
Nucleic Acid Delivery by Solid Lipid Nanoparticles Containing Switchable Lipids: Plasmid DNA vs. Messenger RNA

 ,
,  , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Size, Polydispersity Index and ζ-Potential of SLNs and Vectors
2.2. Transmission Electron Microscopy (TEM) Images
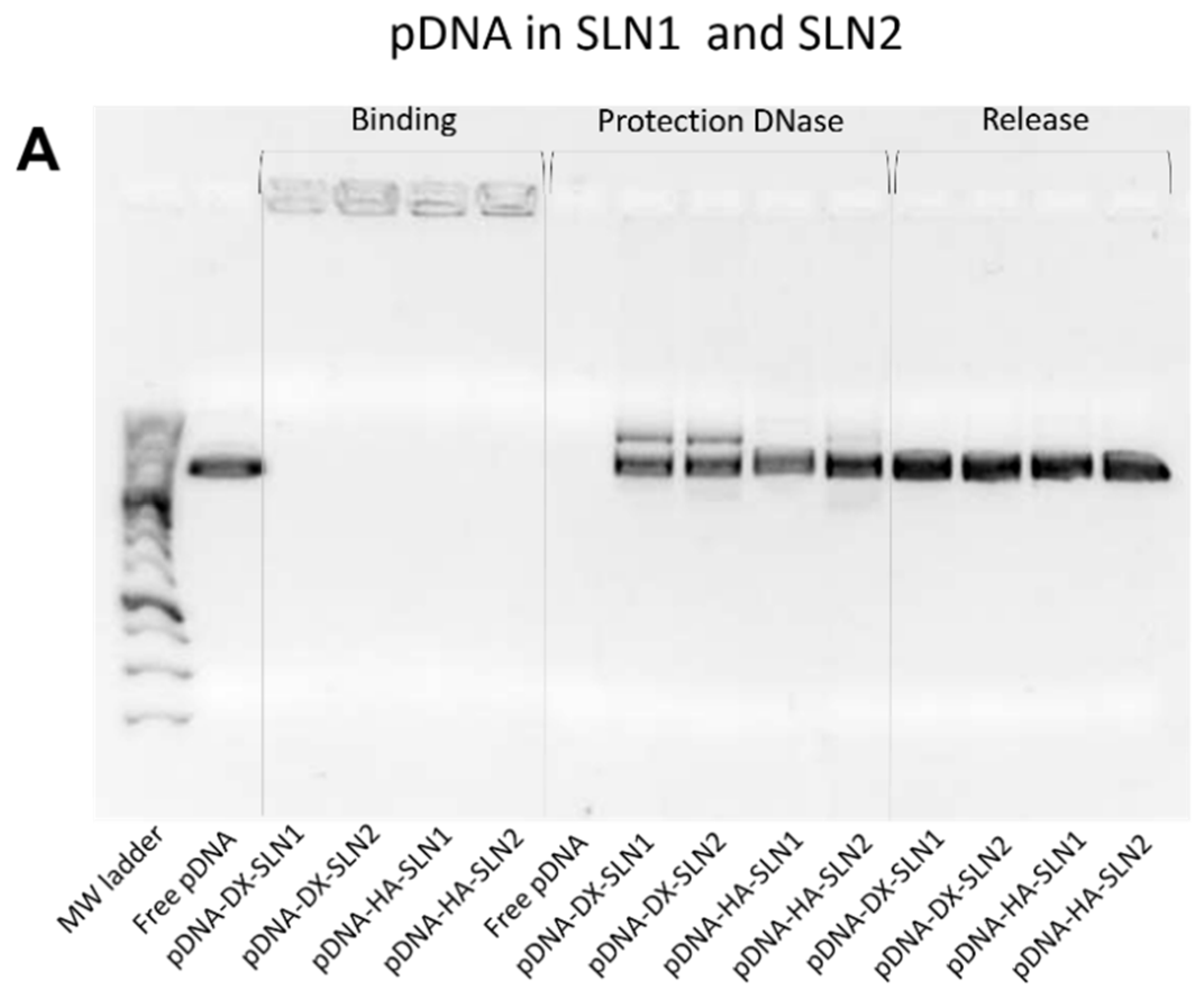
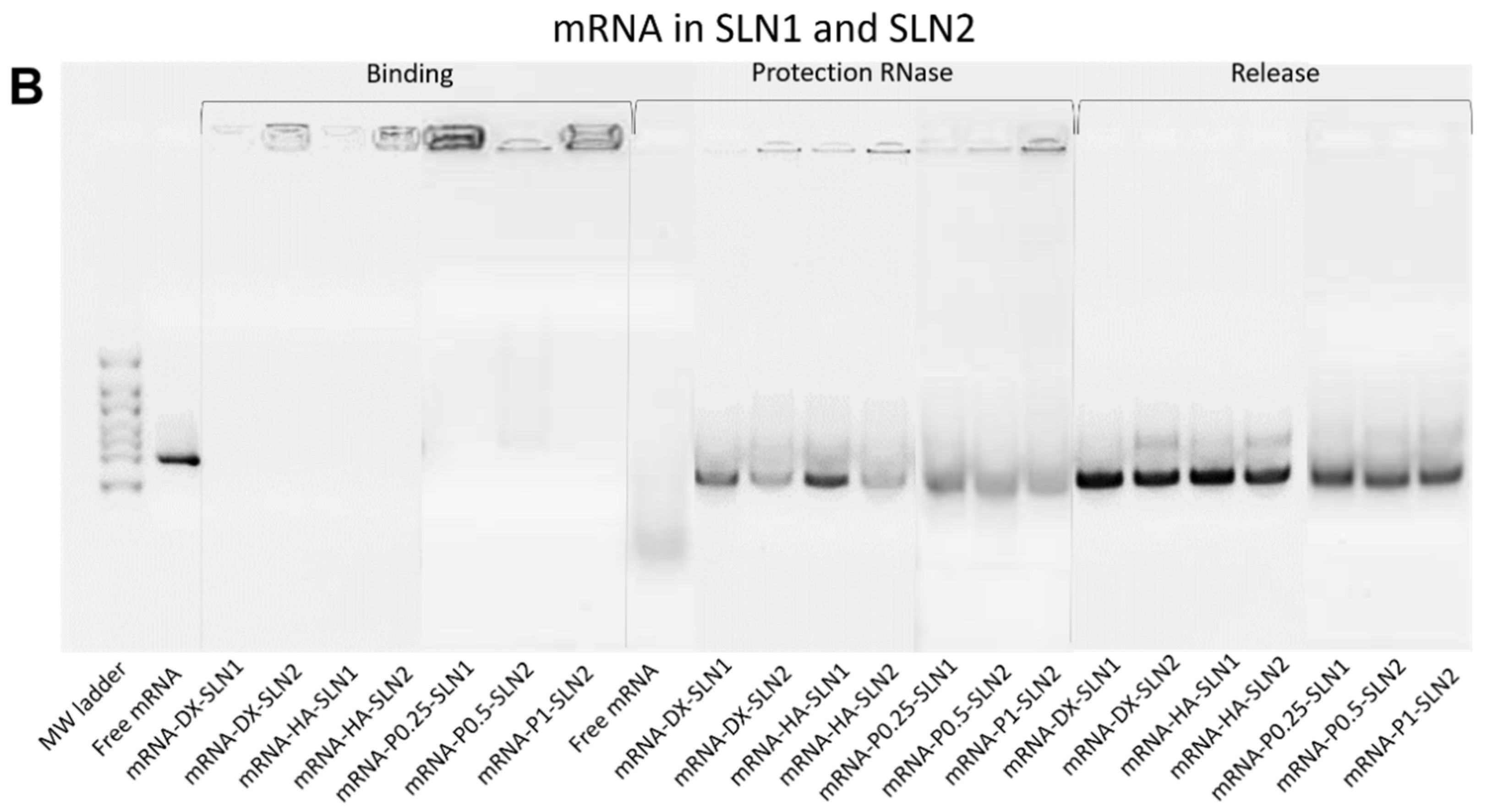
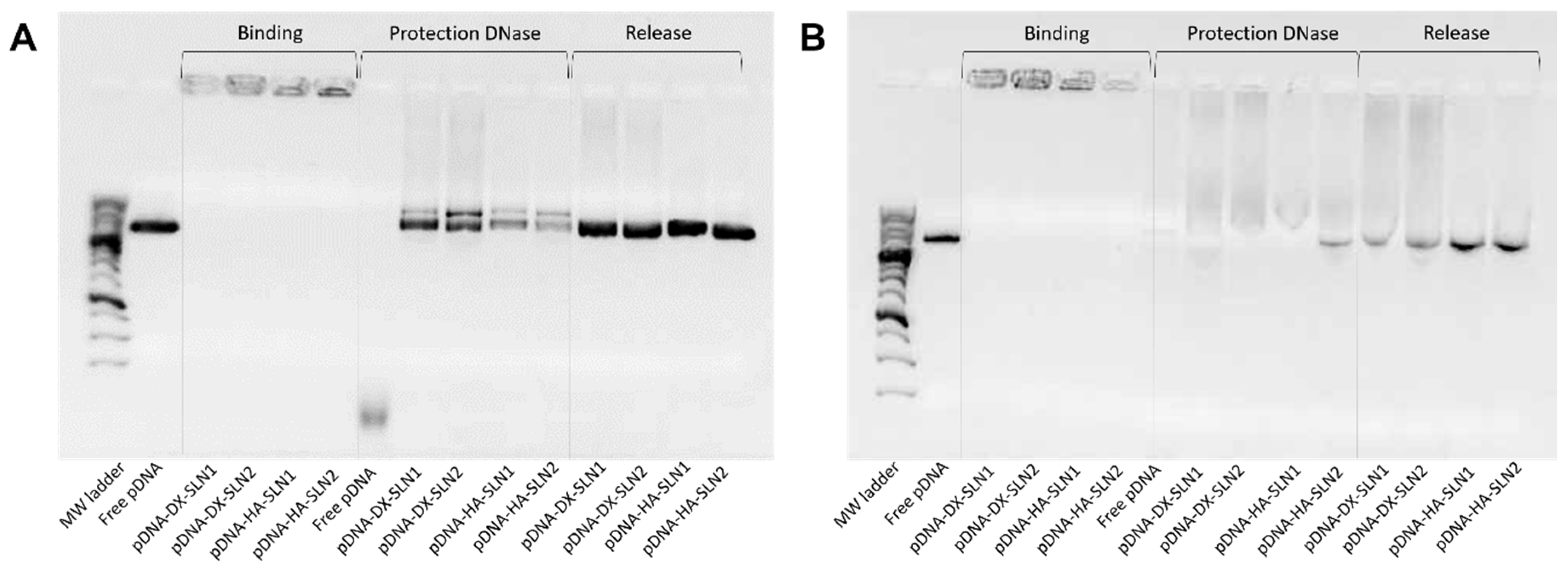
2.3. Binding, Protection and Release of Nucleic Acids
2.4. Cell Culture Studies
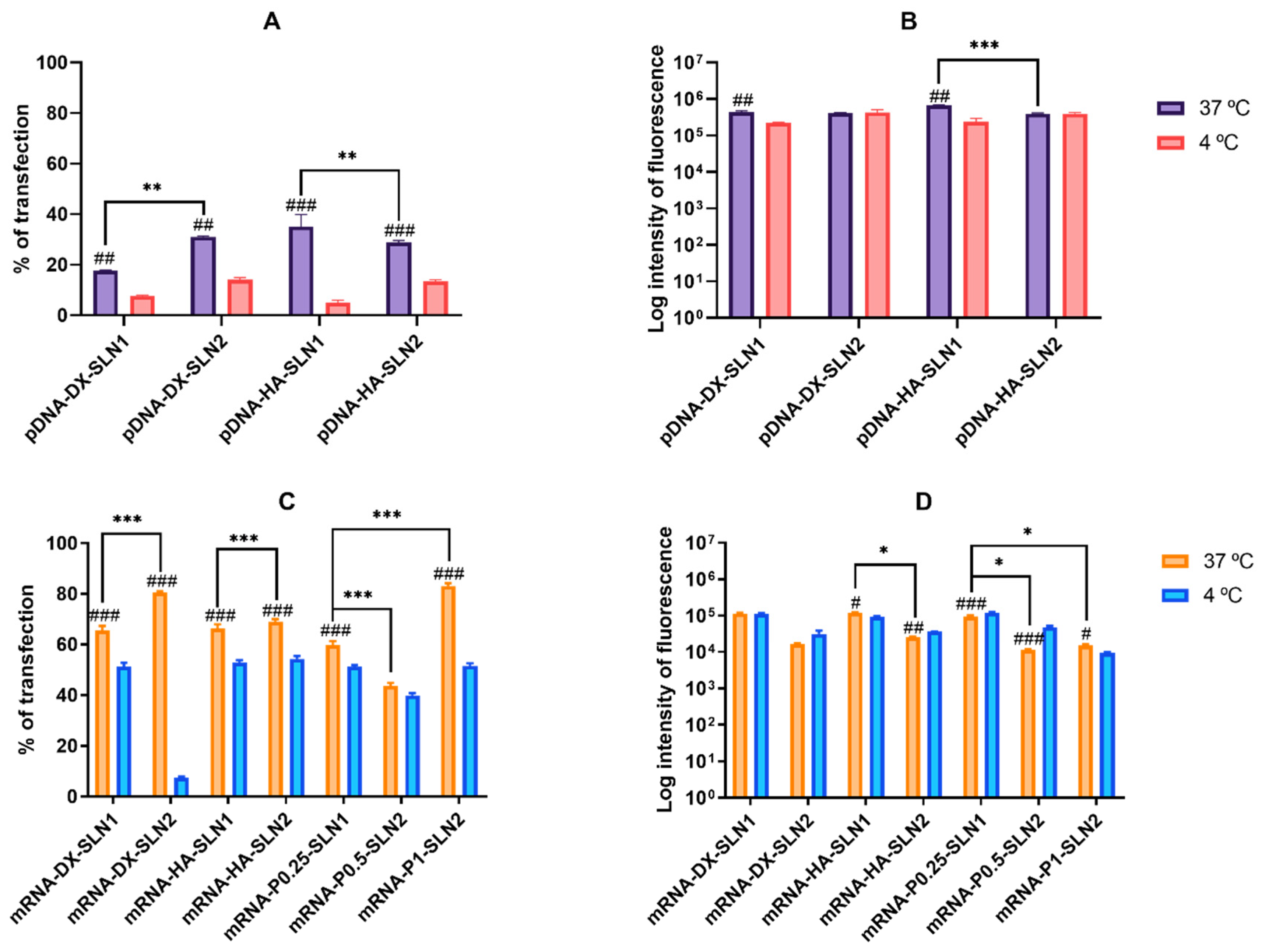
2.4.1. Transfection Efficacy and Cell Viability in ARPE-19 Cells
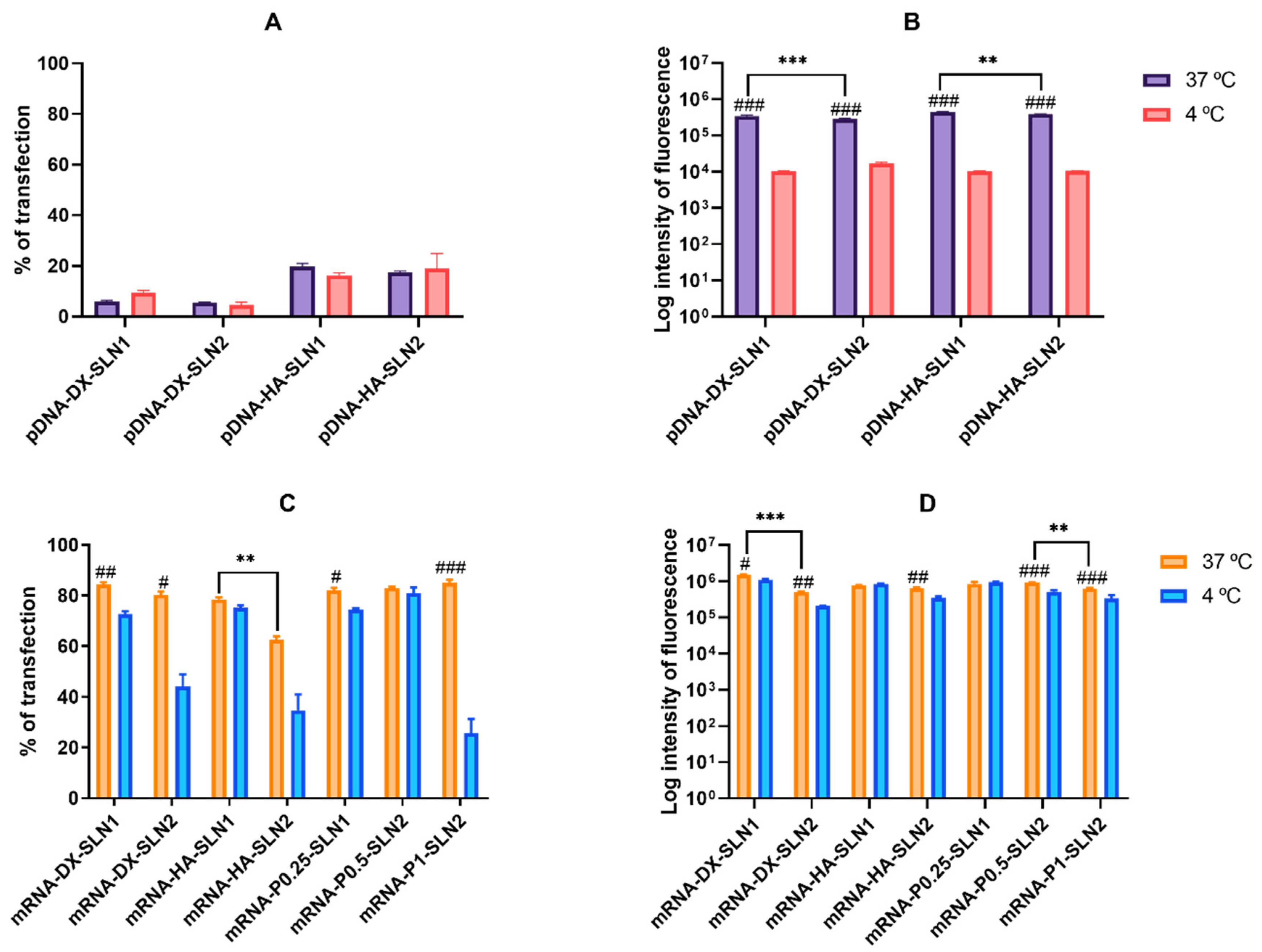
2.4.2. Transfection Efficacy and Cell Viability in HEK-293 Cells
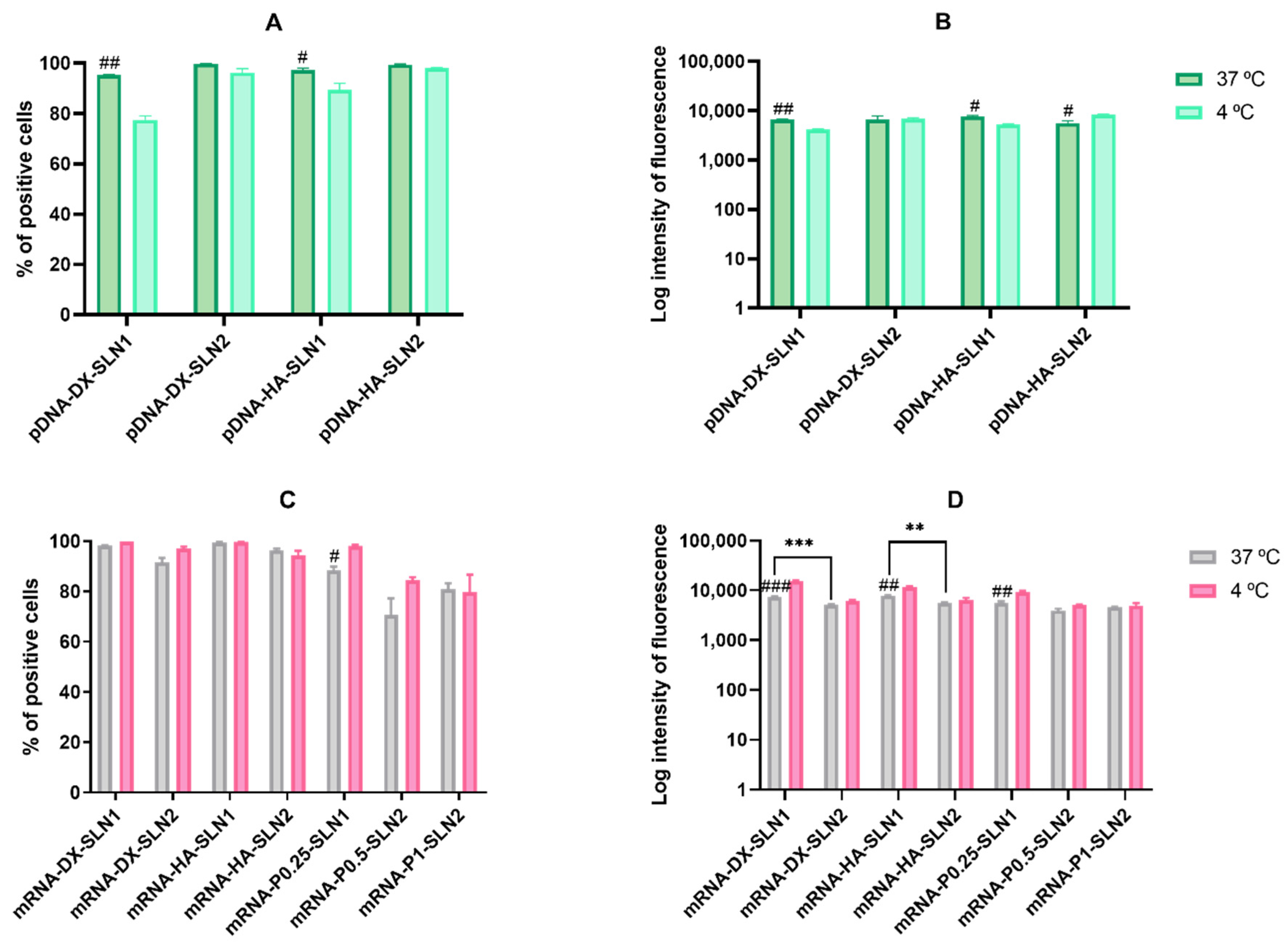
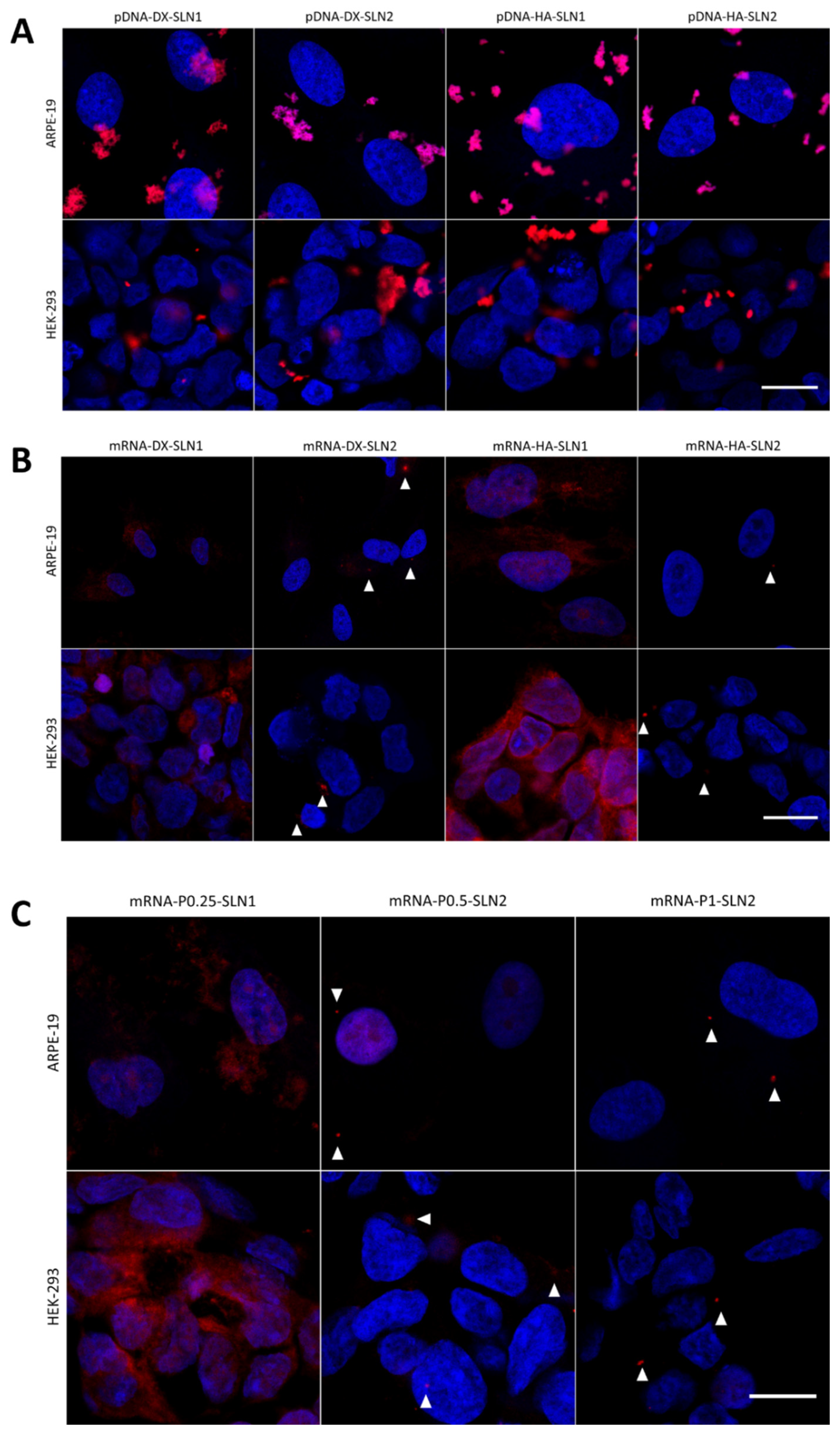
2.4.3. Cellular Uptake in ARPE-19 Cells
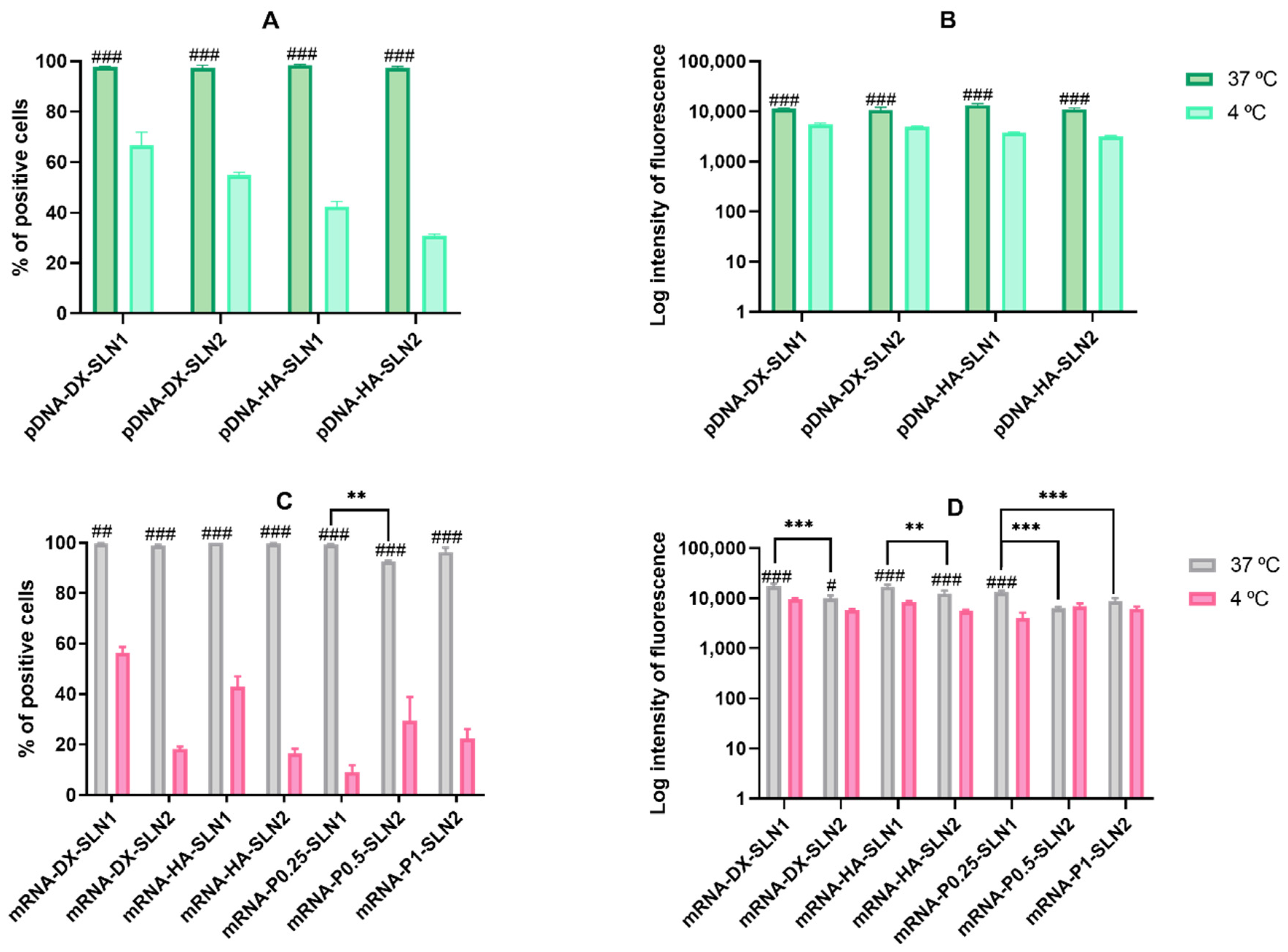
2.4.4. Cellular Uptake of the Vectors in HEK-293 Cells
2.4.5. Intracellular Disposition of Non-Viral Vectors
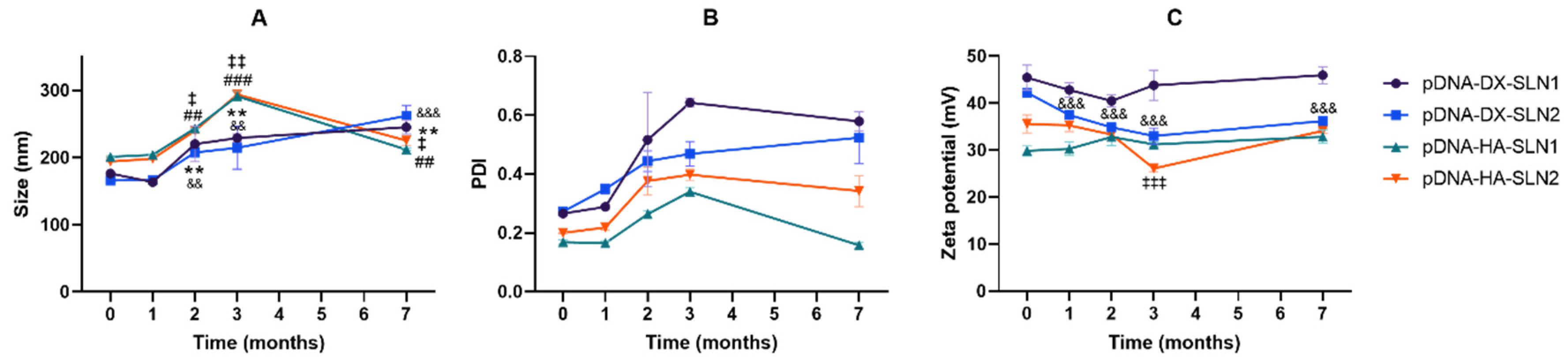
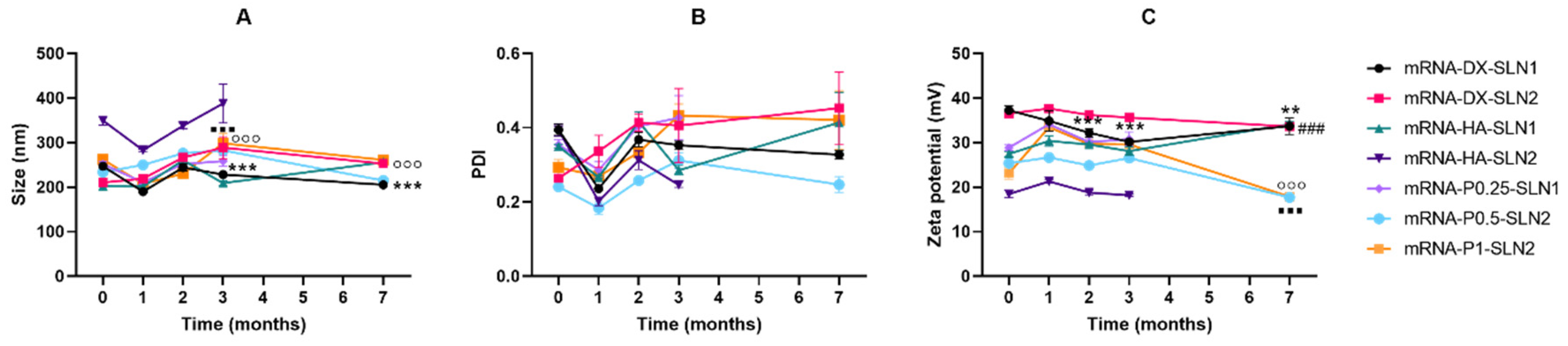
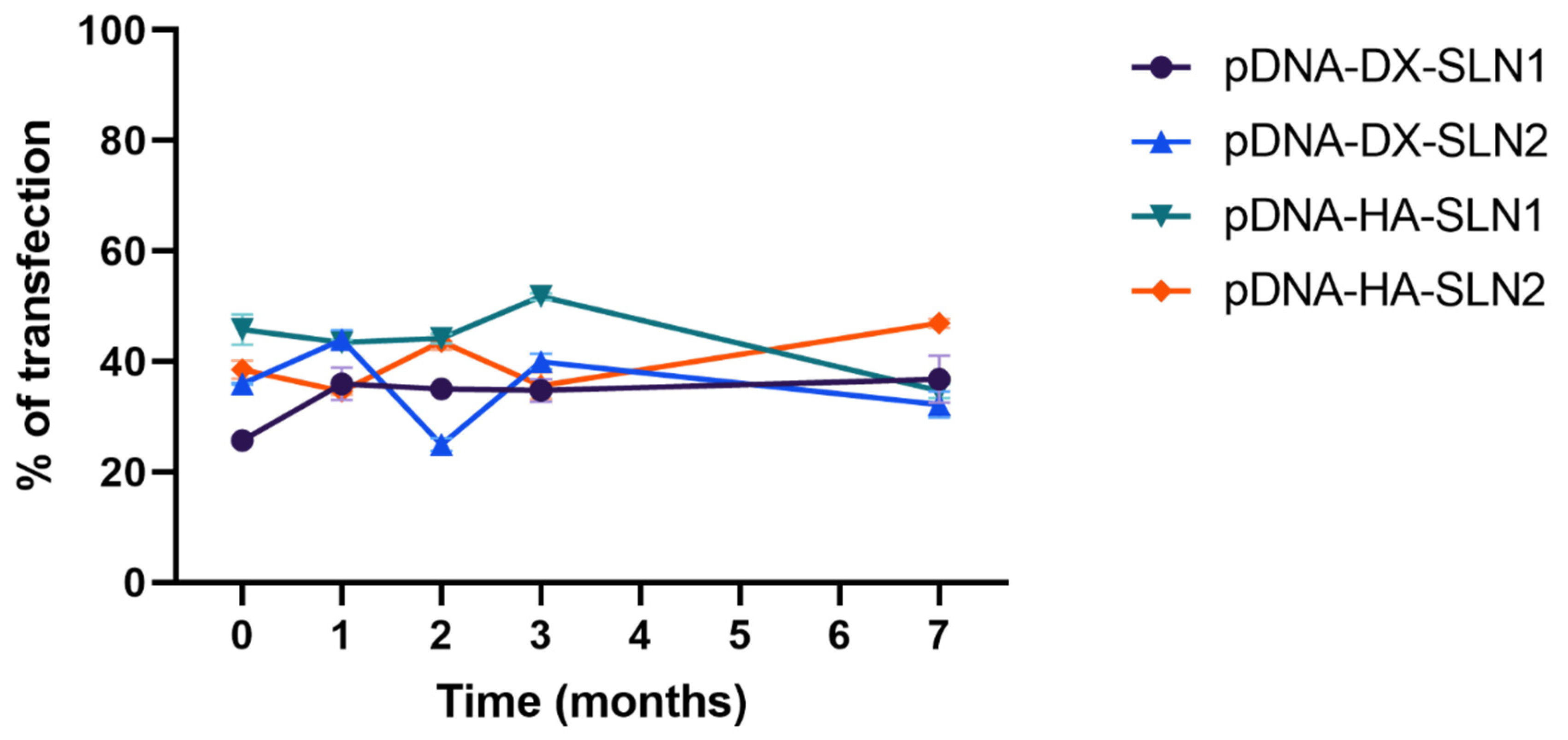
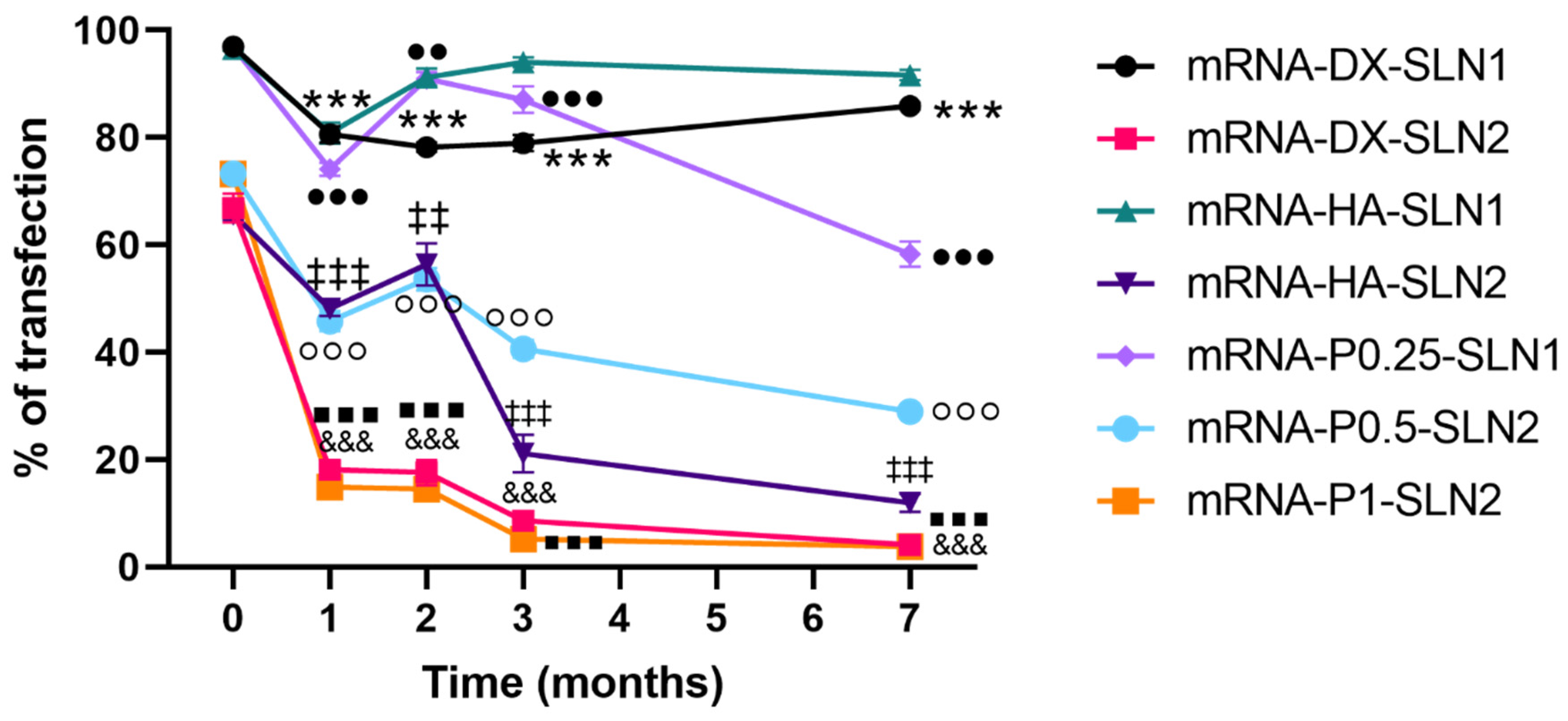
2.5. Long-Term Stability Study of pDNA- and mRNA-Based Vectors
2.5.1. Size, Polydispersity Index and ζ-Potential
2.5.2. Binding, Protection and Release of Nucleic Acids
2.5.3. Transfection Efficacy and Cell Viability
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of SLNs and Vectors
4.3. Size, PDI and ζ-Potential Measurement
4.4. Transmission Electron Microscopy (TEM) Images
4.5. Agarose Gel Electrophoresis Assay
4.6. Cell Culture Studies
4.6.1. Transfection Efficacy and Cell Viability
4.6.2. Cellular Uptake
4.6.3. Intracellular Disposition
4.7. Long-Term Stability Study of pDNA- and mRNA-Based Vectors
Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yu, A.-M.; Choi, Y.H.; Tu, M.-J. RNA Drugs and RNA Targets for Small Molecules: Principles, Progress, and Challenges. Pharmacol. Rev. 2020, 72, 862–898. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines-a new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Mol. Ther. 2019, 27, 710–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Chang, J.; Jiang, Y.; Meng, X.; Sun, T.; Mao, L.; Xu, Q.; Wang, M. Fast and Efficient CRISPR/Cas9 Genome Editing In Vivo Enabled by Bioreducible Lipid and Messenger RNA Nanoparticles. Adv. Mater. 2019, 31, 1902575. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.B.; Zhang, S.; Kos, P.; Xiong, H.; Zhou, K.; Perelman, S.S.; Zhu, H.; Siegwart, D.J. Non-Viral CRISPR/Cas Gene Editing In Vitro and In Vivo Enabled by Synthetic Nanoparticle Co-Delivery of Cas9 mRNA and sgRNA. Angew. Chem. Int. Ed. 2017, 56, 1059–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Aguado, I.; Rodríguez-Castejón, J.; Vicente-Pascual, M.; Rodríguez-Gascón, A.; Solinís, M.Á.; Del Pozo-Rodríguez, A. Nanomedicines to deliver mRNA: State of the art and future perspectives. Nanomaterials 2020, 10, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, U.; Karikó, K.; Türeci, Ö. MRNA-based therapeutics-developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Zhong, Z.; Mc Cafferty, S.; Combes, F.; Huysmans, H.; De Temmerman, J.; Gitsels, A.; Vanrompay, D.; Portela Catani, J.; Sanders, N.N. mRNA therapeutics deliver a hopeful message. Nano Today 2018, 23, 16–39. [Google Scholar] [CrossRef]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef]
- Papachristofilou, A.; Hipp, M.M.; Klinkhardt, U.; Früh, M.; Sebastian, M.; Weiss, C.; Pless, M.; Cathomas, R.; Hilbe, W.; Pall, G.; et al. Phase Ib evaluation of a self-adjuvanted protamine formulated mRNA-based active cancer immunotherapy, BI1361849 (CV9202), combined with local radiation treatment in patients with stage IV non-small cell lung cancer. J. Immunother. Cancer 2019, 7, 38. [Google Scholar] [CrossRef]
