
Unbalanced Sphingolipid Metabolism and Its Implications for the Pathogenesis of Psoriasis



{kind=link}
{kind=link}
{kind=link}
Abstract
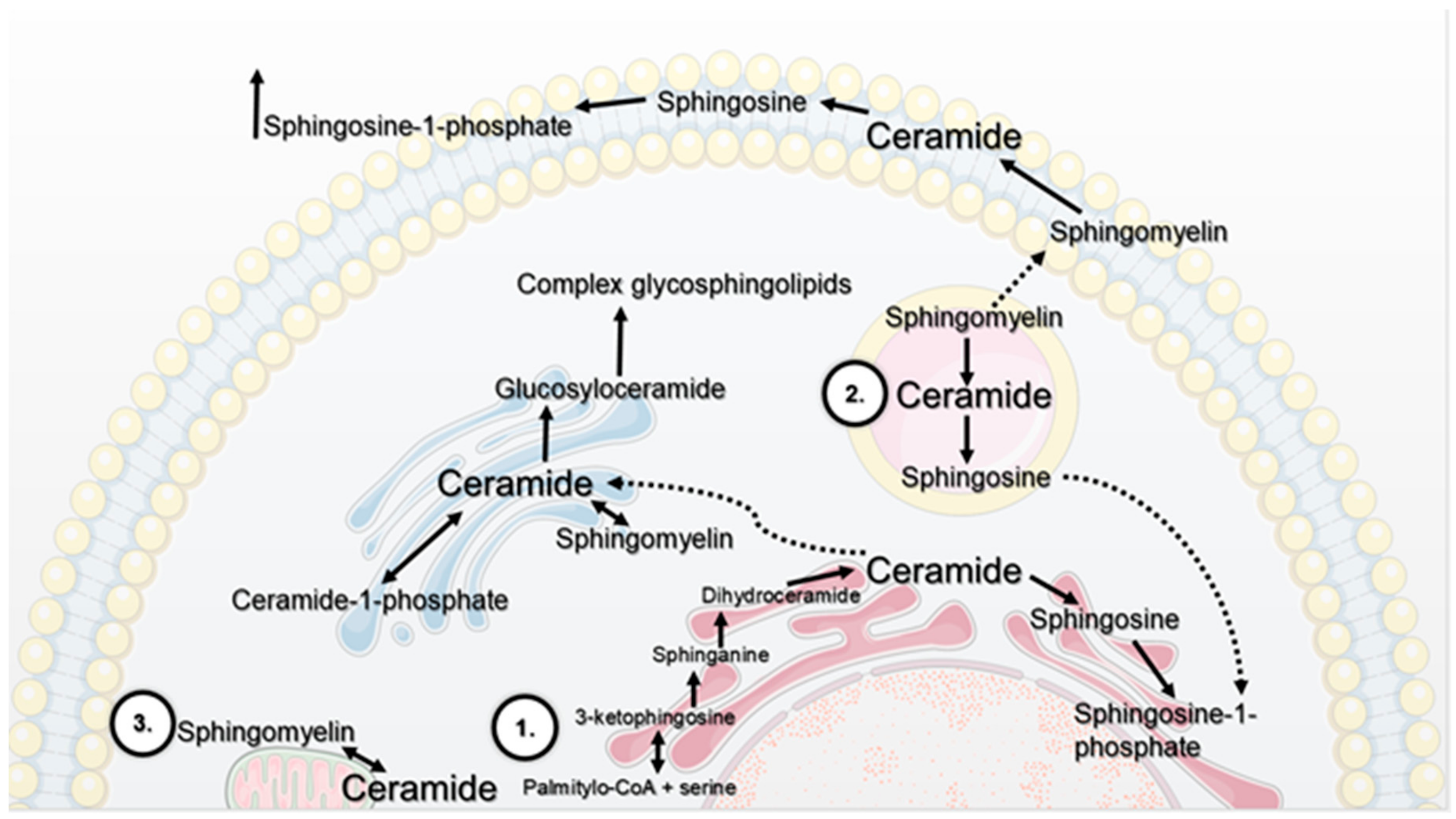
:1. The Contribution of Selected Cellular Organelles In Lipid Metabolism
- de novo synthesis pathway on the cytoplasmic side of the endoplasmatic reticulum. The first stage is the synthesis of 3-ketosphinganine, which consists of the condensation of L-serine and palmityl-CoA catalysed by the enzyme serine palmitoyltransferase. It is a key enzyme in the regulation of the synthesis of 3-ketosphinganine. In the next stage, this compound is reduced to sphinganine, which is acylated to dihydroceramide by the corresponding synthases. The last stage of the synthesis is the oxidation of dihydroceramide to ceramide catalysed by dihydroceramide desaturase [2,3,4].
- salvage pathway in lysosomes. Complex glycosphingolipids, which are important for the construction of biological membranes, undergo cyclic catabolism. Released Cer, with the participation of acid ceramidase, is converted into Sph. Studies show that Sph leaves the lysosome and can be metabolised to Cer or phosphorylated to sphingosine-1-phosphate (S1P). Kitatani et al. indicate that this pathway is responsible for 50–90% of SL biosynthesis [5].
- catabolic pathway on the cellular membrane. As a result of hydrolysis of sphingomyelin (SM) in the presence of the corresponding enzymes, sphingomyelinases (SMases), Cer and phosphocholine are formed. External factors, such as treatment with tumour necrosis factor α (TNF-α) or oxidative stress [5], cause hydrolysis of SM due to activation of sphingomyelinase. The classification of this enzyme is based on the differences in optimum pH values of the catalysed reaction and subcellular distribution; therefore, it stands out as acid SMase (aSMase; encoded by ASAH1); neutral SMases (N-SMases; encoded by ASAH2, ASAH2B and ASAH2C) and alkaline SMase (alk-SMase; encoded by ENPP7). Acid sphingomyelinase is located inside the cell and has the highest activity at pH 4.0–4.5, while the neutral one is placed on the outside of the plasma membrane and has optimal activity at pH 7.4. Moreover, it requires the presence of magnesium (Mg2+) or manganese (Mn2+) ions as an activator. SM degradation through activity of acid and neutral sphingomyelinase lasts a few seconds compared to de novo synthesis of Cer, whose duration is at least several hours [5,6].
2. Sphingolipids As A Structural Molecules - Role In Skin Barrier
Skin Barrier Dysfunction in Psoriasis
3. Sphingolipids As Signaling Molecules
3.1. Sphingolipid Role in Cellular Processes – Deregulation Of Keratinocytes’ Function in Psoriasis
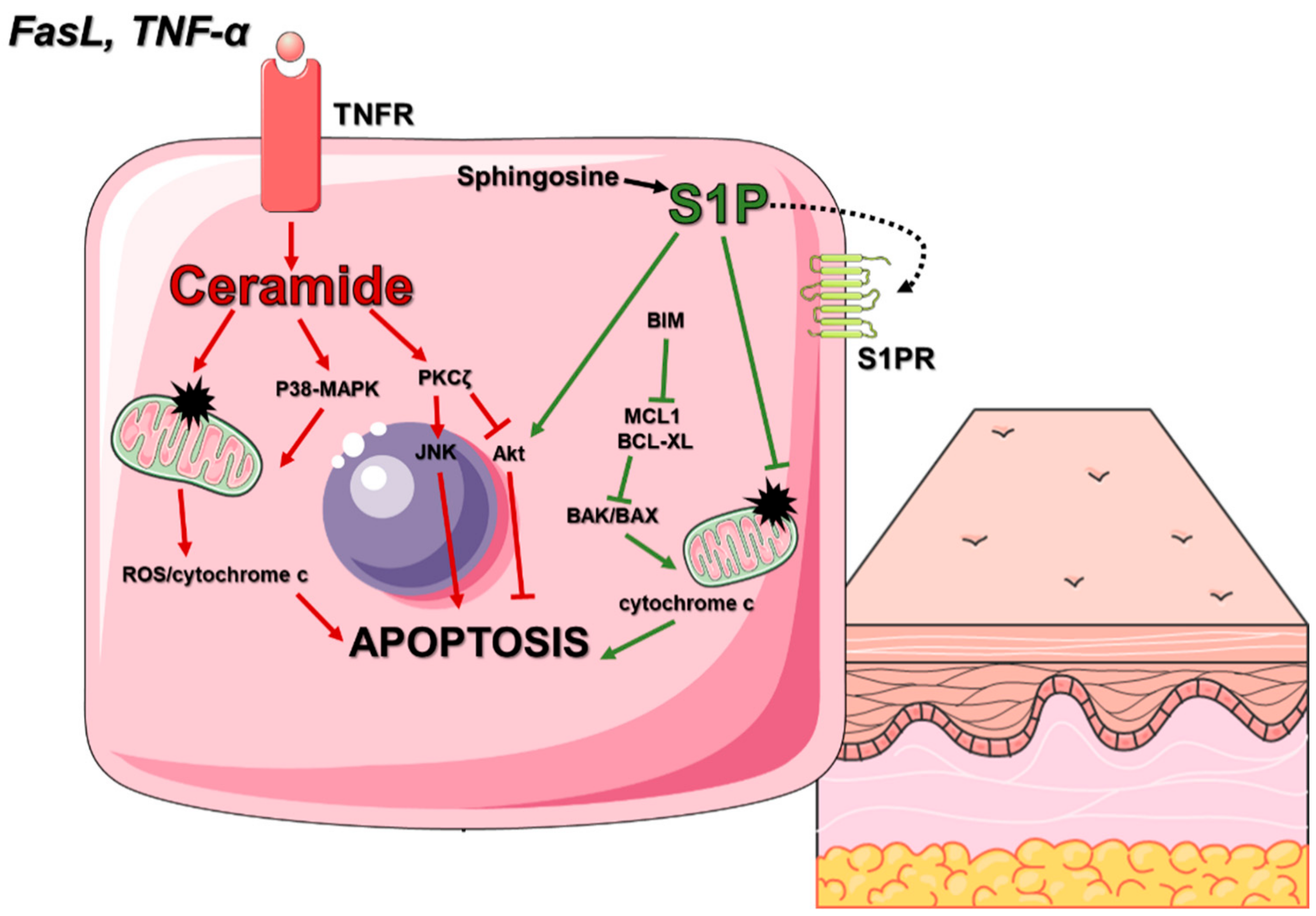
3.1.1. Cell Death In Skin Homeostasis
3.1.2. Cell Proliferation
3.1.3. Keratinocytes’ differentiation
3.2. Sphingolipid Action In Inflammation—Focus On The Molecular Mechanism Of Psoriasis
3.3. Sphingolipids In Endothelial Function—Vascular Endothelium Abnormalities in the Development of Psoriasis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [PubMed] [Green Version]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide Synthases at the Centre of Sphingolipid Metabolism and Biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tidhar, R.; Futerman, A.H. The complexity of sphingolipid biosynthesis in the endoplasmic reticulum. Biochim. Biophys. Acta 2013, 1833, 2511–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wigger, D.; Gulbins, E.; Kleuser, B.; Schumacher, F. Monitoring the Sphingolipid de novo Synthesis by Stable-Isotope Labeling and Liquid Chromatography-Mass Spectrometry. Front. Cell Dev. Biol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitatani, K.; Idkowiak-Baldys, J.; Hannun, Y.A. The sphingolipid salvage pathway in ceramide metabolism and signaling. Cell. Signal. 2008, 20, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cheng, Y.; Hansen, G.H.; Niels-Christiansen, L.L.; Koentgen, F.; Ohlsson, L.; Nilsson, A.; Duan, R.D. Crucial role of alkaline sphingomyelinase in sphingomyelin digestion: A study on enzyme knockout mice. J. Lipid Res. 2011, 52, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Yabu, T.; Imamura, S.; Yamashita, M.; Okazaki, T. Identification of Mg2+ -dependent neutral sphingomyelinase 1 as a mediator of heat stress-induced ceramide generation and apoptosis. J. Biol. Chem. 2008, 283, 29971–29982. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Y.; Sullards, M.C.; Merrill, A.H., Jr. An introduction to sphingolipid metabolism and analysis by new technologies. Neuromolecular Med. 2010, 12, 306–319. [Google Scholar] [CrossRef] [Green Version]
- Merrill, A.H., Jr. Sphingolipid and glycosphingolipid metabolic pathways in the era of sphingolipidomics. Chem. Rev. 2011, 111, 6387–6422. [Google Scholar] [CrossRef]
- Yamaji, T.; Kumagai, K.; Tomishige, N.; Hanada, K. Two sphingolipid transfer proteins, CERT and FAPP2: Their roles in sphingolipid metabolism. IUBMB Life 2008, 60, 511–518. [Google Scholar] [CrossRef]
- Kawano, M.; Kumagai, K.; Nishijima, M.; Hanada, K. Efficient trafficking of ceramide from the endoplasmic reticulum to the Golgi apparatus requires a VAMP-associated protein-interacting FFAT motif of CERT. J. Biol. Chem. 2006, 281, 30279–30288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudo, N.; Kumagai, K.; Tomishige, N.; Yamaji, T.; Wakatsuki, S.; Nishijima, M.; Hanada, K.; Kato, R. Structural basis for specific lipid recognition by CERT responsible for nonvesicular trafficking of ceramide. Proc. Natl. Acad. Sci. USA 2008, 105, 488–493. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chakravarty, D.; Maceyka, M.; Milstien, S.; Spiegel, S. Sphingosine kinases: A novel family of lipid kinases. Prog. Nucleic Acid Res. Mol. Biol. 2002, 71, 493–511. [Google Scholar] [PubMed]
- Pitson, S.M. Regulation of sphingosine kinase and sphingolipid signaling. Trends Biochem. Sci. 2011, 36, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Kono, K.; Liu, H.; Shimizugawa, T.; Minekura, H.; Spiegel, S.; Kohama, T. Ceramide kinase, a novel lipid kinase. Molecular cloning and functional characterization. J. Biol. Chem. 2002, 277, 23294–23300. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Berdyshev, E.V.; Gorshkova, I.; Usatyuk, P.; Kalari, S.; Zhao, Y.; Pyne, N.J.; Pyne, S.; Sabbadini, R.A.; Garcia, J.G.; Natarajan, V. Intracellular S1P generation is essential for S1P-induced motility of human lung endothelial cells: Role of sphingosine kinase 1 and S1P lyase. PLoS ONE 2011, 6, e16571. [Google Scholar] [CrossRef] [Green Version]
- Hatoum, D.; Haddadi, N.; Lin, Y.; Nassif, N.T.; McGowan, E.M. Mammalian sphingosine kinase (SphK) isoenzymes and isoform expression: Challenges for SphK as an oncotarget. Oncotarget 2017, 8, 36898–36929. [Google Scholar] [CrossRef] [Green Version]
- Visscher, M.; Narendran, V. The ontogeny of skin. Advances in Wound Care 2014, 3, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Borodzicz, S.; Rudnicka, L.; Mirowska-Guzel, D.; Cudnoch-Jedrzejewska, A. The role of epidermal sphingolipids in dermatologic diseases. Lipids Health Dis. 2016, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Tadeu, A.M.; Horsley, V. Epithelial stem cells in adult skin. Curr. Top. Dev. Biol. 2014, 107, 109–131. [Google Scholar]
- Kanitakis, J. Anatomy, histology and immunohistochemistry of normal human skin. Eur. J. Dermatol. 2002, 12, 390–399. [Google Scholar] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Les-sons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, A.; Roh, J.; Vávrová, K. The chemistry and biology of 6 hydroxyceramide, the youngest member of the human sphingoli-pid family. ChemBioChem 2014, 15, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Chalfant, C.E.; Spiegel, S. Sphingosine 1-phosphate and ceramide 1-phosphate: Expanding roles in cell signaling. J. Cell Sci. 2005, 118, 4605–4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.S.; Kim, S.Y.; Kleuser, B.; Schäfer-Korting, M.; Kim, K.H.; Park, K.C. Sphingosine-1-phosphate inhibits human keratinocyte proliferation via Akt/protein kinase B inactivation. Cell. Signal. 2004, 16, 89–95. [Google Scholar] [CrossRef]
- Holleran, W.M.; Takagi, Y.; Uchida, Y. Epidermal sphingolipids: Metabolism, function and roles in skin disorders. FEBS Lett. 2006, 580, 5456–5466. [Google Scholar] [CrossRef] [Green Version]
- Rabionet, M.; Gorgas, K.; Sandhoff, R. Ceramide synthesis in the epidermis. Biochim. Biophys. Acta 2014, 1841, 422–434. [Google Scholar] [CrossRef]
- Moskot, M.; Bocheńska, K.; Jakóbkiewicz-Banecka, J.; Banecki, B.; Gabig-Cimińska, M. Abnormal Sphingolipid World in Inflammation Specific for Lysosomal Storage Diseases and Skin Disorders. Int. J. Mol. Sci. 2018, 19, 247. [Google Scholar] [CrossRef] [Green Version]
- Baeyens, A.; Fang, V.; Chen, C.; Schwab, S.R. Exit strategies: S1P signaling and T cell migration. Trends Immunol. 2015, 36, 778–787. [Google Scholar] [CrossRef] [Green Version]
- Checa, A.; Xu, N.; Sar, D.G.; Haeggstrom, J.Z.; Stahle, M.; Wheelock, C.E. Circulating levels of sphingosine-1-phosphate are elevated in severe but not mild psoriasis and are unresponsive to anti-TNF-alpha treatment. Sci. Rep. 2015, 5, 12017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, R.; Symmons, D.P.; Griffiths, C.E.; Ashcroft, D.M. Global epidemiology of psoriasis: A systematic review of incidence and prevalence. J. Investig. Dermatol. 2013, 133, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, S.H.; Kim, J.Y.; Song, E.H.; Shin, M.K.; Cho, Y.H.; Kim, N.I. Altered levels of sphingosine and sphinganine in psoriatic epidermis. Ann. Dermatol. 2013, 25, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Amigo, M.; Schalkwijk, J.; Olthuis, D.; de Rosa, S.; Paya, M.; Terencio, M.C.; Lamme, E. Identification of avarol derivatives as potential antipsoriatic drugs using an in vitro model for keratinocyte growth and differentiation. Life Sci. 2006, 79, 2395–2404. [Google Scholar] [CrossRef] [PubMed]
- Saelee, C.; Thongrakard, V.; Tencomnao, T. Effects of thai medicinal herb extracts with anti-psoriatic activity on the expression on nf-kappab signaling biomarkers in hacat keratinocytes. Molecules 2011, 16, 3908–3932. [Google Scholar] [CrossRef] [Green Version]
- Reich, K. The concept of psoriasis as a systemic inflammation: Implications for disease management. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 3–11. [Google Scholar] [CrossRef]
- Sano, S. Psoriasis as a barrier disease. Dermatol. Sin. 2015, 33, 64–69. [Google Scholar]
- Cho, Y.; Lew, B.L.; Seong, K.; Kim, N.I. An inverse relationship between ceramide synthesis and clinical severity in patients with psoriasis. J. Korean Med. Sci. 2004, 19, 859–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoi, L.C.; Spain, S.L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Ding, J.; Li, Y.; Tejasvi, T.; Gudjonsson, J.E.; et al. Identification of 15 new psoriasis susceptibility loci highlights the role of innate immunity. Nat. Genet. 2012, 44, 1341–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lew, B.L.; Cho, Y.; Kim, J.; Sim, W.Y.; Kim, N.I. Ceramides and cell signaling molecules in psoriatic epidermis: Reduced levels of ceramides, PKC-alpha and JNK. J. Korean Med. Sci. 2006, 21, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Motta, S.; Monti, M.; Sesana, S.; Caputo, R.; Carelli, S.; Ghidoni, R. Ceramide composition of the psoriatic scale. Biochim. Biophys. Acta 1993, 1182, 147–151. [Google Scholar] [CrossRef]
- Nakajima, K.; Terao, M.; Takaishi, M.; Kataoka, S.; Goto-Inoue, N.; Setou, M.; Horie, K.; Sakamoto, F.; Ito, M.; Azukizawa, H.; et al. Barrier abnormality due to ceramide deficiency leads to psoriasiform inflammation in a mouse model. J. Investig. Dermatol. 2013, 133, 2555–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandrini, F.; Pfister, S.; Kremmer, E.; Gerber, J.K.; Ring, J.; Behrendt, H. Alterations of glucosylceramide-beta-glucosidase levels in the skin of patients with psoriasis vulgaris. J. Investig. Dermatol. 2004, 123, 1030–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandrini, F.; Stachowitz, S.; Ring, J.; Behrendt, H. The level of prosaposin is decreased in the skin of patients with psoriasis vulgaris. J. Investig. Dermatol. 2001, 116, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, F.; Mijouin, L.; Pichon, C. Skin Immune Landscape: Inside and Outside the Organism. Mediat. Inflamm. 2017, 2017, 17. [Google Scholar] [CrossRef]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Dermatol. 2018, 45, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Olson, D.K.; Fröhlich, F.; Farese, R.V., Jr.; Walther, T.C. Taming the sphinx: Mechanisms of cellular sphingolipid homeostasis. Biochim. Biophys. Acta 2016, 1861, 784–792. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhang, Z.; Peng, H.; Zeng, K. Recent advances on the roles of epidermal growth factor receptor in psoriasis. Am. J. Transl. Res. 2019, 11, 520–528. [Google Scholar]
- Mullen, T.D.; Obeid, L.M. Ceramide and apoptosis: Exploring the enigmatic connections between sphingolipid metabolism and programmed cell death. Anticancer. Agents Med. Chem. 2012, 12, 340–363. [Google Scholar] [CrossRef]
- Ohanian, J.; Ohanian, V. Sphingolipids in mammalian cell signalling. Cell Mol. Life Sci. 2001, 58, 2053–2068. [Google Scholar] [CrossRef]
- De Cid, R.; Riveira-Munoz, E.; Zeeuwen, P.L.; Robarge, J.; Liao, W.; Dannhauser, E.N.; Giardina, E.; Stuart, P.E.; Nair, R.; Helms, C.; et al. Deletion of the late cornified envelope LCE3B and LCE3C genes as a susceptibility factor for psoriasis. Nat. Genet. 2009, 41, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettus, B.J.; Chalfant, C.E.; Hannun, Y.A. Ceramide in apoptosis: An overview and current perspectives. Biochim. Biophys. Acta 2002, 1585, 114–125. [Google Scholar] [CrossRef]
- Young, M.M.; Kester, M.; Wang, H.G. Sphingolipids: Regulators of crosstalk between apoptosis and autophagy. J. Lipid Res. 2013, 54, 5–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Kleine, L.; Hébert, R.L. Advances in the signal transduction of ceramide and related sphingolipids. Crit. Rev. Clin. Lab. Sci. 1999, 36, 511–573. [Google Scholar] [CrossRef] [PubMed]
- Abou-Ghali, M.; Stiban, J. Regulation of ceramide channel formation and disassembly: Insights on the initiation of apoptosis. Saudi. J. Biol. Sci. 2015, 22, 760–772. [Google Scholar] [CrossRef] [Green Version]
- Hait, N.C.; Maiti, A. The Role of Sphingosine-1-Phosphate and Ceramide-1-Phosphate in Inflammation and Cancer. Mediat. Inflamm. 2017, 2017, 17. [Google Scholar] [CrossRef]
- Lin, C.C.; Lee, I.T.; Hsu, C.H.; Hsu, C.K.; Chi, P.L.; Hsiao, L.D.; Yang, C.M. Sphingosine-1-phosphate mediates ICAM-1-dependent monocyte adhesion through p38 MAPK and p42/p44 MAPK-dependent Akt activation. PLoS ONE 2015, 10, e0118473. [Google Scholar] [CrossRef] [Green Version]
- Hait, N.C.; Oskeritzian, C.A.; Paugh, S.W.; Milstien, S.; Spiegel, S. Sphingosine kinases, sphingosine 1-phosphate, apoptosis and diseases. Biochim. Biophys. Acta 2006, 1758, 2016–2026. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Smith, C.D.; Foroozesh, M.; Miele, L.; Qin, Z. The sphingosine kinase 2 inhibitor ABC294640 displays anti-non-small cell lung cancer activities in vitro and in vivo. Int. J. Cancer 2018, 142, 2153–2162. [Google Scholar] [CrossRef] [Green Version]
- Andrieu, G.; Ledoux, A.; Branka, S.; Bocquet, M.; Gilhodes, J.; Walzer, T.; Kasahara, K.; Inagaki, M.; Sabbadini, R.A.; Cuvillier, O.; et al. Sphingosine 1-phosphate signaling through its receptor S1P5 promotes chromosome segregation and mitotic progression. Sci. Signal. 2017, 472, eaah4007. [Google Scholar] [CrossRef]
- Bonhoure, E.; Lauret, A.; Barnes, D.J.; Martin, C.; Malavaud, B.; Kohama, T.; Melo, J.V.; Cuvillier, O. Sphingosine kinase-1 is a downstream regulator of imatinib-induced apoptosis in chronic myeloid leukemia cells. Leukemia 2008, 22, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Betito, S.; Cuvillier, O. Regulation by sphingosine 1-phosphate of Bax and Bad activities during apoptosis in a MEK-dependent manner. Biochem. Biophys. Res. Commun. 2006, 340, 1273–1277. [Google Scholar] [CrossRef] [PubMed]
- Akao, Y.; Banno, Y.; Nakagawa, Y.; Hasegawa, N.; Kim, T.J.; Murate, T.; Igarashi, Y.; Nozawa, Y. High expression of sphingosine kinase 1 and S1P receptors in chemotherapy-resistant prostate cancer PC-3 cells and their camptothecin-induced up-regulation. Biochem. Biophys. Res. Commun. 2006, 342, 1284–1290. [Google Scholar] [CrossRef]
- Guillermet-Guibert, J.; Davenne, L.; Pchejetski, D.; Saint-Laurent, N.; Brizuela, L.; Guilbeau-Frugier, C.; Delisle, M.B.; Cuvillier, O.; Susini, C.; Bousquet, C. Targeting the sphingolipid metabolism to defeat pancreatic cancer cell resistance to the chemotherapeutic gemcitabine drug. Mol. Cancer Ther. 2009, 8, 809–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, Y. Ceramide signaling in mammalian epidermis. Biochim. Biophys. Acta 2014, 1841, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breiden, B.; Sandhoff, K. The role of sphingolipid metabolism in cutaneous permeability barrier formation. Biochim. Biophys. Acta 2014, 1841, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Sahle, F.F.; Gebre-Mariam, T.; Dobner, B.; Wohlrab, J.; Neubert, R.H. Skin diseases associated with the depletion of stratum corneum lipids and stratum corneum lipid substitution therapy. Skin. Pharmacol. Physiol. 2015, 28, 42–55. [Google Scholar] [CrossRef]
- Van Smeden, J.; Bouwstra, J.A. Stratum corneum lipids: Their role for the skin barrier function in healthy subjects and atopic dermatitis patients. Curr. Probl. Dermatol. 2016, 49, 8–26. [Google Scholar]
- Pasquali, L.; Srivastava, A.; Meisgen, F.; Das Mahapatra, K.; Xia, P.; Xu Landén, N.; Pivarcsi, A.; Sonkoly, E. The Keratinocyte Transcriptome in Psoriasis: Pathways Related to Immune Responses, Cell Cycle and Keratinization. Acta Derm. Venereol. 2019, 99, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Deo, P.N.; Deshmukh, R. Pathophysiology of keratinization. J. Oral. Maxillofac. Pathol. 2018, 22, 86–91. [Google Scholar]
- Bocheńska, K.; Moskot, M.; Malinowska, M.; Jakóbkiewicz-Banecka, J.; Szczerkowska-Dobosz, A.; Purzycka-Bohdan, D.; Pleńkowska, J.; Słomiński, B.; Gabig-Cimińska, M. Lysosome Alterations in the Human Epithelial Cell Line HaCaT and Skin Specimens: Relevance to Psoriasis. Int. J. Mol. Sci. 2019, 20, E2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolińska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Węgrzyn, G.; Banecki, B.; Szczerkowska-Dobosz, A.; Purzycka-Bohdan, D.; Gabig-Cimińska, M. Molecular action of isoflavone genistein in the human epithelial cell line HaCaT. PLoS ONE 2018, 14, e0192297. [Google Scholar] [CrossRef] [PubMed]
