
Influence of the Phenological State of in the Antioxidant Potential and Chemical Composition of Ageratina havanensis. Effects on the P-Glycoprotein Function


 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. P-gp Modulation by Extracts Obtained from Ageratina havanensis
2.2. Antioxidant Effects of Extracts Obtained from Ageratina havanensis
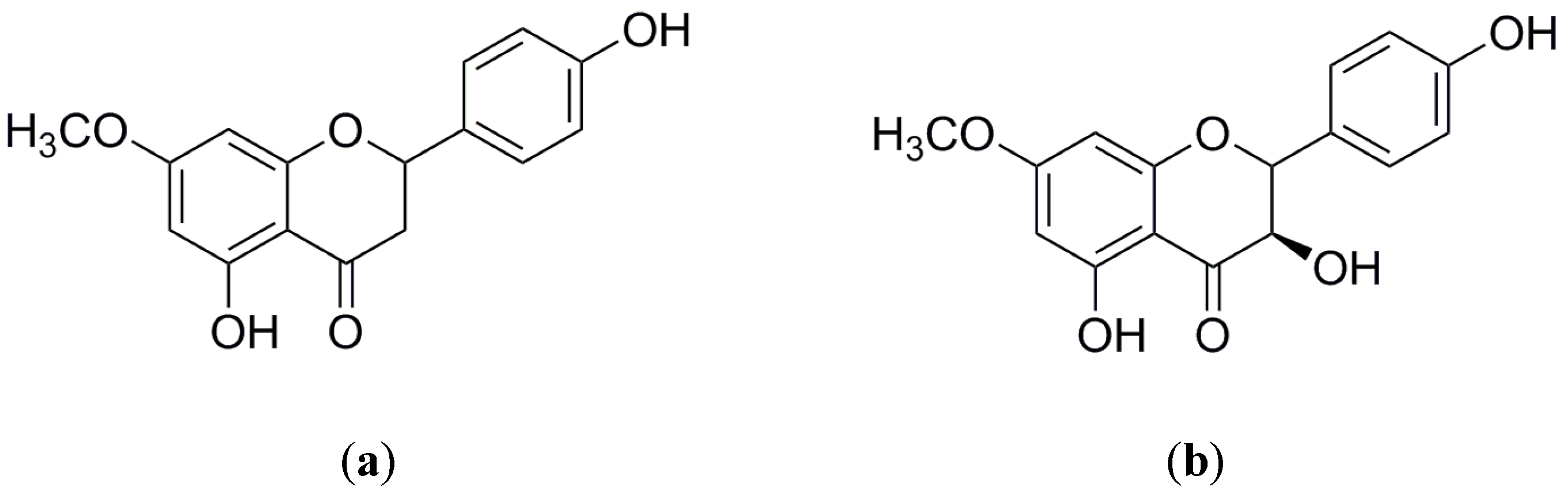
2.3. Quantification of Sakuranetin and 7-Methoxyromadendrin in the Extracts of Ageratina havanensis
2.3.1. Method Validation
Linearity, LDQ and LOQ
Matrix Effect and Recovery
Precision and Accuracy of the System
2.3.2. Concentration of Standards in the Extracts
2.3.3. Principal Component Analysis of Extracts
3. Discussion
4. Materials and Methods
4.1. General
4.2. Plant Material and Extraction
4.3. Cytotoxicity and P-glycoprotein Activity Assays
4.4. Antioxidant Capacity Assays
4.5. Statistical Analysis
4.6. Sample Preparation for UPLC-ESI-MS/MS Analysis
Standard Solutions Preparation for Quantification Analysis
4.7. UPLC-ESI-MS Analysis of the Extracts
4.7.1. Quantification Method Validation
Linearity, LDQ and LOQ
Matrix Effect and Recovery
Precision and Accuracy of the System
4.7.2. PCA of Extracts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Medini, F.; Fellah, H.; Ksouri, R.; Abdelly, C. Total phenolic, flavonoid and tannin contents and antioxidant and antimicrobial activities of organic extracts of shoots of the plant Limonium delicatulum. J. Taibah Univ. Sci. 2014, 8, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Jallali, I.; Megdiche, W.; M’Hamdi, B.; Oueslati, S.; Smaoui, A.; Abdelly, C.; Ksouri, R. Changes in phenolic composition and antioxidant activities of the edible halophyte Crithmum maritimum L. with physiological stage and extraction method. Acta Physiol. Plant. 2012, 34, 1451–1459. [Google Scholar] [CrossRef]
- Bansal, T.; Awasthi, A.; Jaggi, M.; Khar, R.K.; Talegaonkar, S. Pre-clinical evidence for altered absorption and biliary excretion of irinotecan (CPT-11) in combination with quercetin: Possible contribution of P-glycoprotein. Life Sci. 2008, 83, 250–259. [Google Scholar] [CrossRef]
- Ozben, T. Mechanisms and strategies to overcome multiple drug resistance in cancer. FEBS Lett. 2006, 22, 2903–2909. [Google Scholar] [CrossRef] [Green Version]
- Borska, S.; Sopel, M.; Chmielewska, M.; Zabel, M.; Dziegiel, P. Quercetin as a potential modulator of P-glycoprotein expression and function in cells of human pancreatic carcinoma line resistant to daunorubicin. Molecules 2010, 15, 857–870. [Google Scholar] [CrossRef]
- Spengler, G.; Ocsovszki, I.; Tönki, Á.S.; Saijo, R.; Watanabe, G.; Kawase, M.; Molnár, J. Fluorinated β-Diketo Phosphorus Ylides Are Novel Inhibitors of the ABCB1 Efflux Pump of Cancer Cells. Anticancer Res. 2015, 35, 5915–5919. [Google Scholar]
- Wang, Z.; Wong, I.L.; Li, F.X.; Yang, C.; Liu, Z.; Jiang, T.; Jiang, T.F.; Chow, L.M.; Wan, S.B. Optimization of permethyl ningalin B analogs as P-glycoprotein inhibitors. Bioorg. Med. Chem. 2015, 23, 5566–5573. [Google Scholar] [CrossRef]
- Wong, I.L.; Wang, B.C.; Yuan, J.; Duan, L.X.; Liu, Z.; Liu, T.; Li, X.M.; Hu, X.; Zhang, X.Y.; Jiang, T.; et al. Potent and Nontoxic Chemosensitizer of P-Glycoprotein-Mediated MultidrugResistance in Cancer: Synthesis and Evaluation of Methylated Epigallocatechin, Gallocatechin, and Dihydromyricetin Derivatives. J. Med. Chem. 2015, 58, 4529–4549. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Ling, V. The molecular basis of multidrug resistance in cancer: The early years of P-glycoprotein research. FEBS Lett. 2006, 580, 998–1009. [Google Scholar] [CrossRef] [Green Version]
- Bansal, T.; Jaggi, M.; Khar, R.K.; Talegaonkar, S. Emerging Significance of Flavonoids as P Glycoprotein Inhibitors in Cancer Chemotherapy. J. Pharm. Pharmaceut. Sci. 2009, 12, 46–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, W.; Schutte, M.E.; Williamson, G.; van Zanden, J.J. Flavonoid-mediated inhibition of intestinal ABC transporters may affect the oral bioavailability of drugs, food-borne toxic compounds and bioactive ingredients. Biomed. Pharmacother. 2006, 60, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Chieli, E.; Romiti, N.; Rodeiro, I.; Garrido, G. In vitro effects of Mangifera indica and polyphenols derived on ABCB1/P-glycoprotein activity. Food Chem. Toxicol. 2009, 47, 2703–2710. [Google Scholar] [CrossRef] [PubMed]
- Chieli, E.; Romiti, N.; Rodeiro, I.; Garrido, G. In vitro modulation of ABCB1/P-glycoprotein expression by polyphenols from Mangifera indica. Chem. Biol. Interact. 2010, 186, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.; Salgueiro, L.; Fortuna, A.; Cavaleiro, C. P-glycoprotein mediated efflux modulators of plant origin: A Short Review. Nat. Prod. Commun. 2016, 11, 699–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolosa, L.; Rodeiro, I.; Donato, M.T.; Herrera, J.A.; Delgado, R.; Castell, J.V.; Gómez-Lechón, M.J. Multiparametric evaluation of the cytoprotective effect of the Mangifera indica L. stem bark extract and mangiferin in Hep-G2 cells. J. Pharm. Pharmacol. 2013, 65, 1073–1082. [Google Scholar] [CrossRef]
- del Barrio, G.; Spengler, I.; García, T.H.; Roque, A.; Álvarez, A.L.; Calderón, J.S.; Parra, F. Antiviral activity of Ageratina havanensis and major chemical compounds from the most active fraction. Braz. J. Pharmacog. 2011, 21, 915–920. [Google Scholar] [CrossRef]
- García, T.H.; Quintino da Rocha, C.; Dias, M.J.; Pino, L.L.; del Barrio, G.; Roque, A.; Pérez, C.E.; Campaner dos Santos, L.; Spengler, I.; Vilegas, W. Comparison of the Qualitative Chemical Composition of Extracts from Ageratina havanensis Collected in Two Different Phenological Stages by FIA-ESI-IT-MSn and UPLC/ESI-MSn: Antiviral Activity. Nat. Prod. Commun. 2017, 12, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Hosu, A.; Cristea, V.M.; Cimpoiu, C. Analysis of total phenolic, flavonoids, anthocyanins and tannins content in Romanian red wines: Prediction of antioxidant activities and classification of wines using artificial neural networks. Food Chem. 2014, 150, 113–118. [Google Scholar] [CrossRef]
- Lee, L.-S.; Choi, E.-J.; Kim, C.-H.; Sung, J.-M.; Kim, Y.-B.; Seo, D.-H.; Choi, H.-W.; Choi, Y.-S.; Kum, J.-S.; Park, J.-D. Contribution of flavonoids to the antioxidant properties of common and tartary buckwheat. J. Cereal. Sci. 2016, 68, 181–186. [Google Scholar] [CrossRef]
- Ben Sghaier, M.; Skandrani, I.; Nasr, N.; Franca, M.G.; Chekir-Ghedira, L.; Ghedira, K. Flavonoids and sesquiterpenes from Tecurium ramosissimum promote antiproliferation of human cancer cells and enhance antioxidant activity: A structure-activity relationship study. Environ. Toxicol. Pharmacol. 2011, 32, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimzadeh, M.A.; Pourmorad, F.; Bekhradnia, A.R. Iron chelating activity screening, phenol and flavonoid content of some medicinal plants from Iran. Afr. J. Biotechnol. 2008, 7, 3188–3192. [Google Scholar]
- Sirivibulkovit, K.; Nouanthavong, S.; Sameenoi, Y. Paper-based DPPH Assay for Antioxidant Activity Analysis. Anal. Sci. 2018, 34, 795–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehpour, A.A.; Ebrahimzadeh, M.A.; Fazel, N.S.; Mohammad, N.S. Antioxidant activity of methanol extract of Ferula assafoetida and its essential oil composition. Oils Fats 2009, 60, 405–412. [Google Scholar]
- Halliwell, B.; Gutteridge, J. Antioxidant defences: Endogenous and diet derived. Free Radic. Biol. Med. 2007, 4, 79–186. [Google Scholar]
- Badmus, J.A.; Adedosu, T.O.; Fatoki, J.O.; Adegbite, V.A.; Adaramoye, O.A.; Odunola, O.A. Lipid peroxidation inhibition and antiradical activities of some leaf fractions of Mangifera indica. Acta Pol. Pharm. 2011, 68, 23–29. [Google Scholar]
- De Santis, C.E.; Bray, F.; Ferlay, J.; Lortet-Tieulent, J.; Anderson, B.O.; Jemal, A. International variation in female breast cancer incidence and mortality rates. Cancer Epidemiol. Biomark. Prev. 2015, 10, 1495–1506. [Google Scholar] [CrossRef] [Green Version]
- Bradley, G.; Ling, V. P-glycoprotein, multidrug resistance and tumor progression. Cancer Metastasis Rev. 1994, 13, 223–233. [Google Scholar] [CrossRef]
- Varma, M.V.; Ashokraj, Y.; Dey, C.S.; Panchagnula, R. P-glycoprotein inhibitors and their screening: A perspective from bioavailability enhancement. Pharmacol. Res. 2003, 48, 347–359. [Google Scholar] [CrossRef]
- Aszalos, A. Role of ATP-binding cassette (ABC) transporters in interactions between natural products and drugs. Curr. Drug Metab. 2008, 9, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.W.; Sneed, K.B.; Pan, S.Y.; Cao, C.; Kanwar, J.R.; Chew, H.; Zhou, S.F. Herb-drug interactions and mechanistic and clinical considerations. Curr. Drug Metab. 2012, 13, 640–651. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Basu, S.; Das, S.; Sinha, A.; Biswas, M.K.; Choudhuri, S.K. Induction of intrinsic and extrinsic apoptosis through oxidative stress in drug-resistant cancer by a newly synthesized Schiff base copper chelate. Free Radic. Res. 2016, 50, 426–446. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Lu, Y.; Ji, Z.N.; Liu, G.Q. Up-regulation of P-glycoprotein expression by glutathione depletion-induced oxidative stress in rat brain microvessel endothelial cells. J. Neurochem. 2006, 98, 1465–1473. [Google Scholar] [CrossRef] [PubMed]
- Qosa, H.; Lichter, J.; Sarlo, M.; Markandaiah, S.S.; McAvoy, K.; Richard, J.P.; Jablonski, M.R.; Maragakis, N.J.; Pasinelli, P.; Trotti, D. Astrocytes drive up-regulation of the multidrug resistance transporter ABCB1 (P-glycoprotein) in endothelial cells of the blood-brain barrier in mutant superoxide dismutase 1-linked amyotrophic lateral sclerosis. Glia 2016, 64, 1298–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanković, T.; Dankó, B.; Martins, A.; Dragoj, M.; Stojković, S.; Isaković, A.; Wang, H.C.; Wu, Y.C.; Hunyadi, A.; Pešić, M. Lower antioxidative capacity of multidrug-resistant cancer cells confers collateral sensitivity to protoflavone derivatives. Cancer Chemother. Pharmacol. 2015, 76, 555–565. [Google Scholar] [CrossRef]
- Wartenberg, M.; Fischer, K.; Hescheler, J.; Sauer, H. Redox regulation of P-glycoprotein-mediated multidrug resistance in multicellular prostate tumor spheroids. Int. J. Cancer 2000, 85, 267–274. [Google Scholar] [CrossRef]
- Wartenberg, M.; Hoffmann, E.; Schwindt, H.; Grünheck, F.; Petros, J.; Arnold, J.R.; Hescheler, J.; Sauer, H. Reactive oxygen species-linked regulation of the multidrug resistance transporter P-glycoprotein in Nox-1 overexpressing prostate tumor spheroids. FEBS Lett. 2005, 579, 4541–4549. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.A.; Sulaiman, A.A.; Alhaddad, H.; Alhadidi, Q. Natural polyphenols: Influence on membrane transporters. J. Intercult. Ethnopharmacol. 2016, 5, 97–104. [Google Scholar] [CrossRef]
- Michalak, K.; Wesolowska, O. Polyphenols counteract tumor cell chemoresistance conferred by multidrug resistance proteins. Anticancer Agents Med. Chem. 2012, 12, 880–990. [Google Scholar]
- Reyes-Esparza, J.; Gonzaga, A.I.; González-Maya, L.; Rodríguez-Fragoso, L. (-)-Epigallocatechin-3-gallate modulates the activity and expression of P-glycoprotein in breast cancer cells. J. Pharmacol. Clin. Toxicol. 2015, 3, 1044–1051. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, C.P.P.; de Toledo, A.C.; Perini, A.; Pinheiro, N.M.; Hiyane, M.I.; Grecco, S.D.S.; de Fátima Lopes Calvo Tibério, I.; Câmara, N.O.S.; de Arruda Martins, M.; Lago, J.H.G.; et al. Sakuranetin reverses vascular peribronchial and lung parenchyma remodeling in a murine model of chronic allergic pulmonary inflammation. Acta Histochem. 2016, 118, 615–624. [Google Scholar] [CrossRef]
- Arai, H.; Kogure, K.; Sugioka, K.; Nakano, M. Importance of two iron-reducing systems in lipid peroxidation of rat brain: Implications for oxygen toxicity in the central nervous system. Biochem. Int. 1987, 14, 741–749. [Google Scholar]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef]
- Hider, R.C.; Liu, Z.D.; Khodr, H.H. Metal chelation of polyphenols. Methods Enzymol. 2001, 335, 190–203. [Google Scholar]
- Perron, N.R.; Brumaghim, J.L. A review of the antioxidant mechanisms of polyphenol compounds related to iron binding. Cell Biochem. Biophys. 2009, 53, 75–100. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Tabart, J.; Kevers, C.; Pincemail, J.; Defraigne, J.O.; Dommes, J. