
The Use of Fluorescence Spectrometry to Determine the Botanical Origin of Filtered Honeys


Abstract
:1. Introduction
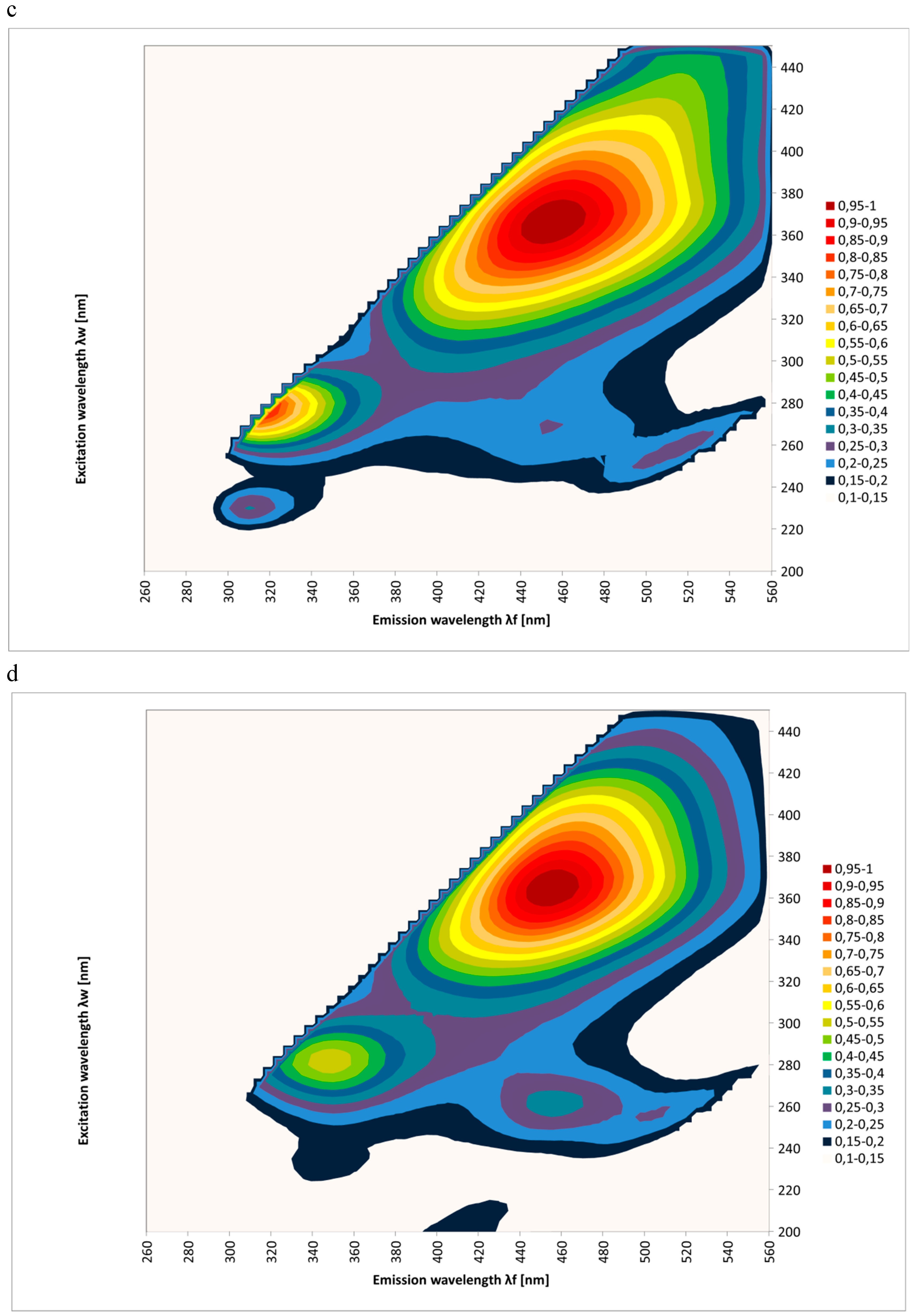
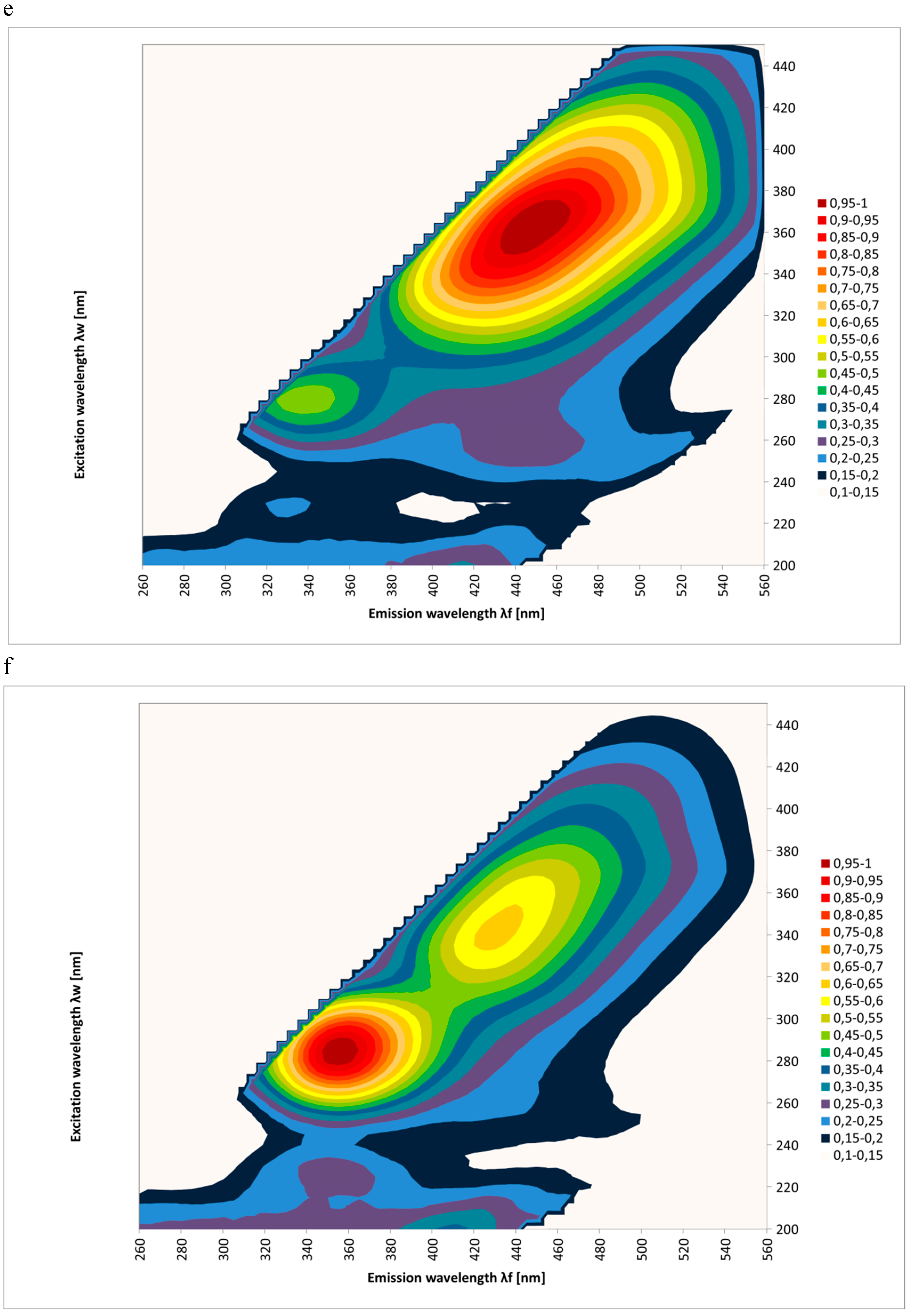
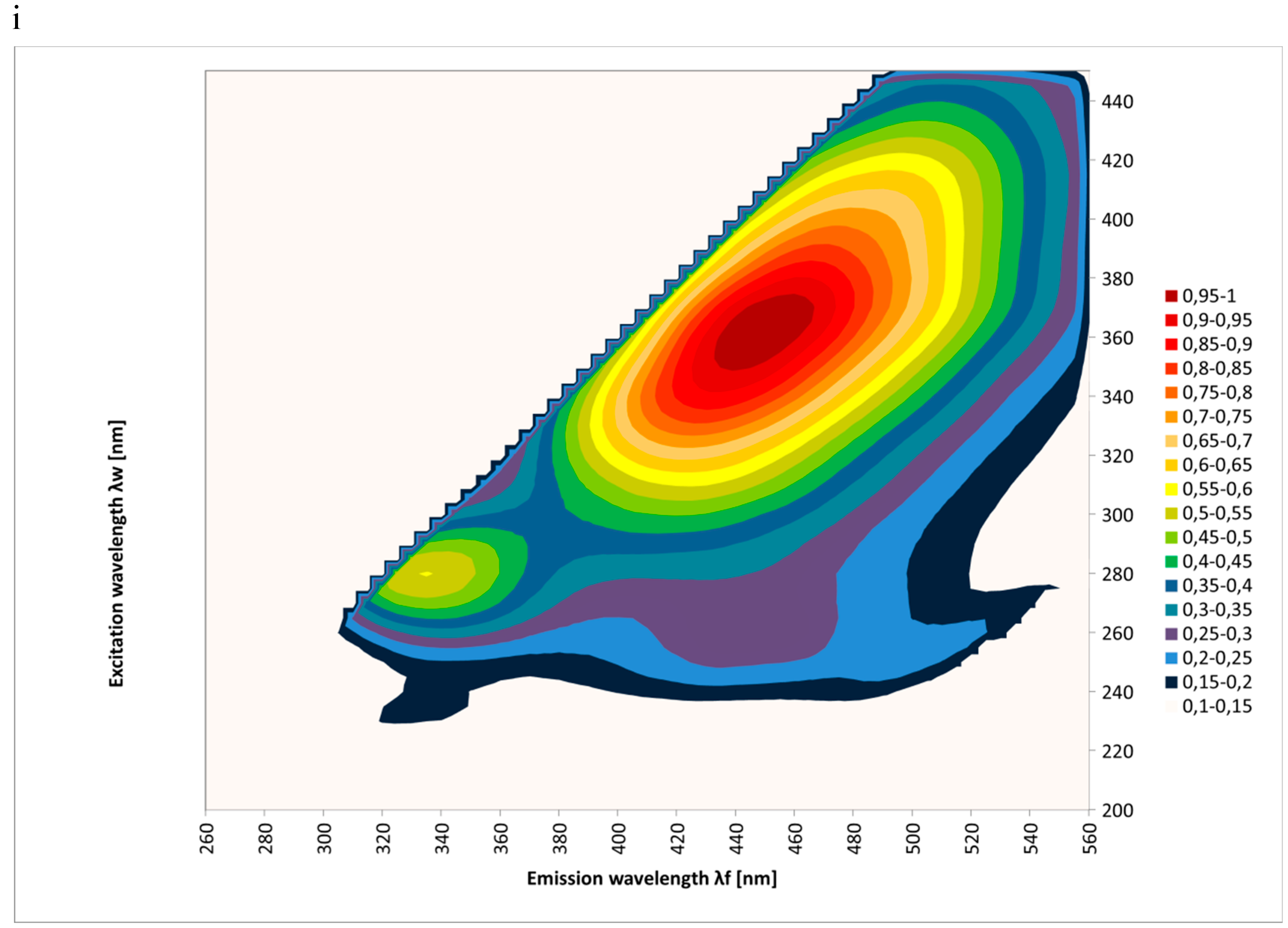
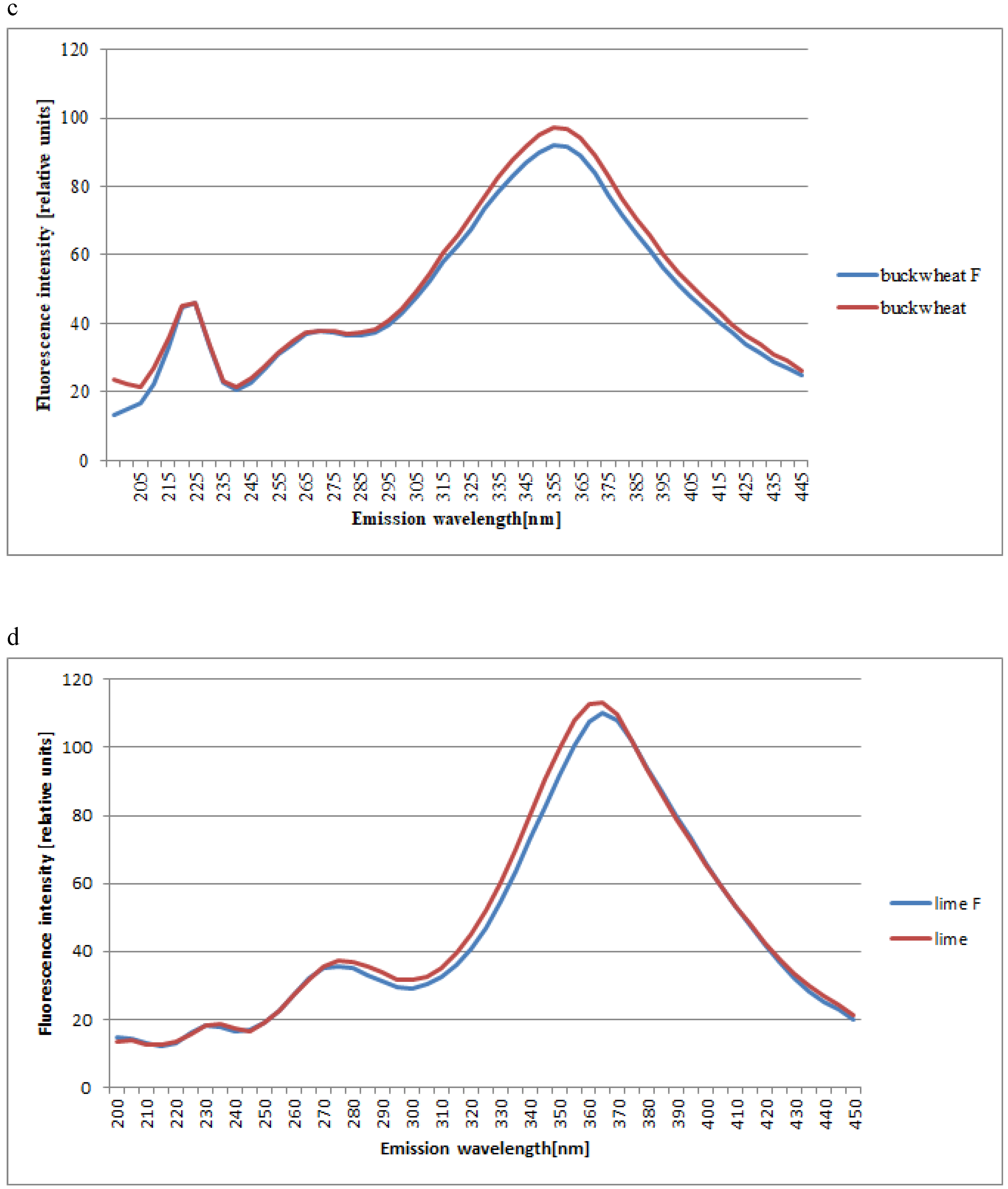
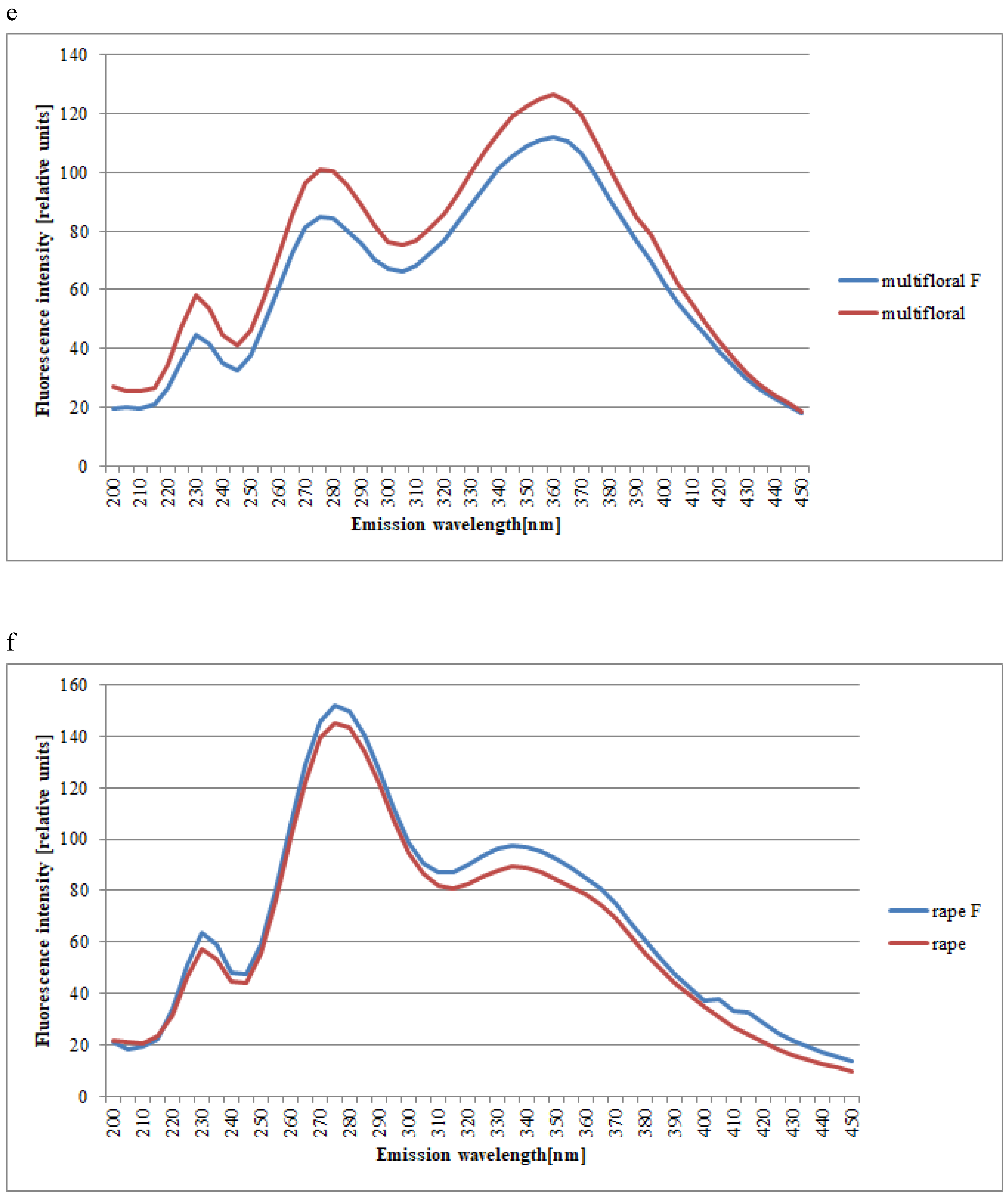
2. Results and Discussion
3. Materials and Methods
3.1. Samples
3.2. Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bogdanov, S.; Jurendic, T.; Sieber, R.; Gallmann, P. Honey for nutrition and health: A review. JACN 2008, 27, 677–689. [Google Scholar] [CrossRef] [PubMed]
- De Silva, P.M.; Gauche, C.; Gonzaga, L.V.; Costa, A.C.O. Honey: Chemical composition, stability and authenticity. Food Chem. 2016, 196, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Escuredo, O.; Dobre, I.; Fernández-González, M.; Seijo, M.C. Contribution of botanical origin and sugar composition of honeys on the crystallization phenomenon. Food Chem. 2014, 149, 84–90. [Google Scholar] [CrossRef]
- Kaskoniene, V.; Venskutonis, P.R.; Ceksterytè, V. Carbohydrate composition and electrical conductivity of different origin honeys from Lithuania. LWT-Food Sci. Technol. 2010, 43, 801–807. [Google Scholar] [CrossRef]
