was very important for tumor growth. In addition, we expected that there would be big differences in cancer biology between xenograft tumors. Hence, we were concerned about the Western blot results, but luckily, the expression of proteins that are involved in cancer cell proliferation, invasion, migration, and angiogenesis as well as NF-κB, were suppressed by AIMs.
In conclusion, this study suggests that AIMs have anti-cancer effects in terms of proliferation, migration, and invasion at low concentrations (10–100 μg/mL), on Hep3B human hepatocellular carcinoma cells in vitro and that the anti-cancer effects of AIMs were valid in the xenograft mouse models of Hep3B human hepatocellular carcinoma cells through the inhibition the of NF-κB and its target proteins that are involved in cancer cell proliferation, invasion, and angiogenesis. Although we found that the anti-cancer effects of AIMs were diminished by TNF-α treatment in vitro, we could not draw the conclusion regarding how much the anti-cancer effect of AIMs was reduced or whether AIMs work in the situation where TNF-α is high, frequently observed in highly metastatic cancer or far advanced cancers. Further investigation is warranted to validate the anti-cancer effects of AIMs in TNF-α, high in an in vivo system.
4. Methods
4.1. Cell Culture and Chemicals
Hep3B human hepatocellular carcinoma cells were obtained from the ATCC (American Type Culture Collection) (Rockville, MD, USA). The culture medium used throughout these experiments was RPMI (Roswell Park Memorial Institute Medium) 1640 medium (Invitrogen Corp, Carlsbad, CA, USA) supplemented with 10% FBS (GIBCO BRL, Grand Island, NY, USA), 100 U/mL penicillin, and 100 μg/mL streptomycin in an incubator at 37 °C in a humidified atmosphere of 95% air and 5% CO2. Molecular mass markers for proteins were obtained from Pharmacia Biotech (Saclay, France). For Western blotting and immunohistochemistry, antibodies against COX-2, cyclin D1, XIAP, MMP-2, MMP-9, Bcl-xL, Bcl-2, NF-κB and CD34 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). p-Akt, and p-NF-κB were purchased from Cell signaling Technology, Inc. (Beverly, MA, USA). Antibodies against cyclin D1and Ki-67 were purchased from DAKO. (Santa clara, CA, USA). Antibody against CD34 was purchased from Abcam (Cambridge, MA, USA). Antibody against β-actin was from Sigma (Beverly, MA, USA). Peroxidase-labeled donkey anti-rabbit and sheep anti-mouse immunoglobulin, and an enhanced chemiluminescence (ECL) kit were purchased from Amersham (Arlington Heights, IL, USA). All other chemicals not specifically mentioned were purchased from Sigma Chemical Co. (St. Louis, MO, USA). We used the anthocyanins isolated from Vitis coignetiae Pulliat (AIMs) which consists of: delphinidin-3,5-diglucoside (1): cyanidin-3,5-diglucoside (2): petunidin-3,5-diglucoside (3): delphinidin-3-glucoside (4): malvdin-3,5-diglucoside (5): peonidin-3,5-diglucoside (6): cyanidin-3-glucoside (7): petunidin-3-glucoside (8): peonidin-3-glucoside (9): malvidin-3-glucoside (10) = 3.5: 3.4: 7.1: 23.9: 8.0: 9.6: 9.1: 16.1: 5.7: 13.4.
4.2. Cell Proliferation Assays
For the cell viability assay, the cells (5 × 104 cells/mL) were seeded onto 24-well plates, grown to 70% confluence and then treated with the indicated concentrations of AIMs for 24 and 48 h. Control cells were supplemented with media containing 0.1% DMSO (vehicle control). Cell viability was determined by trypan blue-exclusion methods.
4.3. Wound Healing Assay
Hep3B cells were grown on 35 mm dish plate to 100% confluent monolayer and then scratched to form a 100 µm “wound” using sterile pipette tips. The cells were then cultured in the presence or absence of the indicated concentration of AIMs in serum-free media for 24 h. The images were recorded at 12 and 24 h after scratch using an Olympus photomicroscope.
4.4. Gelatin Zymography
The gelatinolytic activities for MMP-2 and MMP-9 in the culture medium were analyzed by electrophoresis on 10% polyacrylamide gels containing 1 mg/mL gelatin. Polyacrylamide gels were run at 120 V, washed in 2.5% Triton X-100 for 1 h, and then incubated at 37 °C for 16 h in an activation buffer (50 mM Tris-HCl, pH 7.5, 10 mM CaCl2). After staining with Coomassie blue (10% glacial acetic acid, 30% methanol and 1.5% Coomassie brilliant blue) for 2–3 h, the gel was washed with a solution of 10% glacial acetic acid and 30% methanol without Coomassie blue for 1 h. White lysis zones revealed by staining with Coomassie brilliant blue indicated gelatin degradation.
4.5. Western Blotting
This assay was conducted as described previously [
24]. The cells were harvested and lysed in the lysis buffer. The cells were disrupted by sonication and extracted at 4 °C for 30 min. The protein concentrations were quantified using the BioRad protein assay (BioRad Lab., Hercules, CA, USA). The proteins of the extracts were resolved by electrophoresis, electrotransferred to a polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA), and then the membrane was incubated with the primary antibodies followed by a conjugated secondary antibody to peroxidase. Blots were developed with an ECL detection system. ImageJ software 1.51K (
http://rsb.info.nih.gov) was used to quantify each protein band followed by densitometry reading undertaken after normalization with β-Actin expression.
4.6. Animal Experiment
Athymic (nu/nu) male nude mice, obtained from SLC (Sankyo Lab Service, Tokyo, Japan), were housed under pathogen-free conditions with a 12 h light/12 h dark schedule, and fed with an autoclaved diet and water. Hep3B cells (1 × 106 cells/0.1 mL) were subcutaneously injected into the left flank of the mice. Twelve animals were then randomly divided into two groups with six animals each. The first group of animals received an intraperitoneal injection of 100 AL of a 1:10 ratio of DMSO and normal saline, whereas the animals of the second group received an intraperitoneal injection of AIMs (5 μg/g of animal in 100 AL of 1:10 ratio of DMSO and normal saline) daily. One week after the injection of Hep3B xenograft tumor size was measured using a caliper at 2-day intervals, and tumor volume was calculated by the formula 0.5238 × L1 × L2 × H, where L1 is the long diameter, L2 is the short diameter, and H is the height of the tumor. When the mean tumor volume of the non-treatment group crossed the target volume of 1200 mm3, the animals were all sacrificed, and the tumors were removed and frozen in liquid nitrogen for Western blot analysis or fixed with formalin for immunohistochemistry. All animal experiments were approved by the Ethics Committee for Animal Experimentation, Gyeongsang National University.
4.7. Immunohistochemical Staining of NF-κB p65, Cyclin D1, XIAP, and MMP-9 in Tumor Samples
The nuclear localization of NF-κB p65 and the expression of cyclin D1, XIAP, and MMP-9 were examined using an immunohistochemical method described previously [
25]. Briefly, Hep3B tumor xenografts samples were embedded in paraffin and fixed with paraformaldehyde. The staining procedure was achieved by an immunoperoxidase technique on 5 μm-thick consecutive sections. Primary antibodies raised against human NF-κB p65 (Santa Cruz, CA, USA, 1:100 dilution), Cyclin D1 (DAKO, Carpinteria, CA, USA, dilution 1:2000), XIAP (Santa Cruz, CA, USA, 1:50), MMP-9 (Santa Cruz, CA, USA, 1:50), Ki-67 (DAKO, Carpinteria, CA, USA, dilution 1:1000), and CD34 (Abcam, San Jose, CA, USA, dilution 1:1000) were used. Before incubation with primary antibodies, the specimens were pretreated with microwaves for 60 min to retrieve antigenic epitope. The negative controls were parallel sections treated with the same process as the above, but without the treatment of primary antibodies. Positive staining was visualized with diaminobenzidine and hematoxylin was used as counter staining.
4.8. Assessment of Immunohistochemistry
All findings were evaluated by a pathologist blind to the treatment conditions. Samples were evaluated on a percentage of reactive cells demonstrated with moderate or strong staining intensity when the staining intensity was rated as weak, moderate, and strong as compared to the background stain or some negative controls. Images of representative results were pictured. At least 500 tumor cells were evaluated for evaluation in each case.
4.9. Intratumoral Micro Vessel Density (IMVD) Analysis
Frozen sections were fixed in cold acetone and stained with anti-CD34 monoclonal antibody as previously described in the immunohistochemical staining section. The CD34 stained slides were observed under the microscope equipped with a digital camera (CKX41 with a camera, Nikon, DS-U3) and the images were acquired. For each case, IMVD was determined as the maximum number of microvessels under the light microscope within the “hot spot”, which was defined as those areas containing the greatest number of capillaries and small venules at the invasive edge, based on the criteria of Weidner et al. [
26]. The “hot spot” was determined by scanning the tumor sections at low power (×40 and ×100), and the microvessel density was measured with a light microscope in a single area of invasive tumor (×20 objective and ×10 ocular; 0.739 mm
2/field) at the “hot spot” (the representative of the highest microvessel density). Only vessels distinct from one another were counted separately. The evaluations were performed by a pathologist who was blind to the treatment conditions.
4.10. Statistics
The results were expressed as the means ± SD. The comparison of the effects of various treatments was performed with the one-way analysis of variance (ANOVA) with post-test Neuman–Keuls in the cases with at least three treatment groups and the Student’s t test. Statistical significance was defined as p < 0.05. Each experiment was performed at least in triplicate.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}



