
Korean Red Ginseng Plays an Anti-Aging Role by Modulating Expression of Aging-Related Genes and Immune Cell Subsets

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. KRG Inhibited Aging-Related Thymic Involution
2.2. KRG Altered Protein Expression, But Not mRNA Expression of Aging-Related Genes in Aged Mice
2.3. KRG Regulated the Population of Thymic and Splenic Immune Cells in Aged Mice
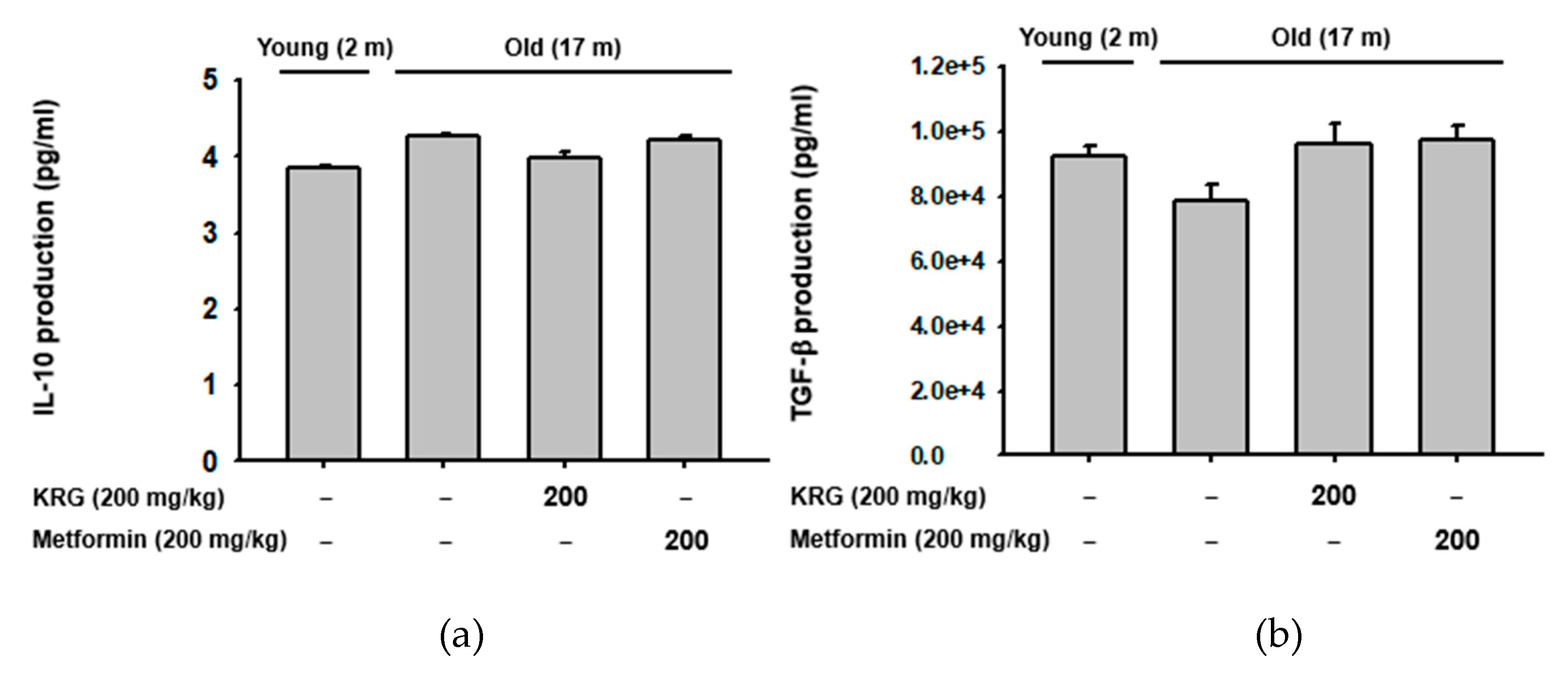
2.4. KRG Did Not Alter the Production of Regulatory T Cell-Specific Cytokines IL-10 and TGF-β in Aged Mice
2.5. KRG Increased the Expression of Aging-Related Genes in Con A-Stimulated T Cells in Aged Mice
3. Materials and Methods
3.1. Materials
3.2. Animal Husbandry and Experiments
3.3. Quantitative Real Time PCR
3.4. Western Blot Analysis
3.5. Flow Cytometry Analysis
3.6. ELISA
3.7. Cell Viability Assay
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Titorenko, V.I. Molecular and Cellular Mechanisms of Aging and Age-related Disorders. Int. J. Mol. Sci. 2018, 19, 2049. [Google Scholar] [CrossRef] [Green Version]
- Jayanthi, P.; Joshua, E.; Ranganathan, K. Ageing and its implications. J. Oral Maxillofac. Pathol. 2010, 14, 48–51. [Google Scholar] [CrossRef] [Green Version]
- Christensen, K.; Doblhammer, G.; Rau, R.; Vaupel, J.W. Ageing populations: The challenges ahead. Lancet 2009, 374, 1196–1208. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and Oxidative Stress in Human Diseases: From Molecular Mechanisms to Novel Treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minciullo, P.L.; Catalano, A.; Mandraffino, G.; Casciaro, M.; Crucitti, A.; Maltese, G.; Morabito, N.; Lasco, A.; Gangemi, S.; Basile, G. Inflammaging and Anti-Inflammaging: The Role of Cytokines in Extreme Longevity. Arch. Immunol. Ther. Exp. (Warsz) 2016, 64, 111–126. [Google Scholar] [CrossRef]
- Kurek, K.; Plitta-Michalak, B.; Ratajczak, E. Reactive Oxygen Species as Potential Drivers of the Seed Aging Process. Plants 2019, 8, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, M.T.; Casciaro, M.; Gangemi, S.; Buquicchio, R. Immunosenescence in aging: Between immune cells depletion and cytokines up-regulation. Clin. Mol. Allergy 2017, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidler, C.; Woycicki, R.; Ilnytskyy, Y.; Metz, G.; Kovalchuk, I.; Kovalchuk, O. Immunosenescence is associated with altered gene expression and epigenetic regulation in primary and secondary immune organs. Front. Genet. 2013, 4, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, B.K.; Kwon, S.W.; Park, J.H. Chemical diversity of ginseng saponins from Panax ginseng. J. Ginseng Res. 2015, 39, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.I.; Park, K.S.; Cho, I.H. Panax ginseng: A candidate herbal medicine for autoimmune disease. J. Ginseng Res. 2019, 43, 342–348. [Google Scholar] [CrossRef]