- Soares, S.; Amaral, J.S.; Oliveira, M.B.P.P.; Mafra, I. A comprehensive review on the main honey authentication issues: Production and origin, Comp. Rev. Food Sci. and Food Safety. 2017, 16, 1072–1100. [Google Scholar] [CrossRef] [Green Version]
- Matysiak-Żurowska, D.; Borowicz, A. A comparison of spectrophotometric Winkler method and HPLC technique for determination of 5-hydroxymethylfurfural in natural honey. Chem. Anal. 2009, 54, 939–947. [Google Scholar]
- Beckmann, K.; Beckh, G.; Luellmann, C.; Speer, K. Characterization of filtered honey by electrophoresis of enzyme fractions. Apidologie 2010, 42, 59. [Google Scholar] [CrossRef] [Green Version]
- Wilczyńska, A. Effect of filtration on colour, antioxidant activity and total phenolics of honey. LWT-Food Sci. Technol. 2014, 57, 767–774. [Google Scholar] [CrossRef]
- Juan-Borrás, M.; Domenech, E.; Hellebrandova, M.; Escriche, I. Effect of country origin on physicochemical, sugar and volatile composition of acacia, sunflower and tilia honeys. Food Res. Int. 2014, 60, 86–94. [Google Scholar] [CrossRef]
- Escriche, I.; Kadar, M.; Juan-Borrás, M.; Domenech, E. Using flavonoids, phenolic compounds and headspace volatile profile for botanical authentication of lemon and orange honeys. Food Res. Int. 2011, 44, 1504–1513. [Google Scholar] [CrossRef]