- Sebastian, M.; Papachristofilou, A.; Weiss, C.; Früh, M.; Cathomas, R.; Hilbe, W.; Wehler, T.; Rippin, G.; Koch, S.D.; Scheel, B.; et al. Phase Ib study evaluating a self-adjuvanted mRNA cancer vaccine (RNActive®) combined with local radiation as consolidation and maintenance treatment for patients with stage IV non-small cell lung cancer. BMC Cancer 2014, 14, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahl, K.; Senn, J.J.; Yuzhakov, O.; Bulychev, A.; Brito, L.A.; Hassett, K.J.; Laska, M.E.; Smith, M.; Almarsson, Ö.; Thompson, J.; et al. Preclinical and Clinical Demonstration of Immunogenicity by mRNA Vaccines against H10N8 and H7N9 Influenza Viruses. Mol. Ther. 2017, 25, 1316–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaridou, E.; Heyes, J.; Lutwyche, P. Lipid nanoparticles for nucleic acid delivery: Current perspectives. Adv. Drug Deliv. Rev. 2020, 154–155, 37–63. [Google Scholar] [CrossRef] [PubMed]
- del Pozo-Rodríguez, A.; Solinís, M.Á.; Rodríguez-Gascón, A. Applications of lipid nanoparticles in gene therapy. Eur. J. Pharm. Biopharm. 2016, 109, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Apaolaza, P.S.; del Pozo-Rodríguez, A.; Torrecilla, J.; Rodríguez-Gascón, A.; Rodríguez, J.M.; Friedrich, U.; Weber, B.H.F.; Solinís, M.A. Solid lipid nanoparticle-based vectors intended for the treatment of X-linked juvenile retinoschisis by gene therapy: In vivo approaches in Rs1h-deficient mouse model. J. Control. Release 2015, 217, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Apaolaza, P.S.; del Pozo-Rodríguez, A.; Solinís, M.A.; Rodríguez, J.M.; Friedrich, U.; Torrecilla, J.; Weber, B.H.F.; Rodríguez-Gascón, A. Structural recovery of the retina in a retinoschisin-deficient mouse after gene replacement therapy by solid lipid nanoparticles. Biomaterials 2016, 90, 40–49. [Google Scholar] [CrossRef]
- Ramamoorth, M.; Narvekar, A. Non viral vectors in gene therapy—An overview. J. Clin. Diagn. Res. 2015, 9, GE01–GE06. [Google Scholar] [CrossRef]
- Paliwal, R.; Paliwal, S.R.; Kenwat, R.; Das Kurmi, B.; Sahu, M.K. Solid lipid nanoparticles: A review on recent perspectives and patents. Expert Opin. Ther. Pat. 2020, 30, 179–194. [Google Scholar] [CrossRef]