- Zhao, B.; Lv, C.; Lu, J. Natural occurring polysaccharides from Panax ginseng C. A. Meyer: A review of isolation, structures, and bioactivities. Int. J. Biol. Macromol. 2019, 133, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Bae, B.S.; Park, H.W.; Ahn, N.G.; Cho, B.G.; Cho, Y.L.; Kwak, Y.S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Qomaladewi, N.P.; Kim, M.Y.; Cho, J.Y. Autophagy and its regulation by ginseng components. J. Ginseng Res. 2019, 43, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.; Kim, J.H.; Kim, J.H.; Yi, Y.S.; Cho, J.Y. Functional role of ginseng-derived compounds in cancer. J. Ginseng Res. 2018, 42, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Mohanan, P.; Subramaniyam, S.; Mathiyalagan, R.; Yang, D.C. Molecular signaling of ginsenosides Rb1, Rg1, and Rg3 and their mode of actions. J. Ginseng Res. 2018, 42, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, H.Y.; Yu, M.; Yeom, E.; Lee, J.H.; Yoon, A.; Lee, K.S.; Min, K.J. Extension of Drosophila lifespan by Korean red ginseng through a mechanism dependent on dSir2 and insulin/IGF-1 signaling. Aging (Albany NY) 2019, 11, 9369–9387. [Google Scholar] [CrossRef] [PubMed]
- Kopalli, S.R.; Cha, K.M.; Ryu, J.H.; Lee, S.H.; Jeong, M.S.; Hwang, S.Y.; Lee, Y.J.; Song, H.W.; Kim, S.N.; Kim, J.C.; et al. Korean red ginseng improves testicular ineffectiveness in aging rats by modulating spermatogenesis-related molecules. Exp. Gerontol. 2017, 90, 26–33. [Google Scholar] [CrossRef]
- Park, S.; Kim, C.S.; Min, J.; Lee, S.H.; Jung, Y.S. A high-fat diet increases oxidative renal injury and protein glycation in D-galactose-induced aging rats and its prevention by Korea red ginseng. J. Nutr. Sci. Vitaminol. (Tokyo) 2014, 60, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Kim, Y.J.; Lim, H.J.; Kim, Y.S.; Park, H.Y.; Choung, Y.H. Red ginseng delays age-related hearing and vestibular dysfunction in C57BL/6 mice. Exp. Gerontol. 2014, 57, 224–232. [Google Scholar] [CrossRef]
- El-Naseery, N.I.; Mousa, H.S.E.; Noreldin, A.E.; El-Far, A.H.; Elewa, Y.H.A. Aging-associated immunosenescence via alterations in splenic immune cell populations in rat. Life Sci. 2020, 241, 117168. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Immunosenescence: The potential role of myeloid-derived suppressor cells (MDSC) in age-related immune deficiency. Cell. Mol. Life Sci. 2019, 76, 1901–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischof, J.; Gartner, F.; Zeiser, K.; Kunz, R.; Schreiner, C.; Hoffer, E.; Burster, T.; Knippschild, U.; Zimecki, M. Immune Cells and Immunosenescence. Folia Biol. (Praha) 2019, 65, 53–63. [Google Scholar] [PubMed]