- Persano-Oddo, L.; Bogdanov, S. Determination of honey botanical origin: Problems and issues. Apidologie 2004, 35, 2–3. [Google Scholar] [CrossRef] [Green Version]
- Aliferis, K.A.; Ttarantilis, P.A.; Harizanis, P.C.; Alissandrakis, E. Botanical discrimination and classification of honey samples applying gas chromatography/mass spectrometry fingerprinting of headspace volatile compounds. Food Chem. 2010, 121, 856–862. [Google Scholar] [CrossRef]
- Ruoff, K.; Luginbuehl, W.; Bogdanov, S.; Bosset, J.O.; Esterman, B.; Ziorko, T.; Kheradmandan, S.; Amad, R. Quantitative determination of physical and chemical measurands in honey by near-infrared spectrometry. Eur. Food Res. Technol. 2007, 225, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Terrab, A.; Gustavo-González, A.; Díez, M.J.; Heredia, F.J. Characterization of Moroccan unifloral honeys using multivariate analysis. Eur. Food Res. Technol. 2003, 218, 88–95. [Google Scholar] [CrossRef]
- Oroian, M.; Ropciuc, S.; Buculei, A. Romanian honey authentication based on physico-chemical parameters and chemometrics. J. Food Measur. Charac. 2017, 11, 719–725. [Google Scholar] [CrossRef]
- Popek, S.; Halagarda, M.; Kursa, K. A new model to identify botanical origin of Polish honeys based on the physicochemical parameters and chemometric analysis. LWT–Food Sci. Technol. 2017, 77, 482–487. [Google Scholar] [CrossRef]
- Kuś, P.M.; van Ruth, S. Discrimination of Polish unifloral honeys using overall PTR-MS and HPLC fingerprints combined with chemometrics. LWT–Food Sci. Technol. 2015, 62, 69–75. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Muzolf-Panek, M.; Tomaszewska-Gras, J.; Konieczny, P. Predicting the botanical origin of honeys with chemometric analysis according to their antioxidant and physicochemical properties. Pol. J. Food Nutr. Sci. 2019, 69, 191–201. [Google Scholar] [CrossRef]
- Dżugan, M.; Tomczyk, M.; Sowa, P.; Grabek-Lejko, D. Antioxidant Activity as Biomarker of Honey Variety. Molecules 2018, 23, 2069. [Google Scholar] [CrossRef] [Green Version]
- Kropf, U.; Korosec, M.; Bertoncelj, J.; Ogrinc, N.; Necemer, M.; Kump, P.; Golob, T. Determination of the geographical origin of Slovenian black locust, lime and chestnut honey. Food Chem. 2010, 121, 839–846. [Google Scholar] [CrossRef]
- Etzold, E.; Lichtenberg-Kraag, B. Determination of the botanical origin of honey by Fourier-transformed infrared spectroscopy: An approach for routine analysis. Eur. Food Res. Technol. 2007, 227, 579–586. [Google Scholar] [CrossRef]
- Lenhardt, L.; Zekovic, I.; Dramicanin, T.; Dramicanin, M.D.; Bro, R. Determination of the botanical origin of honey by front-face synchronous fluorescence spectroscopy. Appl. Spectroscopy. 2014, 68, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Ruoff, K. Authentication of the Botanical Origin of Honey. Doctoral Thesis, ETH Zurich, Zurich, Switzerland, 2006. [Google Scholar] [CrossRef]
- Karoui, R.; Dufour, E.; Bosset, J.-O.; De Baerdemaeker, J. The use of front face fluorescence spectroscopy to classify the botanical origin of honey samples produced in Switzerland. Food Chem. 2007, 101, 314–323. [Google Scholar] [CrossRef]
- Lenhardt, L.; Bro, R.; Zekovic, I.; Dramicanin, T.; Dramicanin, M.D. Fluorescence spectroscopy coupled with PARAFAC and PLS DA for characterization and classification of honey. Food Chem. 2015, 175, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Ruoff, K.; Karoui, R.; Dufour, E.; Luginbuhl, W.; Bosset, J.O.; Bogdanov, S.; Amadò, R. Authentication of the botanical origin of honey by front-face fluorescence spectroscopy, a preliminary study. J. Agric. Food Chem. 2005, 53, 1343–1347. [Google Scholar] [CrossRef]
- Ruoff, K.; Luginbühl, W.; Künzli, R.; Bogdanov, S.; Bosset, J.O.; von der Ohe, K.; von der Ohe, W.; Amadò, R. Authentication of the botanical and geographical origin of honey by front-face fluorescence spectroscopy. J. Agric. Food Chem. 2006, 54, 6858–6866. [Google Scholar] [CrossRef]