- Guan, S.; Rosenecker, J. Nanotechnologies in delivery of mRNA therapeutics using nonviral vector-based delivery systems. Gene Ther. 2017, 24, 133–143. [Google Scholar] [CrossRef]
- Balazs, D.A.; Godbey, W. Liposomes for Use in Gene Delivery. J. Drug Deliv. 2011, 2011, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ramezanpour, M.; Schmidt, M.L.; Bodnariuc, I.; Kulkarni, J.A.; Leung, S.S.W.; Cullis, P.R.; Thewalt, J.L.; Tieleman, D.P. Ionizable amino lipid interactions with POPC: Implications for lipid nanoparticle function. Nanoscale 2019, 11, 14141–14146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayour, E.J.; De Leon, G.; Pham, C.; Grippin, A.; Kemeny, H.; Chua, J.; Huang, J.; Sampson, J.H.; Perez, L.S.; Flores, C.; et al. Systemic activation of antigen-presenting cells via RNA-Loaded nanoparticles. Oncoimmunology 2017, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siewert, C.D.; Haas, H.; Cornet, V.; Nogueira, S.S.; Nawroth, T.; Uebbing, L.; Ziller, A.; Al-Gousous, J.; Radulescu, A.; Schroer, M.A.; et al. Hybrid Biopolymer and Lipid Nanoparticles with Improved Transfection Efficacy for mRNA. Cells 2020, 9, 2034. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Fotin-Mleczek, M.; Roth, G.; Becker, C.; Dam, T.C.; Verdurmen, W.P.R.; Brock, R.; Probst, J.; Schlake, T. Protein expression from exogenous mRNA: Uptake by receptor-mediated endocytosis and trafficking via the lysosomal pathway. RNA Biol. 2011, 8, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Delgado, D.; Gascón, A.R.; Del Pozo-Rodríguez, A.; Echevarría, E.; Ruiz De Garibay, A.P.; Rodríguez, J.M.; Solinís, M.Á. Dextran-protamine-solid lipid nanoparticles as a non-viral vector for gene therapy: In vitro characterization and in vivo transfection after intravenous administration to mice. Int. J. Pharm. 2012, 425, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Delgado, D.; Del Pozo-Rodríguez, A.; Solinís, M.Á.; Rodríguez-Gascón, A. Understanding the mechanism of protamine in solid lipid nanoparticle-based lipofection: The importance of the entry pathway. Eur. J. Pharm. Biopharm. 2011, 79, 495–502. [Google Scholar] [CrossRef]
- Uchida, S.; Kataoka, K. Design concepts of polyplex micelles for in vivo therapeutic delivery of plasmid DNA and messenger RNA. J. Biomed. Mater. Res. Part A 2019, 107, 978–990. [Google Scholar] [CrossRef]
- Fishman, S.; Lewis, M.D.; Siew, L.K.; De Leenheer, E.; Kakabadse, D.; Davies, J.; Ziv, D.; Margalit, A.; Karin, N.; Gross, G.; et al. Adoptive Transfer of mRNA-Transfected T Cells Redirected against Diabetogenic CD8 T Cells Can Prevent Diabetes. Mol. Ther. 2017, 25, 456–464. [Google Scholar] [CrossRef]
- Van Hoecke, L.; Roose, K. How mRNA therapeutics are entering the monoclonal antibody field. J. Transl. Med. 2019, 17, 54. [Google Scholar] [CrossRef] [Green Version]
- Apaolaza, P.S.; Delgado, D.; Pozo-Rodríguez, A.; Gascón, A.R.; Solinís, M.Á. A novel gene therapy vector based on hyaluronic acid and solid lipid nanoparticles for ocular diseases. Int. J. Pharm. 2014, 465, 413–426. [Google Scholar] [CrossRef]
- Delgado, D.; Del Pozo-Rodríguez, A.; Solinís, M.Á.; Avilés-Triqueros, M.; Weber, B.H.F.; Fernández, E.; Gascón, A.R. Dextran and protamine-based solid lipid nanoparticles as potential vectors for the treatment of X-linked juvenile retinoschisis. Hum. Gene Ther. 2011, 23, 345–355. [Google Scholar] [CrossRef]
- Li, J.; Chen, Q.; Zha, Z.; Li, H.; Toh, K.; Dirisala, A.; Matsumoto, Y.; Osada, K.; Kataoka, K.; Ge, Z. Ternary polyplex micelles with PEG shells and intermediate barrier to complexed DNA cores for efficient systemic gene delivery. J. Control. Release 2015, 209, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Hasanzadeh Davarani, F.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M.R. Impact of particle size and polydispersity index on the clinical applications of lipidic nanocarrier systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resina, S.; Prevot, P.; Tjierry, A.R. Physico-chemical characteristics of lipoplexes influence cell uptake mechanisms and