Influence of Altitude on Phytochemical Composition of Hemp Inflorescence: A Metabolomic Approach





Abstract
:1. Introduction
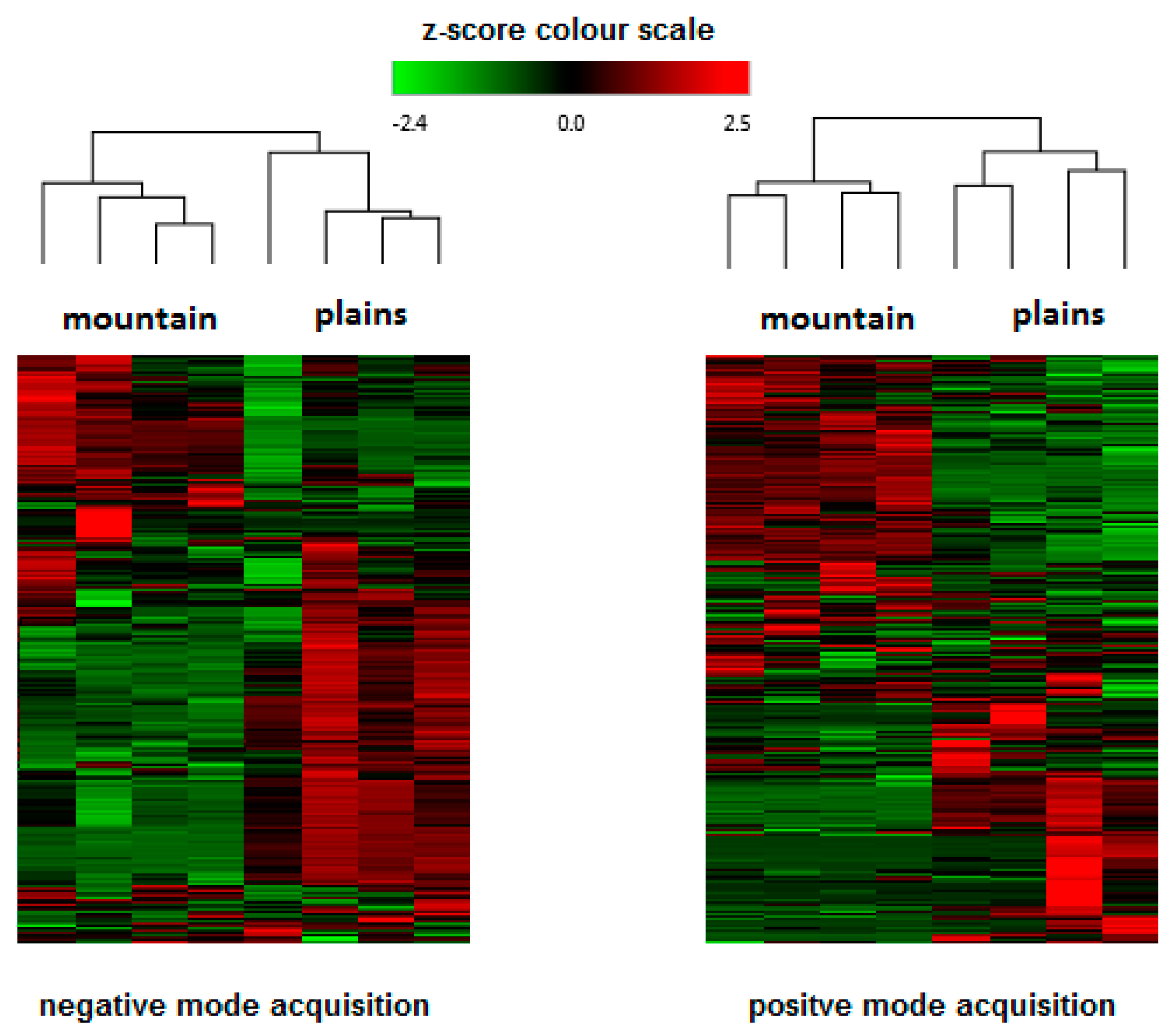
2. Results and Discussion
2.1. Terpenoids Profile Estimated by HS-SPME- GC-MS Analytical Procedure
2.2. Quantification of Cannabinoids by HPLC-Q-Exactive-Orbitrap®-MS Analysis
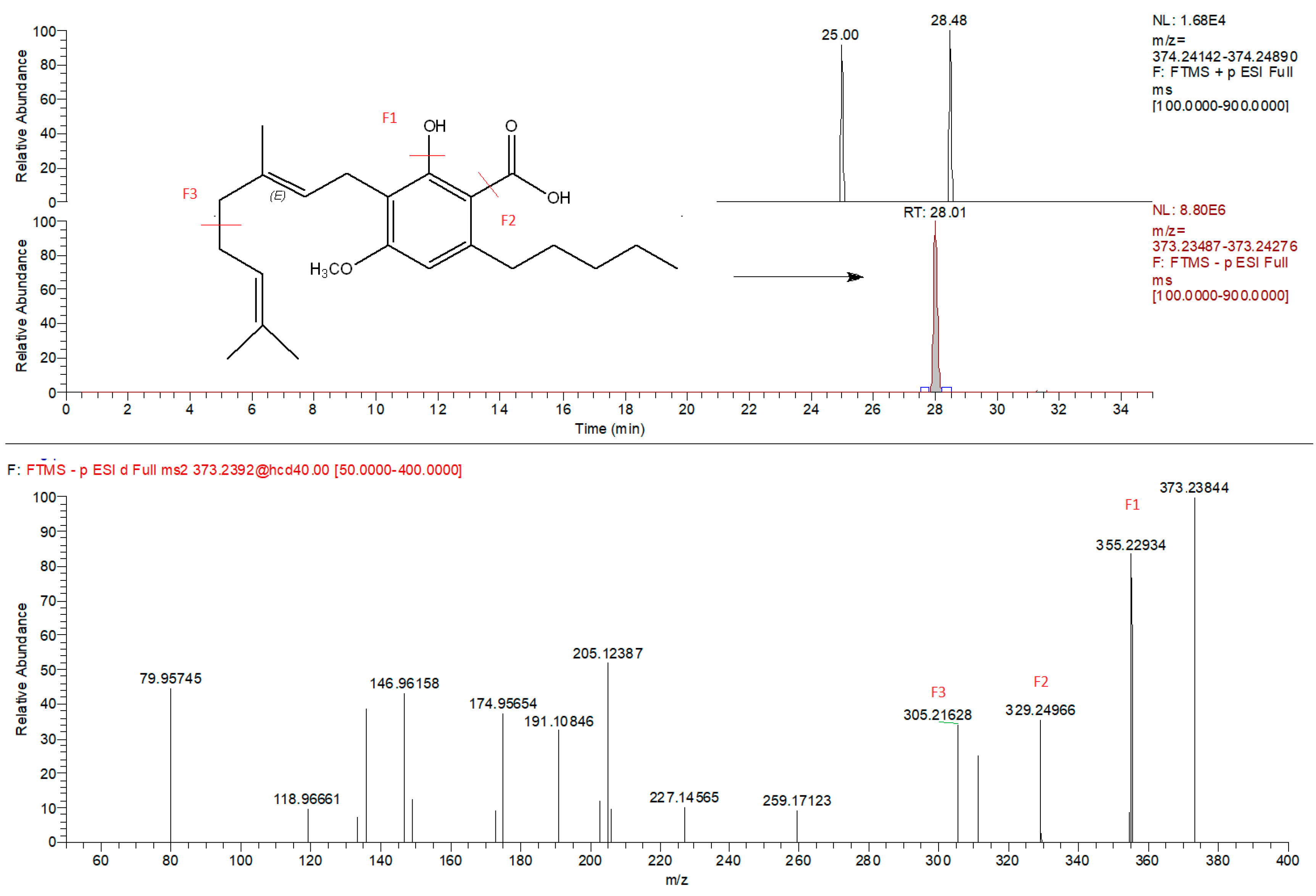
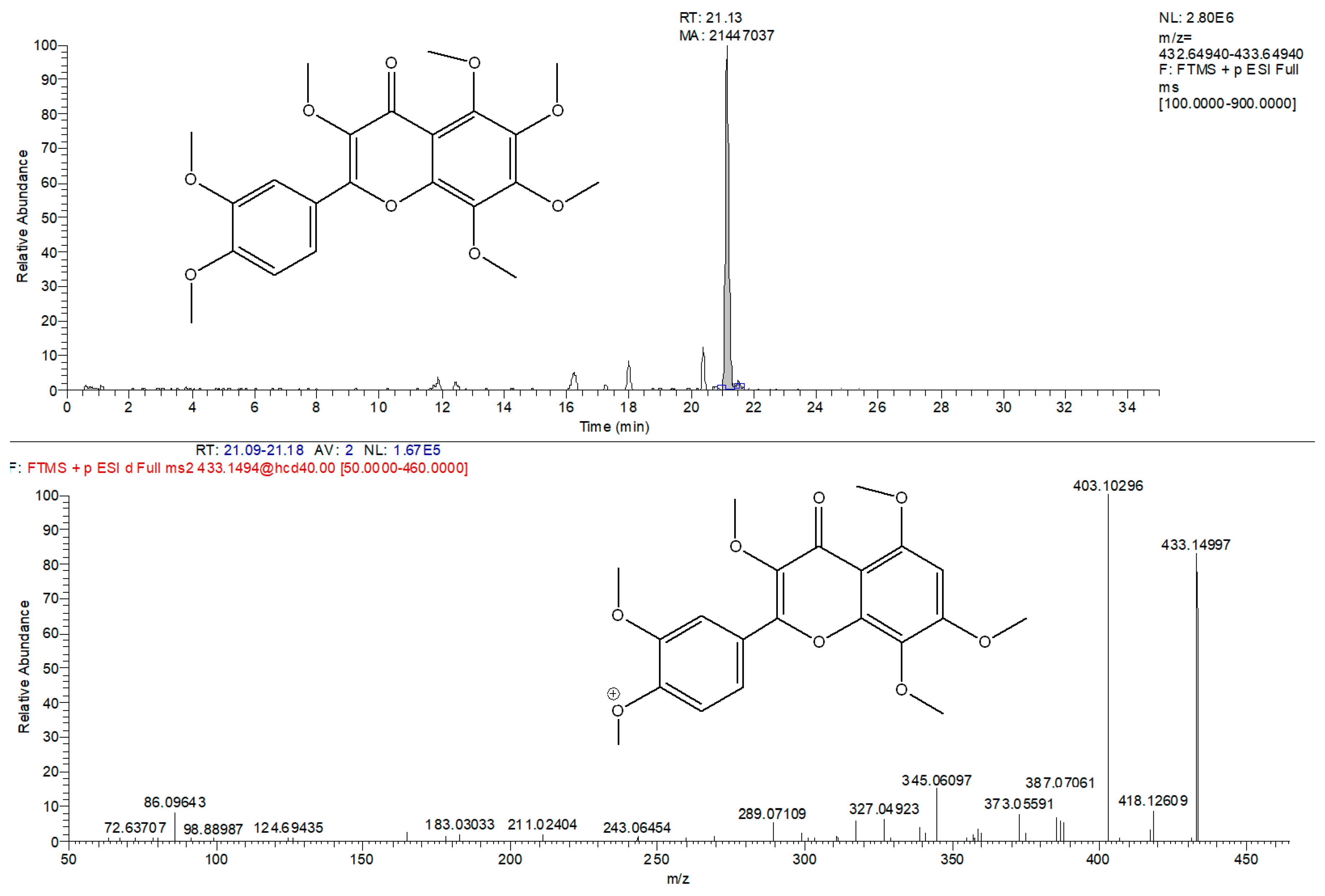
2.3. HPLC-Q-Exactive-Orbitrap®-MS Untargeted Metabolomics Approach: Phytocannabinoids Profiling and Identification of Polyphenolic Structures
3. Materials and Methods
3.1. Experimental Fields and Samples Collection
3.1.1. Clones
3.1.2. Plant Parts Sampled
3.2. Chemical and Reagents
3.3. Superfine Grinding (SFG) Sample Preparation
3.4. Accelerated Solvent Extraction (ASE) for Cannabinoids Profiling
3.5. Cannabinoids HPLC-Q-Exactive-Orbitrap-MS Evaluation
3.6. HPLC-Q-Exactive-Orbitrap-MS Untargeted Metabolomics Approach
3.7. HS-SPME and GC-MS Analysis for Terpenes Examination
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Regulation (EU) No 1307/2013 of the European Parliament and of the Council of 17 December 2013 Establishing Rules for Direct Payments to Farmers under Support Schemes within the Framework of the Common Agricultural Policy and Repealing Council Regulation (EC) No 637/2008 and Council Regulation (EC) No 73/2009. Available online: https://eur-lex.europa.eu/ (accessed on 27 November 2019).