- Drami´canin, T.; Lenhardt Ackovic, L.; Zekovic, I.; Dramicanin, M.D. Detection of Adulterated Honey by Fluorescence Excitation-Emission Matrices. J. Spectrosc. 2018, 8395212. [Google Scholar] [CrossRef] [Green Version]
- Gebala, S.; Przybylowski, P.; Borawska, M.H.; Piekut, J. Klasyfikacja naturalnych miodów pszczelich na podstawie analizy ksztaltu widm fluorescencyjnych (Classification of natural honeys based on the analysis of surface spectrofluorimetry shapes). Brom. Chem. Toksykol. 2005, 627–631. [Google Scholar]
- Gębala, S. Measurements of solution fluorescence–a new concept. Opt. Appl. 2009, 39, 391–399. [Google Scholar]
- Gębala, S.; Przybyłowski, P. Sposób Identyfikacji Odmian Miodu. Polska Patent nr 214784, 15 March 2010. [Google Scholar]
Sample Availability: Samples of the honeys are available from the authors. |














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectral Region | Excitation | Type of Honey | Kind of Processing | Mean Intensities of Fluorescence | SD |
|---|---|---|---|---|---|
| 1st | 200 | dandelion | unfiltered | 21.19 | 1.69 |
| 1st | 230 | honeydew | 33.34 | 3.43 | |
| 1st | 230 | goldenrods | 8.96 | 2.62 | |
| 1st | 230 | rape | 45.78 | 2.82 | |
| 1st | 230 | multifloral | 58.15 | 3.98 | |
| 1st | 230 | acacia | 51.04 | 3.50 | |