- Palmer, S.; Albergante, L.; Blackburn, C.C.; Newman, T.J. Thymic involution and rising disease incidence with age. Proc. Natl. Acad. Sci. USA 2018, 115, 1883–1888. [Google Scholar] [CrossRef] [Green Version]
- Odinokov, D.; Hamblin, M.R. Aging of lymphoid organs: Can photobiomodulation reverse age-associated thymic involution via stimulation of extrapineal melatonin synthesis and bone marrow stem cells? J. Biophotonics 2018, 11, e201700282. [Google Scholar] [CrossRef]
- Valencia, W.M.; Palacio, A.; Tamariz, L.; Florez, H. Metformin and ageing: Improving ageing outcomes beyond glycaemic control. Diabetologia 2017, 60, 1630–1638. [Google Scholar] [CrossRef] [Green Version]
- Glossmann, H.H.; Lutz, O.M.D. Metformin and Aging: A Review. Gerontology 2019, 65, 581–590. [Google Scholar] [CrossRef]
- Docherty, C.K.; Salt, I.P.; Mercer, J.R. Lin28A induces energetic switching to glycolytic metabolism in human embryonic kidney cells. Stem Cell Res. Ther. 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Loffredo, F.S.; Steinhauser, M.L.; Jay, S.M.; Gannon, J.; Pancoast, J.R.; Yalamanchi, P.; Sinha, M.; Dall’Osso, C.; Khong, D.; Shadrach, J.L.; et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell 2013, 153, 828–839. [Google Scholar] [CrossRef] [Green Version]
- Katsimpardi, L.; Litterman, N.K.; Schein, P.A.; Miller, C.M.; Loffredo, F.S.; Wojtkiewicz, G.R.; Chen, J.W.; Lee, R.T.; Wagers, A.J.; Rubin, L.L. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science 2014, 344, 630–634. [Google Scholar] [CrossRef] [Green Version]
- Sinha, M.; Jang, Y.C.; Oh, J.; Khong, D.; Wu, E.Y.; Manohar, R.; Miller, C.; Regalado, S.G.; Loffredo, F.S.; Pancoast, J.R.; et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science 2014, 344, 649–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Wei, X.; Wang, D.; Lv, Z.; Geng, X.; Li, P.; Lu, J.; Wang, K.; Wang, X.; Sun, J.; et al. Positive effects of the young systemic environment and high GDF-11 levels on chondrocyte proliferation and cartilage matrix synthesis in old mice. Arthritis Rheumatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, Y.; Liu, D.; Liu, F.; Li, X.; Pan, L.; Pang, Y.; Chen, D. Role of growth differentiation factor 11 in development, physiology and disease. Oncotarget 2017, 8, 81604–81616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russomanno, G.; Corbi, G.; Manzo, V.; Ferrara, N.; Rengo, G.; Puca, A.A.; Latte, S.; Carrizzo, A.; Calabrese, M.C.; Andriantsitohaina, R.; et al. The anti-ageing molecule sirt1 mediates beneficial effects of cardiac rehabilitation. Immun. Ageing 2017, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, X.; Zhu, W.; Hu, X.; Jiang, Z.; Xu, Y.; Wang, L.; Zhou, Y.; Chen, P.; Zhang, N.; et al. SIRT1 ameliorates age-related senescence of mesenchymal stem cells via modulating telomere shelterin. Front. Aging Neurosci. 2014, 6, 103. [Google Scholar] [CrossRef] [Green Version]