- LEGGE 2 dicembre 2016, n. 242. Disposizioni per la promozione della coltivazione e della filiera agroindustriale della canapa. (16G00258). Available online: https://www.gazzettaufficiale.it/eli/gu/2016/12/30/304/sg/pdf (accessed on 27 November 2019).
- Salentijn, E.M.J.; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L. New developments in fiber hemp (Cannabis sativa L.) breeding. Ind. Crops Prod 2015, 68, 32–41. [Google Scholar] [CrossRef]
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key cultivation techniques for hemp in Europe and China. Ind. Crops Prod 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Plant Variety Catalogues, Databases & Information Systems. Available online: https://ec.europa.eu/food/plant/plant_propagation_material/plant_variety_catalogues_databases_en (accessed on 18 January 2019).
- de Meijer, E.P.M. Fibre hemp cultivars: A survey of origin, ancestry, availability and brief agronomic characteristics. J. Int. Hemp Assoc. 1995, 2, 66–73. [Google Scholar]
- ElSohly, M.; Radwan, M.; Gul, W.; Chandra, S.; Galal, A. Phytochemistry of Cannabis sativa. In Phytocannabinoids: Progress in the Chemistry of Organic Natural Products 103; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J., Eds.; Springer: Basel, Switzerland, 2017. [Google Scholar]
- Hanuš, L.O.; Meyer, S.M.; Muñoz, E.; Taglialatela-Scafati, O.; Appendino, G. Phytocannabinoids: A unified critical inventory. Nat. Prod. Rep. 2016, 33, 1357–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citti, C.; Braghiroli, D.M.; Vandelli, A.; Cannazza, G. Pharmaceutical and biomedical analysis of cannabinoids: A critical review. J. Pharm. Biomed. Anal. 2018, 147, 566–579. [Google Scholar] [CrossRef]
- Whiting, P.F.; Wolff, R.F.; Deshpande, S.; Di Nisio, M.; Duffy, S.; Hernandez, A.V.; Keurentjes, J.C.; Lang, S.; Misso, K.; Ryder, S.; et al. Cannabinoids for medical use: A systematic review and meta-analysis. JAMA 2015, 313, 2456–2473. [Google Scholar] [CrossRef]
- Pavlovic, R.; Nenna, G.; Calvi, L.; Panseri, S.; Borgonovo, G.; Giupponi, L.; Cannazza, G.; Giorgi, A. Quality traits of “cannabidiol oils”: Cannabinoids content, terpene fingerprint and oxidation stability of european commercially available preparations. Molecules 2018, 23, 1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlovic, R.; Panseri, S.; Giupponi, L.; Leoni, V.; Citti, C.; Cattaneo, C.; Cavaletto, M.; Giorgi, A. Phytochemical and Ecological Analysis of Two Varieties of Hemp (Cannabis sativa L.) Grown in a Mountain Environment of Italian Alps. Front. Plant Sci. 2019, 10, 1265. [Google Scholar] [CrossRef]
- Brenneisen, R. Chemistry and Analysis of Phytocannabinoids and Other Cannabis Constituents. In Marijuana and the Cannabinoids; ElSohly, M.A., Ed.; Humana Press: Clifton, NJ, USA, 2007; pp. 17–49. [Google Scholar]
- Pollastro, F.; Minassi, A.; Fresu, L.G. Cannabis Phenolics and their Bioactivities. Curr. Med. Chem. 2018, 25, 1160–1185. [Google Scholar] [CrossRef]
- Calzolari, D.; Magagnini, G.; Lucini, L.; Grassi, G.; Appendino, G.B.; Amaducci, S. High added-value compounds from Cannabis threshing residues. Ind. Crop. Prod. 2017, 108, 558–563. [Google Scholar] [CrossRef]
- Frassinetti, S.; Moccia, E.; Caltavuturo, L.; Gabriele, M.; Longo, V.; Bellani, L.; Giorgi, G.; Giorgetti, L. Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food Chem. 2018, 262, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B. The Case for the Entourage Effect and Conventional Breeding of Clinical Cannabis: No “Strain,” No Gain. Front. Plant Sci. 2019, 9, 1969. [Google Scholar]
- Elzinga, S.; Fischedick, R.; Podkolinski, J.; Raber, C. Cannabinoids and terpenes as chemotaxonomic markers in cannabis. Nat. Prod. Chem. Res. 2015, 3, 81. [Google Scholar]
- Aizpurua-Olaizola, O.; Soydaner, U.; Öztürk, E.; Schibano, D.; Simsir, Y.; Navarro, P.; Etxebarria, N.; Usobiaga, A. Evolution of the Cannabinoid and Terpene Content during the Growth of Cannabis sativa Plants from Different Chemotypes. J. Nat. Prod. 2016, 79, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.A.; Russo, E.B.; Smith, K.M. Pharmacological Foundations of Cannabis Chemovars. Planta Med. 2018, 84, 225–233. [Google Scholar] [CrossRef]
- Bertoli, A.; Tozzi, S.; Pistelli, L.; Angelini, L.G. Fibre hemp inflorescences: From crop-residues to essential oil production. Ind. Crop. Prod. 2010, 32, 329–337. [Google Scholar] [CrossRef]
- Schluttenhofer, C.; Yuan, L. Challenges towards Revitalizing Hemp: A Multifaceted Crop. Trends Plant Sci. 2017, 11, 917–929. [Google Scholar] [CrossRef] [Green Version]
- Calvi, L.; Pentimalli, D.; Panseri, S.; Giupponi, L.; Gelmini, F.; Beretta, G.; Vitali, D.; Bruno, M.; Zilio, E.; Pavlovic, R.; et al. Comprehensive quality evaluation of medical Cannabis sativa L. inflorescence and macerated oils based on HS-SPME coupled to GC–MS and LC-HRMS (q-exactive orbitrap®) approach. J. Pharm. Biomed. Anal. 2018, 150, 208–219. [Google Scholar] [CrossRef]
- Calvi, L.; Pavlovic, R.; Panseri, S.; Giupponi, L.; Leoni, V.; Giorgi, A. Quality traits of Medical Cannabis sativa L. inflorescences and derived products based on comprehensive analytical investigation. In Recent Advcance in Cannabinoid Research; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Giupponi, L.; Pentimalli, D.; Manzo, A.; Panseri, S.; Giorgi, A. Effectiveness of fine root fingerprinting as a tool to identify plants of the Alps: Results of preliminary study. Plant Biosyst. 2017, 152, 464–473. [Google Scholar] [CrossRef]
- Giorgi, A.; Manzo, A.; Nanayakkara, N.N.; Giupponi, L.; Cocucci, M.; Panseri, S. Effect of biotic and abiotic stresses on volatile emission of Achillea collina Becker ex Rchb. Nat. Prod. Res. 2015, 29, 1695–1702. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, R.; Borgonovo, G.; Leoni, V.; Giupponi, L.; Ceciliani, G.; Sala, S.; Bassoli, A.; Giorgi, A. Effectiveness of Different Analytical Methods for the Characterization of Propolis: A Case of Study in Northern Italy. Molecules 2020, 25, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namdar, D.; Moran, M.; Ion, A.; Koltai, H. Variation in the compositions of cannabinoid and terpenoids in Cannabis sativa derived from inflorescence position along the stem and extraction methods. Ind. Crops Prod. 2018, 113, 376–382. [Google Scholar] [CrossRef]
- Ascrizzi, R.; Ceccarini, L.; Tavarini, S.; Flaminia, G.; Angelini, L.G. Valorisation of hemp inflorescence after seed harvest: Cultivation site and harvest time influence agronomic characteristics and essential oil yield and composition. Ind. Crops Prod. 2019, 139, 111541. [Google Scholar] [CrossRef]
- Marchini, L.M.; Charvoz, C.; Dujourdy, L.; Baldovini, N.; Filippi, J.J. Multidimensional analysis of cannabis volatile constituents: Identification of 5,5-dimethyl-1-vinylbicyclo [2 .1.1] hexane as a volatile marker of hashish, the resin of Cannabis Sativa. J. Chromatogr. A 2014, 1370, 200–215. [Google Scholar] [CrossRef]
- Glivar, T.; Eržen, J.; Kreft, S.; Zagožen, M.; Čerenak, A.; Čeh, B.; Benković, E.T. Cannabinoid content in industrial hemp (Cannabis sativa L.) varieties grown in Slovenia. Ind. Crops Prod. 2020, 145, 112082. [Google Scholar] [CrossRef]