- Manggau, M.; Kim, D.S.; Ruwisch, L.; Vogler, R.; Korting, H.C.; Schäfer-Korting, M.; Kleuser, B. 1Alpha,25-dihydroxyvitamin D3 protects human keratinocytes from apoptosis by the formation of sphingosine-1-phosphate. J. Investig. Dermatol. 2001, 117, 1241–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Morales, J.C.; Romero-Ávila, M.T.; Reyes-Cruz, G.; García-Sáinz, J.A. S1P1 receptor phosphorylation, internalization, and interaction with Rab proteins: Effects of sphingosine 1-phosphate, FTY720-P, phorbol esters, and paroxetine. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendelson, K.; Evans, T.; Hla, T. Sphingosine 1-phosphate signalling. Development 2014, 141, 5–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Fernández, B.; Sánchez, D.I.; González-Gallego, J.; Tuñón, M.J. Sphingosine 1-Phosphate Signaling as a Target in Hepatic Fibrosis Therapy. Front. Pharmacol. 2017, 25, 579. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.C.; Han, M.H. Sphingosine-1-Phosphate (S1P) and S1P Signaling Pathway: Therapeutic Targets in Autoimmunity and Inflammation. Drugs 2016, 76, 1067–1079. [Google Scholar] [CrossRef]
- Kraveka, J.M.; Li, L.; Bielawski, J.; Obeid, L.M.; Ogretmen, B. Involvement of endogenous ceramide in the inhibition of telomerase activity and induction of morphologic differentiation in response to all-trans-retinoic acid in human neuroblastoma cells. Arch Biochem. Biophys. 2003, 419, 110–119. [Google Scholar] [CrossRef]
- Van Brocklyn, J.R.; Williams, J.B. The control of the balance between ceramide and sphingosine-1-phosphate by sphingosine kinase: Oxidative stress and the seesaw of cell survival and death. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2012, 163, 26–36. [Google Scholar] [CrossRef]
- Foskett, J.K.; White, C.; Cheung, K.H.; Mak, D.O. Inositol trisphosphate receptor Ca2+ release channels. Physiol. Rev. 2007, 87, 593–658. [Google Scholar] [CrossRef] [Green Version]
- Borowiec, A.S.; Delcourt, P.; Dewailly, E.; Bidaux, G. Optimal differentiation of in vitro keratinocytes requires multifactorial external control. PLoS ONE 2013, 10, e77507. [Google Scholar] [CrossRef] [Green Version]
- Arul, S.; Dayalan, H.; Jegadeesan, M.; Damodharan, P. Induction of differentiation in psoriatic keratinocytes by propylthiouracil and fructose. BBA Clin. 2016, 6, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Sayama, K.; Hanakawa, Y.; Shirakata, Y.; Yamasaki, K.; Sawada, Y.; Sun, L.; Yamanishi, K.; Ichijo, H.; Hashimoto, K. Apoptosis signal-regulating kinase 1 (ASK1) is an intracellular inducer of keratinocyte differentiation. J. Biol. Chem. 2001, 276, 999–1004. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.J.; Kim, P.; Uchida, Y.; Elias, P.M.; Bikle, D.D.; Grunfeld, C.; Feingold, K.R. Ceramides stimulate caspase-14 expression in human keratinocytes. Exp. Dermatol. 2013, 22, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Meyer Zu Heringdorf, D. Lysophospholipid receptor-dependent and -independent calcium signaling. J. Cell Biochem. 2004, 92, 937–948. [Google Scholar] [CrossRef]
- Mehta, D.; Konstantoulaki, M.; Ahmmed, G.U.; Malik, A.B. Sphingosine 1-phosphate-induced mobilization of intracellular Ca2+ mediates rac activation and adherens junction assembly in endothelial cells. J. Biol. Chem. 2005, 280, 17320–17328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichte, K.; Rossi, R.; Danneberg, K.; Ter Braak, M.; Kurschner, U.; Jakobs, K.H.; Kleuser, B.; Zu Meyer Heringdorf, D. Lysophospholipid receptor-mediated calcium signaling in human keratinocytes. J. Investig. Dermatol. 2008, 128, 1487–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, Y.; Houben, E.; Park, K. Hydrolytic pathway protects against ceramide-induced apoptosis in keratinocytes exposed to UVB. J. Investig. Dermatol. 2010, 130, 2472–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.; Lee, S.; Lee, Y.M. Sphingolipids and antimicrobial peptides: Function and roles in atopic dermatitis. Biomol. Ther. 2013, 21, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Elias, P.M.; Shin, K.O.; Lee, Y.M.; Hupe, M.; Borkowski, A.W.; Gallo, R.L.; Saba, J.; Holleran, W.M.; Uchida, Y. A novel role of a lipid species, sphingosine-1-phosphate, in epithelial innate immunity. Mol. Cell Biol. 2013, 33, 752–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lande, R.; Gregorio, J.; Facchinetti, V.; Chatterjee, B.; Wang, Y.H.; Homey, B.; Cao, W.; Wang, Y.H.; Su, B.; Nestle, F.O.; et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 2007, 449, 564–569. [Google Scholar] [CrossRef]
- Bocheńska, K.; Smolińska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Gabig-Cimińska, M. Models in the Research Process of Psoriasis. Int. J. Mol. Sci. 2017, 24, E2514. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.G.; Kim, D.S.; Kim, H.P.; Lee, M.G. The pathophysiological role of dendritic cell subsets in psoriasis. BMB Rep. 2014, 47, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Pritz, C.J.; Seo, Y.J.; Xia, C.; Vijayan, M.; Stokes, Z.D.; Hahm, B. A ceramide analogue stimulates dendritic cells to promote T cell responses upon virus infections. J. Immunol. 2015, 194, 4339–4349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arlt, O.; Schwiebs, A.; Japtok, L.; Rüger, K.; Katzy, E.; Kleuser, B.; Radeke, H.H. Sphingosine-1-phosphate modulates dendritic cell function: Focus on non-migratory effects in vitro and in vivo. Cell Physiol. Biochem. 2014, 34, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Wang, T.; Zhu, C.; Xing, X.; Ye, Y.; Lai, X.; Song, B.; Zeng, Y. Topographical and biological evidence revealed FTY720-mediated anergy-polarization of mouse bone marrow-derived dendritic cells in vitro. PLoS ONE 2012, 7, e34830. [Google Scholar] [CrossRef] [PubMed]
- Collenburg, L.; Schneider-Schaulies, S.; Avota, E. The neutral sphingomyelinase 2 in T cell receptor signaling and polarity. Biol. Chem. 2018, 399, 1147–1155. [Google Scholar] [CrossRef]
- Boucher, L.M.; Wiegmann, K.; Fütterer, A.; Pfeffer, K.; Machleidt, T.; Schütze, S.; Mak, T.W.; Krönke, M. CD28 signals through acidic sphingomyelinase. J. Exp. Med. 1995, 181, 2059–2068. [Google Scholar] [CrossRef] [Green Version]
- Tonnetti, L.; Verí, M.C.; Bonvini, E.; D’Adamio, L. A role for neutral sphingomyelinase-mediated ceramide production in T cell receptor-induced apoptosis and mitogen-activated protein kinase-mediated signal transduction. J. Exp. Med. 1999, 189, 1581–1589. [Google Scholar] [CrossRef]
- Mueller, N.; Avota, E.; Collenburg, L.; Grassmé, H.; Schneider-Schaulies, S. Neutral sphingomyelinase in physiological and measles virus induced T cell suppression. PLoS Pathog. 2014, 10, e1004574. [Google Scholar] [CrossRef]
- Liao, W.; Lin, J.X.; Leonard, W.J. Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity 2013, 38, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abboushi, N.; El-Hed, A.; El-Assaad, W.; Kozhaya, L.; El-Sabban, M.E.; Bazarbachi, A.; Badreddine, R.; Bielawska, A.; Usta, J.; Dbaibo, G.S. Ceramide inhibits IL-2 production by preventing protein kinase C-dependent NF-kappaB activation: Possible role in protein kinase Ctheta regulation. J. Immunol. 2004, 173, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Bai, A.; Guo, Y. Acid sphingomyelinase mediates human CD4+ T-cell signaling: Potential roles in T-cell responses and diseases. Cell Death Dis. 2017, 8, e2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, A.; Kokkotou, E.; Zheng, Y.; Robson, S.C. Role of acid sphingomyelinase bioactivity in human CD4+ T-cell activation and immune responses. Cell Death Dis. 2015, 6, e1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, L.D.; Hessler, G.; Goodall, J.E.; Rider, D.A.; Workman, C.J.; Vignali, D.A.; Bacon, P.A.; Gulbins, E.; Young, S.P. TNFR1-induced sphingomyelinase activation modulates TCR signaling by impairing store-operated Ca2+ influx. J. Leukoc. Biol. 2005, 78, 266–278. [Google Scholar] [CrossRef]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef] [Green Version]
- Molino, S.; Tate, E.; McKillop, W.M.; Medin, J.A. Sphingolipid pathway enzymes modulate cell fate and immune responses. Immunotherapy 2017, 14, 1185–1198. [Google Scholar] [CrossRef]
- Xiong, Y.; Piao, W.; Brinkman, C.C.; Li, L.; Kulinski, J.M.; Olivera, A.; Cartier, A.; Hla, T.; Hippen, K.L.; Blazar, B.R.; et al. CD4 T cell sphingosine 1-phosphate receptor (S1PR)1 and S1PR4 and endothelial S1PR2 regulate afferent lymphatic migration. Sci. Immunol. 2019, 33, eaav1263. [Google Scholar] [CrossRef]
- Herz, J.; Pardo, J.; Kashkar, H.; Schramm, M.; Kuzmenkina, E.; Bos, E.; Wiegmann, K.; Wallich, R.; Peters, P.J.; Herzig, S.; et al. Acid sphingomyelinase is a key regulator of cytotoxic granule secretion by primary T lymphocytes. Nat. Immunol. 2009, 10, 761–768. [Google Scholar] [CrossRef]
- Di Cesare, A.; Di Meglio, P.; Nestle, F.O. The IL-23/Th17 axis in the immunopathogenesis of psoriasis. J. Investig. Dermatol. 2009, 129, 1339–1350. [Google Scholar] [CrossRef] [Green Version]
- Di Meglio, P.; Nestle, F.O. The role of IL-23 in the immunopathogenesis of psoriasis. F1000 Biol. Rep. 2010, 2, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaumik, S.; Basu, R. Cellular and Molecular Dynamics of Th17 Differentiation and its Developmental Plasticity in the Intestinal Immune Response. Front. Immunol. 2017, 8, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayama, H. Development of psoriasis by continuous neutrophil infiltration into the epidermis. Exp. Dermatol. 2018, 10, 1084–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espaillat, M.P.; Kew, R.R.; Obeid, L.M. Sphingolipids in neutrophil function and inflammatory responses: Mechanisms and implications for intestinal immunity and inflammation in ulcerative colitis. Adv. Biol. Regul. 2017, 63, 140–155. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Galisteo, R.; Gutkind, J.S. CXCL8/IL8 stimulates vascular endothelial growth factor (VEGF) expression and the autocrine activation of VEGFR2 in endothelial cells by activating NFkappaB through the CBM (Carma3/Bcl10/Malt1) complex. J. Biol. Chem. 2009, 284, 6038–6042. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Guo, Y.; Zhang, C.; Fan, F.; Yang, W. Sphingosine Kinase 1 and Sphingosine-1-Phosphate Signaling in Colorectal Cancer. Int. J. Mol. Sci. 2017, 8, E2109. [Google Scholar] [CrossRef] [Green Version]
- Langlois, S.; Nyalendo, C.; Di Tomasso, G.; Labrecque, L.; Roghi, C.; Murphy, G.; Gingras, D.; Béliveau, R. Membrane-type 1 matrix metalloproteinase stimulates cell migration through epidermal growth factor receptor transactivation. Mol. Cancer Res. 2007, 6, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Syed, S.N.; Raue, R.; Weigert, A.; von Knethen, A.; Brüne, B. Macrophage S1PR1 Signaling Alters Angiogenesis and Lymphangiogenesis During Skin Inflammation. Cells 2019, 8, E785. [Google Scholar] [CrossRef] [Green Version]
- Weigert, A.; Olesch, C.; Brüne, B. Sphingosine-1-Phosphate and Macrophage Biology-How the Sphinx Tames the Big Eater. Front. Immunol. 2019, 10, 1706. [Google Scholar] [CrossRef]
- Adada, M.; Canals, D.; Hannun, Y.A.; Obeid, L.M. Sphingosine-1-phosphate receptor 2. FEBS J. 2013, 280, 6354–6366. [Google Scholar] [CrossRef] [Green Version]
- Mak, R.K.; Hundhausen, C.; Nestle, F.O. Progress in understanding the immunopathogenesis of psoriasis. Actas Dermosifiliogr. 2009, 100 (Suppl. 2), 2–13. [Google Scholar] [CrossRef] [Green Version]
- Louis, S.F.; Zahradka, P. Vascular smooth muscle cell motility: From migration to invasion. Exp. Clin. Cardiol. 2010, 15, e75–e85. [Google Scholar] [PubMed]
- Heidenreich, R.; Röcken, M.; Ghoreschi, K. Angiogenesis drives psoriasis pathogenesis. Int. J. Exp. Pathol. 2009, 90, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, M.; Tölle, M.; Prüfer, J.; van der Giet, M. Pharmacological relevance and potential of sphingosine 1-phosphate in the vascular system. Br. J. Pharmacol. 2011, 163, 1140–1162. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Shaaya, M.; Harijith, A.; Jacobson, J.R.; Karginov, A.; Natarajan, V. Sphingolipids Signaling in Lamellipodia Formation and Enhancement of Endothelial Barrier Function. Curr. Top. Membr. 2018, 82, 1–31. [Google Scholar] [PubMed]
- Langlois, S.; Gingras, D.; Béliveau, R. Membrane type 1-matrix metalloproteinase (MT1-MMP) cooperates with sphingosine 1-phosphate to induce endothelial cell migration and morphogenic differentiation. Blood 2004, 103, 3020–3028. [Google Scholar] [CrossRef] [Green Version]
- Jernigan, P.L.; Makley, A.T.; Hoehn, R.S.; Edwards, M.J.; Pritts, T.A. The role of sphingolipids in endothelial barrier function. Biol. Chem. 2015, 396, 681–691. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocheńska, K.; Gabig-Cimińska, M. Unbalanced Sphingolipid Metabolism and Its Implications for the Pathogenesis of Psoriasis. Molecules 2020, 25, 1130. https://doi.org/10.3390/molecules25051130
Bocheńska K, Gabig-Cimińska M. Unbalanced Sphingolipid Metabolism and Its Implications for the Pathogenesis of Psoriasis. Molecules. 2020; 25(5):1130. https://doi.org/10.3390/molecules25051130
Chicago/Turabian StyleBocheńska, Katarzyna, and Magdalena Gabig-Cimińska. 2020. "Unbalanced Sphingolipid Metabolism and Its Implications for the Pathogenesis of Psoriasis" Molecules 25, no. 5: 1130. https://doi.org/10.3390/molecules25051130
APA StyleBocheńska, K., & Gabig-Cimińska, M. (2020). Unbalanced Sphingolipid Metabolism and Its Implications for the Pathogenesis of Psoriasis. Molecules, 25(5), 1130. https://doi.org/10.3390/molecules25051130