- Waters, R.S.; Perry, J.S.A.; Han, S.; Bielekova, B.; Gedeon, T. The effects of interleukin-2 on immune response regulation. Math. Med. Biol. 2018, 35, 79–119. [Google Scholar] [CrossRef]
- Ross, S.H.; Cantrell, D.A. Signaling and Function of Interleukin-2 in T Lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [Google Scholar] [CrossRef]
- Chinen, T.; Kannan, A.K.; Levine, A.G.; Fan, X.; Klein, U.; Zheng, Y.; Gasteiger, G.; Feng, Y.; Fontenot, J.D.; Rudensky, A.Y. An essential role for the IL-2 receptor in Treg cell function. Nat. Immunol. 2016, 17, 1322–1333. [Google Scholar] [CrossRef]
- Fan, M.Y.; Low, J.S.; Tanimine, N.; Finn, K.K.; Priyadharshini, B.; Germana, S.K.; Kaech, S.M.; Turka, L.A. Differential Roles of IL-2 Signaling in Developing versus Mature Tregs. Cell Rep. 2018, 25, 1204–1213. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q. Therapeutic window of interleukin-2 for autoimmune diseases. Diabetes 2015, 64, 1912–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Tang, Q. Manipulating IL-2 and IL-2R in autoimmune diseases and transplantation. Immunotherapy 2015, 7, 1231–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenobia, C.; Hajishengallis, G. Basic biology and role of interleukin-17 in immunity and inflammation. Periodontol 2000 2015, 69, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Mysliwska, J.; Bryl, E.; Foerster, J.; Mysliwski, A. Increase of interleukin 6 and decrease of interleukin 2 production during the ageing process are influenced by the health status. Mech. Ageing Dev. 1998, 100, 313–328. [Google Scholar] [CrossRef]
- Ouyang, X.; Yang, Z.; Zhang, R.; Arnaboldi, P.; Lu, G.; Li, Q.; Wang, W.; Zhang, B.; Cui, M.; Zhang, H.; et al. Potentiation of Th17 cytokines in aging process contributes to the development of colitis. Cell. Immunol. 2011, 266, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Gillis, S.; Kozak, R.; Durante, M.; Weksler, M.E. Immunological studies of aging. Decreased production of and response to T cell growth factor by lymphocytes from aged humans. J. Clin. Investig. 1981, 67, 937–942. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Yao, D.; Zeng, X.; Kasakovski, D.; Zhang, Y.; Chen, S.; Zha, X.; Li, Y.; Xu, L. Age related human T cell subset evolution and senescence. Immun. Ageing 2019, 16, 24. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Jing, X.; Qiu, Z.; Cao, W.; Jiao, Y.; Routy, J.P.; Li, T. Aging of immune system: Immune signature from peripheral blood lymphocyte subsets in 1068 healthy adults. Aging (Albany NY) 2016, 8, 848–859. [Google Scholar] [CrossRef] [Green Version]