- Zengin, G.; Menghini, L.; Di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A.; Angiolella, L.; et al. Chromatographic analyses, in vitro biological activities, and cytotoxicity of Cannabis sativa L. essential oil: A multidisciplinary study. Molecules 2018, 23, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharbach, M.; Marmouzic, I.; El Jemlid, M.; Bouklouzeb, A.; Heydena, Y.V. Recent advances in untargeted and targeted approaches applied in herbal-extracts and essential-oils fingerprinting—A review. J. Pharm. Biomed. Anal. 2020, 177, 112849. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef]
- Cimaglia, P.; Vieceli Dalla Sega, F.; Vitali, F.; Lodolini, V.; Bernucci, D.; Passarini, G.; Fortini, F.; Marracino, L.; Aquila, G.; Rizzo, P.; et al. Effectiveness of a Novel Nutraceutical Compound Containing Red Yeast Rice, Polymethoxyflavones and Antioxidants in the Modulation of Cholesterol Levels in Subjects With Hypercholesterolemia and Low-Moderate Cardiovascular Risk: The NIRVANA Study. Front. Physiol. 2019, 10, 217. [Google Scholar] [CrossRef]
- Raskin, I.; Turner, I.; Melander, W.R. Regulation of Heat Production in the Inflorescences of an Arum Lily by Endogenous Salicylic Acid. PNAS 1989, 86, 2214–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 1991, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansouri, H.; Asrar, Z. Effects of abscisic acid on content and biosynthesis of terpenoids in Cannabis sativa at vegetative stage. Biol. Plant. 2012, 56, 153–156. [Google Scholar] [CrossRef]
- Blasi, C.; Capotorti, G.; Copiz, R.; Guida, D.; Mollo, B.; Smiraglia, D.; Zavattero, L. Classification and mapping of the ecoregions of Italy. Plant Biosyst. 2014, 148, 1255–1345. [Google Scholar] [CrossRef]
- Mediavilla, V.; Jonquera, M.; Schmid-Slembrouck, I.; Soldati, A. Decimal code for growth stages of hemp (Cannabis sativa L.). J. Int. Hemp Assoc. 1998, 5, 68–74. [Google Scholar]
- Hillig, K.W. A chemotaxonomic analysis of terpenoid variation in Cannabis. Biochem. Syst. Ecol. 2004, 32, 875–891. [Google Scholar] [CrossRef]
- Li, L.; Li, R.; Zhou, J.; Zuniga, A.; Stanislaus, A.E.; Wu, Y.; Huan, T.; Zheng, J.; Shi, Y.; Wishart, D.S.; et al. MyCompoundID: Using an evidence-based metabolome library for metabolite identification. Anal. Chem. 2013, 85, 3401–3408. [Google Scholar] [CrossRef]
- Berman, P.; Futoran, K.; Lewitus, G.M.; Mukha, D.; Benami, M.; Shlomi, T.; Meiri, D. A new ESI-LC/MS approach for comprehensive metabolic profiling of phytocannabinoids in Cannabis. Sci. Rep. 2018, 8, 14280. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, A.; Panseri, S.; Mattara, M.S.; Andreis, C.; Chiesa, L.M. Secondary metabolites and antioxidant capacities of waldheimia glabra (decne.) regel from Nepal. J. Sci. Food Agric. 2013, 93, 1026–1034. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| MONO/DI/TRI Terpenes | M1 | P1 | p Value b | M2 | P2 | p Value | M3 | P3 | p Value | M4 | P4 | p Value | ||||||||
| mean a | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | |||||
| α-Pinene | 3823.9 | 81.1 | 2705.8 | 152.3 | 0.014 | 2401.3 | 56.3 | 4659.2 | 394.3 | 0.013 | 4870.9 | 3.7 | 1944.8 | 65.0 | <0.001 | 1788.6 | 190.1 | 1560.3 | 89.1 | n.s. |
| α-Fenchene | n.d. | n.d. | 26.3 | 3.0 | <0.001 | 44.4 | 27.6 | n.d. | n.d. | <0.001 | n.d. | n.d. | 13.1 | 1.1 | <0.001 | n.d. | n.d. | 12.4 | 2.6 | <0.001 |
| Camphene | 202.6 | 4.7 | 139.1 | 5.2 | 0.008 | 991.6 | 6.8 | 270.2 | 10.3 | <0.001 | 292.5 | 5.3 | 91.0 | 1.1 | <0.001 | 162.0 | 20.1 | 121.6 | 0.2 | 0.004 |
| β-Pinene | 2074.0 | 152.8 | 2067.3 | 149.8 | 0.050 | 12965.6 | 55.6 | 2979.8 | 233.7 | <0.001 | 2258.6 | 20.7 | 1058.5 | 94.8 | 0.0015 | 1456.0 | 336.7 | 1230.8 | 3.0 | n.s. |
| β-Myrcene | 26,294.5 | 450.3 | 23745.1 | 1070.4 | 0.020 | 76,993.1 | 4716.5 | 16,893.5 | 519.5 | 0.0008 | 23,597.9 | 716.7 | 5949.9 | 24.1 | 0.0002 | 26,723.6 | 2988.9 | 11,660.5 | 1778.3 | 0.004 |
| Limonene | 2603.1 | 112.5 | 3023.2 | 150.3 | n.s. | 11472.7 | 276.0 | 4184.1 | 134.1 | <0.001 | 5565.0 | 846.0 | 962.2 | 1.4 | 0.011 | 5538.9 | 93.8 | 3883.5 | 8.0 | <0.001 |
| β-Phellandrene | 642.8 | 6.2 | 628.5 | 40.6 | n.s. | 2017.8 | 10.1 | 515.9 | 34.6 | 0.0012 | 662.0 | 348.3 | 188.5 | 2.2 | 0.0015 | 681.6 | 54.9 | 417.5 | 20.4 | 0.026 |
| Cis-ocimene | 352.70 | 1.6 | 321.5 | 60.3 | n.s. | 132.0 | 2.2 | 38.8 | 4.5 | <0.001 | 44.6 | 1.0 | 8.6 | 0.7 | <0.001 | 34.4 | 9.8 | 34.5 | 3.2 | n.s. |
| γ-Terpinene | 37.0 | 5.9 | 23.3 | 7.6 | 0.002 | 86.5 | 1.0 | 77.6 | 0.4 | 0.004 | 69.0 | 10.9 | 49.9 | 1.4 | 0.054 | 18.5 | 5.3 | 15.6 | 0.8 | n.s. |
| β-Ocimene | 6571.3 | 25.5 | 5555.3 | 40.1 | 0.001 | 135.20 | 5.1 | 138.0 | 2.0 | n.s. | 493.7 | 16.3 | 28.9 | 28.9 | <0.001 | 139.6 | 6.7 | 110.4 | 6.3 | 0.001 |
| α-Terpinolene | 136.4 | 15.4 | 143.1 | 26.3 | n.s. | 486.1 | 48.4 | 190.2 | 41.2 | 0.0002 | 274.3 | 7.7 | 45.4 | 0.9 | <0.001 | 192.0 | 6.5 | 144.6 | 10.3 | 0.002 |
| Terpene | 6.0 | 0.1 | 4.8 | 0.1 | 0.007 | 12.8 | 0.9 | 5.3 | 0.7 | <0.001 | 4.5 | 0.2 | 1.2 | 0.1 | <0.001 | 11.4 | 0.4 | 0.8 | 0.0 | <0.001 |
| α-Fenchone | 47.5 | 2.6 | 20.0 | 7.0 | 0.009 | 84.2 | 28.2 | 74.6 | 24.6 | 0.045 | 74.3 | 14.3 | 9.4 | 0.6 | 0.017 | 41.1 | 0.5 | 36.6 | 1.6 | 0.020 |
| Alloocimene | 80.9 | 0.1 | 91.7 | 17.1 | n.s. | 37.6 | 4.1 | 24.2 | 1.2 | 0.015 | 12.1 | 4.1 | 2.2 | 1.2 | <0.001 | 23.3 | 8.3 | 17.2 | 6.2 | 0.036 |
| Linalyl oxide | 19.6 | 0.7 | 16.2 | 2.1 | 0.070 | 8.5 | 0.1 | 7.2 | 5.3 | 0.040 | 18.7 | 1.2 | 17.4 | 1.7 | n.s. | 21.1 | 0.8 | 16.6 | 1.6 | n.s. |
| 4,8-Epoxy-p-menth-1-ene | 147.8 | 16.8 | 167.9 | 13.0 | 0.012 | n.d. | n.d. | n.d. | n.d. | - | n.d. | n.d. | n.d. | n.d. | - | n.d. | n.d. | n.d. | n.d. | - |
| Pinalol | 117.0 | 1.6 | 130.0 | 20.0 | n.s. | 271.2 | 2.1 | 269.6 | 11.7 | n.s. | 248.2 | 31.2 | 229.8 | 20.2 | 0.1 | 139.8 | 12.4 | 131.2 | 5.9 | 0.1 |
| β-Linalool | 149.6 | 7.7 | 163.0 | 49.5 | n.s. | 693.6 | 3.4 | 576.4 | 31.3 | 0.028 | 617.0 | 49.8 | 54.9 | 2.4 | <0.001 | 1118.5 | 94.2 | 880.7 | 8.6 | 0.010 |
| α-Fenchol | 68.0 | 43.0 | 29.0 | 10.0 | n.s. | 164.6 | 48.2 | 130.8 | 35.9 | n.s. | 368.7 | 36.6 | 12.2 | 2.3 | 0.0014 | 171.9 | 71.6 | 12.7 | 2.7 | 0.060 |
| Verbenol | 89.8 | 36.4 | 37.7 | 18.9 | 0.035 | 204.5 | 155.1 | 65.1 | 0.6 | n.s. | 212.1 | 159.9 | 2.4 | 0.5 | n.s. | 6.8 | 6.5 | 11.7 | 1.3 | n.s. |
| tot | 43,464.5 | 39,038.8 | 109,203.3 | 31,100.5 | 40,141.9 | 11,038.4 | 38,269.1 | 20,299.2 | ||||||||||||
| Sesquiterpenes | M1 | P1 | p Value | M2 | P2 | p Value | M3 | P3 | p Value | M4 | P4 | p Value | ||||||||
| mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | mean | ± SD | |||||
| α-Ylangene | 130.8 | 7.3 | 201 | 19 | 0.044 | 47.1 | 13.7 | 60.7 | 6.6 | 0.05 | 15.6 | 6.5 | 3.4 | 0.9 | 0.007 | 13.3 | 6.5 | 9.2 | 0.2 | n.s. |
| α-Copaene | 41.7 | 1.2 | 50.7 | 4 | n.s. | 21.9 | 0.1 | 48.8 | 14.6 | n.s. | 9.7 | 0.6 | 1.9 | 0.2 | 0.002 | 22.1 | 2.3 | 20.8 | 0.8 | 0.027 |
| Zingiberene | 62.6 | 6.4 | 106.8 | 10.5 | 0.02 | 68.3 | 6.8 | 41.7 | 11.2 | <0.001 | 26 | 0.6 | 35 | 12.3 | 0.081 | 122.7 | 11.7 | 111.8 | 4.9 | n.s. |
| Longicyclene | 314.2 | 80.9 | 417.5 | 59.9 | 0.09 | 44.2 | 0.7 | 25 | 17.2 | n.s. | 43 | 0.9 | 2.5 | 0.8 | <0.001 | 302.6 | 63.2 | 207.1 | 11.2 | 0.08 |
| α-Bergamotene | 976.9 | 58.1 | 1440.9 | 115.3 | n.s. | 831.8 | 310.9 | 381.8 | 49.2 | n.s. | 326.5 | 54.1 | 77.9 | 13.9 | 0.004 | 3140.1 | 204.7 | 2395.8 | 86.2 | 0.008 |