transfection efficacy. PLoS ONE 2009, 4, e6058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, H.; Inoue, K.; Imamura, K.; Yuvienco, C.; Montclare, J.K.; Yamano, S. Efficient siRNA delivery and gene silencing using a lipopolypeptide hybrid vector mediated by a caveolae-mediated and temperature-dependent endocytic pathway. J. Nanobiotechnol. 2019, 17, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Pozo-Rodríguez, A.; Pujals, S.; Delgado, D.; Solinís, M.A.; Gascón, A.R.; Giralt, E.; Pedraz, J.L. A proline-rich peptide improves cell transfection of solid lipid nanoparticle-based non-viral vectors. J. Control. Release 2009, 133, 52–59. [Google Scholar] [CrossRef]
- Yamada, Y.; Sato, Y.; Nakamura, T.; Harashima, H. Evolution of drug delivery system from viewpoint of controlled intracellular trafficking and selective tissue targeting toward future nanomedicine. J. Control. Release 2020, 327, 533–545. [Google Scholar] [CrossRef]
- Patel, S.; Ashwanikumar, N.; Robinson, E.; Duross, A.; Sun, C.; Murphy-Benenato, K.E.; Mihai, C.; Almarsson, Ö.; Sahay, G. Boosting Intracellular Delivery of Lipid Nanoparticle-Encapsulated mRNA. Nano Lett. 2017, 17, 5711–5718. [Google Scholar] [CrossRef]
- Uchida, H.; Itaka, K.; Nomoto, T.; Ishii, T.; Suma, T.; Ikegami, M.; Miyata, K.; Oba, M.; Nishiyama, N.; Kataoka, K. Modulated protonation of side chain aminoethylene repeats in N-substituted polyaspartamides promotes mRNA transfection. J. Am. Chem. Soc. 2014, 136, 12396–12405. [Google Scholar] [CrossRef] [Green Version]
- Maruggi, G.; Zhang, C.; Li, J.; Ulmer, J.B.; Yu, D. mRNA as a Transformative Technology for Vaccine Development to Control Infectious Diseases. Mol. Ther. 2019, 27, 757–772. [Google Scholar] [CrossRef] [Green Version]