- Gounder, S.S.; Abdullah, B.J.J.; Radzuanb, N.; Zain, F.; Sait, N.B.M.; Chua, C.; Subramani, B. Effect of Aging on NK Cell Population and Their Proliferation at Ex Vivo Culture Condition. Anal. Cell. Pathol. (Amst) 2018, 2018, 7871814. [Google Scholar] [CrossRef] [Green Version]
- Solana, R.; Campos, C.; Pera, A.; Tarazona, R. Shaping of NK cell subsets by aging. Curr. Opin. Immunol. 2014, 29, 56–61. [Google Scholar] [CrossRef]
- Komai, T.; Inoue, M.; Okamura, T.; Morita, K.; Iwasaki, Y.; Sumitomo, S.; Shoda, H.; Yamamoto, K.; Fujio, K. Transforming Growth Factor-beta and Interleukin-10 Synergistically Regulate Humoral Immunity via Modulating Metabolic Signals. Front. Immunol. 2018, 9, 1364. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Santner-Nanan, B.; Hu, M.; Skarratt, K.; Lee, C.H.; Stormon, M.; Wong, M.; Fuller, S.J.; Nanan, R. IL-10 Potentiates Differentiation of Human Induced Regulatory T Cells via STAT3 and Foxo1. J. Immunol. 2015, 195, 3665–3674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Moreno, P.L.; Tripathi, S.; Chandraker, A. Regulatory T Cells and Kidney Transplantation. Clin. J. Am. Soc. Nephrol. 2018, 13, 1760–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Yamada, T.; Miller, R.A. Pgp-1hi T lymphocytes accumulate with age in mice and respond poorly to concanavalin A. Eur. J. Immunol. 1989, 19, 977–982. [Google Scholar] [CrossRef]
- Lefebvre, J.S.; Haynes, L. Aging of the CD4 T Cell Compartment. Open Longev. Sci. 2012, 6, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Baek, K.S.; Yi, Y.S.; Son, Y.J.; Yoo, S.; Sung, N.Y.; Kim, Y.; Hong, S.; Aravinthan, A.; Kim, J.H.; Cho, J.Y. In vitro and in vivo anti-inflammatory activities of Korean Red Ginseng-derived components. J. Ginseng Res. 2016, 40, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Rhee, M.H.; Lee, J.; Kim, S.H.; Yang, Y.; Kim, H.G.; Kim, Y.; Kim, C.; Kwak, Y.S.; Kim, J.H.; et al. Ginsenoside Rc from Korean Red Ginseng (Panax ginseng C.A. Meyer) Attenuates Inflammatory Symptoms of Gastritis, Hepatitis and Arthritis. Am. J. Chin. Med. 2016, 44, 595–615. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, W.S.; Yu, T.; Sung, G.H.; Park, K.W.; Yoon, K.; Son, Y.J.; Hwang, H.; Kwak, Y.S.; Lee, C.M.; et al. ATF-2/CREB/IRF-3-targeted anti-inflammatory activity of Korean red ginseng water extract. J. Ethnopharmacol. 2014, 154, 218–228. [Google Scholar] [CrossRef]
- Anisimov, V.N. Metformin: Do we finally have an anti-aging drug? Cell Cycle 2013, 12, 3483–3489. [Google Scholar] [CrossRef] [Green Version]
- Kinaan, M.; Ding, H.; Triggle, C.R. Metformin: An Old Drug for the Treatment of Diabetes but a New Drug for the Protection of the Endothelium. Med. Princ. Pract. 2015, 24, 401–415. [Google Scholar] [CrossRef]