| 1st | 235 | phacelia | 41.46 | 3.62 | |
| 2nd | 275 | rape | 37.59 | 2.26 | |
| 2nd | 275 | lime | 77.52 | 9.15 | |
| 2nd | 275 | multifloral | 100.96 | 6.17 | |
| 2nd | 275 | buckwheat | 37.59 | 2.21 | |
| 2nd | 275 | phacelia | 63.43 | 4.26 | |
| 2nd | 280 | honeydew | 59.42 | 6.32 | |
| 2nd | 280 | acacia | 86.65 | 5.83 | |
| 2nd | 285 | dandelion | 110.27 | 6.28 | |
| 3rd | 335 | rape | 67.70 | 2.46 | |
| 3rd | 335 | acacia | 108.37 | 4.63 | |
| 3rd | 355 | dandelion | 106.82 | 2.46 | |
| 3rd | 355 | phacelia | 108.45 | 3.96 | |
| 3rd | 360 | honeydew | 114.42 | 4.69 | |
| 3rd | 360 | multifloral | 126.41 | 4.19 | |
| 3rd | 360 | buckwheat | 91.98 | 2.90 | |
| 3rd | 365 | goldenrods | 64.49 | 4.13 | |
| 3rd | 365 | lime | 107.44 | 4.13 | |
| 1st | 200 | dandelion | filtered | 25.41 | 1.69 |
| 1st | 230 | honeydew | 43.98 | 2.96 | |
| 1st | 230 | goldenrods | 10.66 | 4.94 | |
| 1st | 230 | rape | 45.59 | 3.06 | |
| 1st | 230 | multifloral | 44.56 | 3.79 | |
| 1st | 230 | acacia | 37.89 | 3.78 | |
| 1st | 235 | phacelia | 40.16 | 3.05 | |
| 2nd | 275 | rape | 38.65 | 2.21 | |
| 2nd | 275 | lime | 35.35 | 1.74 | |
| 2nd | 275 | multifloral | 84.70 | 6.76 | |
| 2nd | 275 | buckwheat | 37.91 | 2.20 | |
| 2nd | 275 | phacelia | 65.57 | 3.96 | |
| 2nd | 275 | acacia | 80.11 | 5.39 | |
| 2nd | 280 | honeydew | 82.98 | 5.58 | |
| 2nd | 285 | dandelion | 105.81 | 6.07 | |
| 3rd | 335 | rape | 72.68 | 2.78 | |
| 3rd | 340 | acacia | 100.66 | 3.69 | |
| 3rd | 350 | dandelion | 103.96 | 2.71 | |
| 3rd | 355 | phacelia | 115.21 | 3.68 | |
| 3rd | 360 | honeydew | 124.82 | 4.69 | |
| 3rd | 360 | goldenrods | 69.06 | 6.26 | |
| 3rd | 360 | multifloral | 111.97 | 3.89 | |
| 3rd | 360 | buckwheat | 97.24 | 3.58 | |
| 3rd | 365 | lime | 110.20 | 4.75 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilczyńska, A.; Żak, N. The Use of Fluorescence Spectrometry to Determine the Botanical Origin of Filtered Honeys. Molecules 2020, 25, 1350. https://doi.org/10.3390/molecules25061350
Wilczyńska A, Żak N. The Use of Fluorescence Spectrometry to Determine the Botanical Origin of Filtered Honeys. Molecules. 2020; 25(6):1350. https://doi.org/10.3390/molecules25061350
Chicago/Turabian StyleWilczyńska, Aleksandra, and Natalia Żak. 2020. "The Use of Fluorescence Spectrometry to Determine the Botanical Origin of Filtered Honeys" Molecules 25, no. 6: 1350. https://doi.org/10.3390/molecules25061350
APA StyleWilczyńska, A., & Żak, N. (2020). The Use of Fluorescence Spectrometry to Determine the Botanical Origin of Filtered Honeys. Molecules, 25(6), 1350. https://doi.org/10.3390/molecules25061350