- del Pozo-Rodríguez, A.; Delgado, D.; Solinís, M.A.; Gascón, A.R.; Pedraz, J.L. Solid lipid nanoparticles: Formulation factors affecting cell transfection capacity. Int. J. Pharm. 2007, 339, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Pascual, M.; Albano, A.; Solinís, M.; Serpe, L.; Rodríguez-Gascón, A.; Foglietta, F.; Muntoni, E.; Torrecilla, J.; Del Pozo-Rodríguez, A.; Battaglia, L. Gene delivery in the cornea: In vitro & ex vivo evaluation of solid lipid nanoparticle-based vectors. Nanomedicine 2018, 13, 1847–1864. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SLNs | Cationic/Ionizable Lipid | Size (nm) | PDI | ζ-Potential (mV) |
|---|---|---|---|---|
| SLN1 | DOTAP/- | 185.1 ± 3.5 | 0.30 ± 0.03 | +59.5 ± 1.9 & |
| SLN2 | DOTAP/DODAP | 208.8 ± 0.4 | 0.27 ± 0.01 | +50.2 ± 1.1 & |
| SLN3 | -/DOBAQ | 423.5 ± 51.7 *** | 0.57 ± 0.05 | −28.0 ± 0.8 & |
| SLN4 | DOTAP/DOBAQ | 211.3 ± 3.5 | 0.36 ± 0.01 | +42.4 ± 1.2 & |
| Nucleic Acid | Vectors | Size (nm) | PDI | ζ-Potential (mV) |
|---|---|---|---|---|
| pcDNA3-EGFP plasmid | pDNA-DX-SLN1 | 176.4 ± 0.4 | 0.27 ± 0.01 | +45.4 ± 2.7 |
| pDNA-DX-SLN2 | 165.8 ± 1.7 | 0.27 ± 0.01 | +42.2 ± 0.9 | |
| pDNA-DX-SLN4 | 211.9 ± 14.6 * | 0.40 ± 0.07 ### | +32.6 ± 0.9 | |
| pDNA-HA-SLN1 | 201.2 ± 1.3 | 0.17 ± 0.01 | +29.8 ± 1.1 &&& | |
| pDNA-HA-SLN2 | 194.2 ± 0.8 | 0.20 ± 0.00 | +35.6 ± 1.9 & | |
| CleanCap™ EGFP mRNA (5moU) | mRNA-DX-SLN1 | 246.8 ± 1.3 | 0.39 ± 0.02 | +37.2 ± 1.0 |
| mRNA-DX-SLN2 | 210.1 ± 0.8 ** | 0.26 ± 0.01 | +36.5 ± 0.3 | |
| mRNA-HA-SLN1 | 202.4 ± 2.2 ** | 0.35 ± 0.00 | +27.5 ± 0.6 | |
| mRNA-HA-SLN2 | 349.2 ± 9.9 ●●● | 0.39 ± 0.02 | +18.5 ± 0.9 ● | |
| mRNA-P0.25-SLN1 | 251.6 ± 6.5 | 0.35 ± 0.01 | +28.8 ± 0.7 | |
| mRNA-P0.5-SLN2 | 233.7 ± 2.8 | 0.24 ± 0.00 | +25.4 ± 0.6 | |
| mRNA-P1-SLN2 | 261.7 ± 4.0 | 0.29 ± 0.02 | +23.1 ± 1.3 |
| Type of SLN | Cationic Lipid (%) | Tween 80 (%) | ||
|---|---|---|---|---|
| DOTAP | DODAP | DOBAQ | ||
| SLN1 | 0.4 | 0.1 | ||
| SLN2 | 0.2 | 0.2 | 0.1 | |
| SLN3 | 0.4 | 0.1 | ||
| SLN4 | 0.2 | 0.2 | 0.1 | |
| Name of the Vector | Nucleic Acid | Polysaccharide | SLN | Weight Ratio (w:w:w:w) |
|---|---|---|---|---|
| pDNA-DX-SLN1 | pDNA | DX | SLN1 | DX:P:DNA:SLN1 1:2:1:5 |
| pDNA-DX-SLN2 | pDNA | DX | SLN2 | DX:P:DNA:SLN2 1:2:1:5 |