- Xu, Q.; Ming, Z.; Dart, A.M.; Du, X.J. Optimizing dosage of ketamine and xylazine in murine echocardiography. Clin. Exp. Pharmacol. Physiol. 2007, 34, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Yi, Y.S.; Jeong, S.G.; Hong, Y.H.; Choi, K.J.; Hossain, M.A.; Hwang, H.; Rho, H.S.; Lee, J.; Kim, J.H.; et al. Ethanol Extract of Lilium Bulbs Plays an Anti-Inflammatory Role by Targeting the IKK[Formula: See text]/[Formula: See text]-Mediated NF-[Formula: See text]B Pathway in Macrophages. Am. J. Chin. Med. 2018, 46, 1281–1296. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Lorz, L.R.; Yi, D.K.; Noh, J.K.; Yi, Y.S.; Cho, J.Y. Viburnum pichinchense methanol extract exerts anti-inflammatory effects via targeting the NF-kappaB and caspase-11 non-canonical inflammasome pathways in macrophages. J. Ethnopharmacol. 2019, 245, 112161. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds, Korean red ginseng, are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Amount |
|---|---|
| Ginsenoside Rg1 | 5.5 mg |
| Ginsenoside Rb1 | |
| Ginsenoside Rg3 | |
| Carbohydrate | 0.33 g |
| Targets | Sequences (5′→3′) | |
|---|---|---|
| Lin28a | For | TCGGTGTCCAACCAGCAGTT |
| Rev | GGCGGTCATAGACAGGAAGC | |
| GDF-11 | For | GATCCTGGACCTACACGACTTC |
| Rev | GGCCTTCAGTACCTTTGTGAAC | |
| Sirt1 | For | CAGTGTCATGGTTCCTTTGC |
| Rev | CACCGAGGAACTACCTGAT | |
| IL-2 | For | TTAGGACAGCACAAAGTAAGCG |
| Rev | TGAGCTGATGTTAGCTCCCTG | |
| IL-17 | For | GCTGACCCCTAAGAAACCCC |
| Rev | GAAGCAGTTTGGGACCCCTT | |
| GAPDH | For | CAATGAATACGGCTACAGCAAC |
| Rev | AGGGAGATGCTCAGTGTTGG | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, K.K.; Yi, Y.-S.; Kim, J.K.; Kim, H.; Hossain, M.A.; Kim, J.-H.; Cho, J.Y. Korean Red Ginseng Plays an Anti-Aging Role by Modulating Expression of Aging-Related Genes and Immune Cell Subsets. Molecules 2020, 25, 1492. https://doi.org/10.3390/molecules25071492
Shin KK, Yi Y-S, Kim JK, Kim H, Hossain MA, Kim J-H, Cho JY. Korean Red Ginseng Plays an Anti-Aging Role by Modulating Expression of Aging-Related Genes and Immune Cell Subsets. Molecules. 2020; 25(7):1492. https://doi.org/10.3390/molecules25071492
Chicago/Turabian StyleShin, Kun Kuk, Young-Su Yi, Jin Kyeong Kim, Haeyeop Kim, Mohammad Amjad Hossain, Jong-Hoon Kim, and Jae Youl Cho. 2020. "Korean Red Ginseng Plays an Anti-Aging Role by Modulating Expression of Aging-Related Genes and Immune Cell Subsets" Molecules 25, no. 7: 1492. https://doi.org/10.3390/molecules25071492
APA StyleShin, K. K., Yi, Y. -S., Kim, J. K., Kim, H., Hossain, M. A., Kim, J. -H., & Cho, J. Y. (2020). Korean Red Ginseng Plays an Anti-Aging Role by Modulating Expression of Aging-Related Genes and Immune Cell Subsets. Molecules, 25(7), 1492. https://doi.org/10.3390/molecules25071492