| Trans-Caryophyllene | 6487.1 | 113.7 | 3668.7 | 502 | 0.008 | 9206.9 | 144.9 | 3797.7 | 41.8 | 0.007 | 3345.5 | 456.2 | 829.2 | 28.2 | <0.001 | 7017.4 | 922.6 | 1684.6 | 38.2 | <0.001 |
| Aristolene | 55.8 | 3.6 | 105 | 9.8 | 0.005 | 18.4 | 0.7 | 28 | 10.3 | n.s. | 2.63 | 2.63 | 0.1 | 0.91 | n.s. | n.d. | 819 | 114.4 | 0 | <0.001 |
| Isoledene | 33.2 | 6.9 | 67.3 | 10.9 | 0.004 | 31.9 | 11.9 | 31.8 | 10 | n.s. | 8.4 | 4.8 | 2.69 | 1.8 | n.s. | 13.4 | 1.1 | 12.7 | 0.4 | n.s. |
| β-Santalene | 12.3 | 2.2 | 23.9 | 3.7 | 0.005 | 6.9 | 0.2 | 7 | 2.5 | n.s. | 4.1 | 3.15 | 0.38 | 0.21 | n.s. | 8.2 | 1.3 | 10.7 | 2.5 | n.s. |
| Aromadendrene | 152 | 40.2 | 76.3 | 9.3 | n.s. | 195.7 | 1.4 | 7.6 | 0.1 | <0.001 | 19.9 | 1.4 | 14.3 | 2.8 | n.s. | 22.4 | 1.4 | 2.8 | 0.3 | 0.001 |
| α-Humulene | 3190 | 15 | 2099 | 183.2 | 0.007 | 3107.1 | 177.4 | 1284.2 | 9.9 | 0.002 | 3206.1 | 204.5 | 306.2 | 15.8 | 0.014 | 3431.4 | 87.2 | 2660.4 | 100.2 | <0.001 |
| β-Farnesene | 51.9 | 41.3 | 1495.4 | 4.6 | <0.001 | 1105.7 | 14.3 | 249.6 | 59.6 | 0.002 | n.d. | n.d. | n.d. | n.d. | - | 1494.2 | 17.4 | 1244.8 | 13.4 | 0.006 |
| β-Selinene | 416.4 | 29.7 | 632.4 | 49.2 | 0.003 | 245.5 | 47.2 | 502.8 | 50.4 | <0.001 | 123.2 | 1.9 | 211.7 | 10.6 | 0.003 | 112.6 | 28.5 | 271.5 | 7.6 | 0.004 |
| α-Selinene | 260.8 | 11.9 | 379.9 | 46.2 | 0.027 | 61.8 | 6.4 | 145.4 | 35.1 | 0.034 | 4.9 | 0.4 | 23.3 | 6.1 | 0.033 | 74.5 | 4.2 | 71.2 | 8.7 | n.s. |
| β-Bisabolene | 739.6 | 30.4 | 293.7 | 156.9 | 0.05 | 1515.6 | 29.5 | 581.2 | 21.5 | <0.001 | 557 | 51.2 | 129.9 | 8.8 | 0.003 | 1278.2 | 8.9 | 1024.2 | 14.3 | 0.003 |
| α-Farnesene | 213.8 | 29.6 | 357.2 | 68.4 | 0.045 | 661.2 | 21.3 | 314.2 | 14.4 | 0.008 | 160.8 | 26.6 | 37.9 | 21.2 | 0.031 | 500.2 | 18.2 | 583.2 | 86.2 | n.s. |
| δ-Cadinene | 128.2 | 20.8 | 215.1 | 38.5 | n.s. | 63.5 | 4.2 | 74.2 | 4.6 | 0.013 | 124.4 | 0.2 | 23.9 | 3.7 | 0.008 | 66.8 | 5.2 | 33.4 | 20.2 | n.s. |
| β-Maaliene | 657.2 | 25.9 | 1048 | 224.2 | 0.008 | 297.8 | 12.4 | 197.2 | 10.2 | 0.0078 | 63.3 | 10.5 | 59.6 | 14.1 | n.s. | 354.2 | 25.1 | 362 | 2.7 | n.s. |
| Selina-3,7(11)-diene | 1821 | 134.4 | n.d. | n.d. | <0.001 | 1623.3 | 157.2 | n.d. | n.d. | <0.001 | 581.6 | 81.6 | 132.5 | 12.6 | 0.007 | 1196.2 | 25.2 | 992.3 | 102 | n.s. |
| Caryophyllene oxide | 59.2 | 6.3 | 63.2 | 32 | n.s. | 68.2 | 6.9 | 30.1 | 4.3 | 0.024 | 18.9 | 4.1 | 6 | 0.7 | 0.006 | 67.2 | 2.4 | 73.6 | 4.6 | n.s. |
| Guaiol | 114.5 | 20.9 | 144.8 | 48.2 | n.s. | 326.2 | 28.1 | 280.4 | 70.7 | n.s. | 214.4 | 56.6 | 337 | 20.2 | n.s. | 289.2 | 15.4 | 227.9 | 8.6 | n.s. |
| 10-Epi-γ-Eudesmol | 329.6 | 26.7 | 170.7 | 47.2 | 0.003 | 425.2 | 42.4 | 331.2 | 3.1 | 0.004 | 243.2 | 51.4 | 42.5 | 0.1 | 0.005 | 181.1 | 21 | 295.2 | 3.1 | n.s. |
| tot | 16,248.8 | 13,057.5 | 19,974.2 | 8420.6 | 9099.2 | 2277.9 | 19,708.0 | 12,409.6 |
| Mountain | Plains | Statistical Significance | |||
|---|---|---|---|---|---|
| Mean | SD (±) | Mean | SD (±) | ||
| Neutral forms | |||||
| CBD | 5300 | 3500 | 6000 | 3800 | ns |
| Δ9-THC | <LOQ | / | <LOQ | / | / |
| CBN | <LOQ | / | <LOQ | / | / |
| CBC | 460 | 120 | 120 | 50 | 0.005 |
| CBG | 110 | 10 | 180 | 80 | <0.001 |
| CBDV | 250 | 400 | 450 | 40 | <0.001 |
| Δ9-THCV | <LOQ | / | <LOQ | / | / |
| Acid forms | |||||
| CBDA | 99,600 | 24,800 | 68,220 | 15,000 | 0.01 |
| Δ9-THCA | 840 | 200 | 1010 | 400 | ns |
| CBNA | 40 | 4 | 50 | 10 | ns |
| CBCA | 1570 | 200 | 570 | 30 | 0.008 |
| CBGA | 7410 | 900 | 4510 | 400 | 0.015 |
| CBDVA | 310 | 70 | 240 | 20 | ns |
| Δ9- THCVA | <LOQ | / | <LOQ | / | / |
| Class | Compound | Formula | (M + H)+/Main Fragment | (M − H)−/Main Fragment | RegulationMountain vs. Pains |
|---|---|---|---|---|---|
| Phytocannabinoids | |||||
| CBG cannabigerol class | CBG | C21H32O2 | 317.2475/193.1223 | 315.2329/191.1078 | Upregulated in mountain |
| Sesqui-CBG | C26H40O2 | 385.3173/193.1223 | n.i. | ||
| 6,7-epoxy-CBG | C21H32O3 | 333.2424/315.1867 | n.i. | ||
| CBGVA | C20H27O4 | 333.2060/173.0962 | 331.1915/313.1809 | ||
| 6,7-epoxy-CBGA | C22H32O5 | 377.2323/341.2113 | 375.2185/257.3077 | ||
| CBGA | C22H31O4 | 361.2375/219.1017 | 359.2228/191.1078 | ||
| CBGMA | C23H34O4 | n.i. | 373.2384/355.2293 | ||
| Sesqui-CBGA | C27H40O4 | n.i. | 427.2854/409.2748 | ||
| CBD (cannabidiol) class | CBDV | C19H26O2 | 287.2006/165.0914 | 285.1860/217.1234 | Upregulated in mountain |
| Nor-CBD | C20H28O2 | 301.2162/179.1070 | 299.2017/231.1391 | ||
| CBD | C21H30O2 | 315.2319/193.1223 | 313.2173/191.1078 | ||
| CBDM | C22H32O2 | 329.2475/229.0812 | 327.2329/205.1234 | ||
| CBDVA | C20H26O4 | 331.1904/313.1801 | 329.1758/217.1123 | ||
| Nor-CBDA | C20H28O4 | 345.2060/327.1956 | 343.1915/231.1391 | ||
| CBDA | C22H30O4 | 359.2219/341.2114 | 357.2017/245.1547 | ||
| CBDMA | C32H46O4 | n.i. | 371.2228/259.1704 | ||
| Sesquiterpene-CBDA ester | C32H46O4 | 495.3469/341.2114 | 493.3323/357.2017 | ||
| γ-Eudesmyl-CBDA ester | C37H54O4 | 562.4017/341.2114 | 561.3949/357.2017 | ||
| Δ9-THC tetrahydrocannabinol class | THCV | C19H26O2 | 287.2006/165.0914 | 285.1860/217.1234 | ns |
| THC | C21H30O2 | 315.2319/193.1223 | 313.2173/n.i. | ||
| THCVA | C20H26O4 | 331.1904/313.1801 | 329.1758/189.0921 | ||
| THCA | C22H30O4 | 359.2219/341.2114 | 357.2071/245.1547 | ||
| CBC cannabichromene class | CBCV | C19H26O2 | 287.2006/165.0914 | 285.1860/163.0765 | Upregulated in mountain |
| CBC | C21H30O2 | 315.2319/193.1223 | 313.2173/n.d. | ||
| CBCVA | C20H26O4 | 331.1904/313.1801 | 329.1758/189.0921 | ||
| CBCA | C22H30O4 | 359.2219/341.2114 | 357.2071/313.2179 | ||
| CBN cannabinol class | CBN | C21H26O2 | 311.2007/223.1118 | 309.1860/n.i. | ns |
| CBNA | C22H26O4 | 355.1904 | 337.1800 | ||
| Cannaflavin A | C26H28O6 | 437.1964/313.0709 | 435.1813/309.0413 | ||
| Isoprenoid flavones | Cannaflavin B | C21H20O6 | 369.1333/313.0706 | 367.1195/309.0499 | Upregulated in mountain |
| Cannaflavin C | C26H28O6 | 437.1964/313.0709 | 435.1813/309.0414 | ||
| Polymethoxyflavones | 3-Methoxynobiletin | C22H24O9 | 433.14980/403.10296 | n.i. | Upregulated in mountain |
| Flavones | apigenin | C15H10O5 | 271.0601/nd | 269.0455/117.0348 | Upregulated in plains |
| Phenolic acid | Salicylic acid | C7H6O3 | n.i. | 137.0426/95.8554 | Upregulated in plains |
| Abscisic acid | C15H20O4 | n.i. | 263.1289/219.1391 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giupponi, L.; Leoni, V.; Pavlovic, R.; Giorgi, A. Influence of Altitude on Phytochemical Composition of Hemp Inflorescence: A Metabolomic Approach. Molecules 2020, 25, 1381. https://doi.org/10.3390/molecules25061381
Giupponi L, Leoni V, Pavlovic R, Giorgi A. Influence of Altitude on Phytochemical Composition of Hemp Inflorescence: A Metabolomic Approach. Molecules. 2020; 25(6):1381. https://doi.org/10.3390/molecules25061381
Chicago/Turabian StyleGiupponi, Luca, Valeria Leoni, Radmila Pavlovic, and Annamaria Giorgi. 2020. "Influence of Altitude on Phytochemical Composition of Hemp Inflorescence: A Metabolomic Approach" Molecules 25, no. 6: 1381. https://doi.org/10.3390/molecules25061381
APA StyleGiupponi, L., Leoni, V., Pavlovic, R., & Giorgi, A. (2020). Influence of Altitude on Phytochemical Composition of Hemp Inflorescence: A Metabolomic Approach. Molecules, 25(6), 1381. https://doi.org/10.3390/molecules25061381