| pDNA-DX-SLN4 | pDNA | DX | SLN4 | DX:P:DNA:SLN4 1:2:1:5 |
| pDNA-HA-SLN1 | pDNA | HA | SLN1 | HA:P:DNA:SLN1 0.5:2:1:2 |
| pDNA-HA-SLN2 | pDNA | HA | SLN2 | HA:P:DNA:SLN2 0.5:2:1:5 |
| mRNA-DX-SLN1 | mRNA | DX | SLN1 | DX:P:mRNA:SLN1 1:0.25:1:5 |
| mRNA-DX-SLN2 | mRNA | DX | SLN2 | DX:P:mRNA:SLN2 1:1:1:5 |
| mRNA-HA-SLN1 | mRNA | HA | SLN1 | HA:P:mRNA:SLN1 0.5:0.5:1:5 |
| mRNA-HA-SLN2 | mRNA | HA | SLN2 | HA:P:mRNA:SLN2 0.5:1:1:5 |
| mRNA-P-SLN1 | mRNA | - | SLN1 | P:mRNA:SLN1 0.25:1:5 |
| mRNA-P0.5-SLN2 | mRNA | - | SLN2 | P:mRNA:SLN2 0.5:1:5 |
| mRNA-P1-SLN2 | mRNA | - | SLN2 | P:mRNA:SLN2 1:1:5 |
Sample Availability: Samples of the compounds are not available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Aguado, I.; Rodríguez-Castejón, J.; Vicente-Pascual, M.; Rodríguez-Gascón, A.; del Pozo-Rodríguez, A.; Solinís Aspiazu, M.Á. Nucleic Acid Delivery by Solid Lipid Nanoparticles Containing Switchable Lipids: Plasmid DNA vs. Messenger RNA. Molecules 2020, 25, 5995. https://doi.org/10.3390/molecules25245995
Gómez-Aguado I, Rodríguez-Castejón J, Vicente-Pascual M, Rodríguez-Gascón A, del Pozo-Rodríguez A, Solinís Aspiazu MÁ. Nucleic Acid Delivery by Solid Lipid Nanoparticles Containing Switchable Lipids: Plasmid DNA vs. Messenger RNA. Molecules. 2020; 25(24):5995. https://doi.org/10.3390/molecules25245995
Chicago/Turabian StyleGómez-Aguado, Itziar, Julen Rodríguez-Castejón, Mónica Vicente-Pascual, Alicia Rodríguez-Gascón, Ana del Pozo-Rodríguez, and María Ángeles Solinís Aspiazu. 2020. "Nucleic Acid Delivery by Solid Lipid Nanoparticles Containing Switchable Lipids: Plasmid DNA vs. Messenger RNA" Molecules 25, no. 24: 5995. https://doi.org/10.3390/molecules25245995
APA StyleGómez-Aguado, I., Rodríguez-Castejón, J., Vicente-Pascual, M., Rodríguez-Gascón, A., del Pozo-Rodríguez, A., & Solinís Aspiazu, M. Á. (2020). Nucleic Acid Delivery by Solid Lipid Nanoparticles Containing Switchable Lipids: Plasmid DNA vs. Messenger RNA. Molecules, 25(24), 5995. https://doi.org/10.3390/molecules25245995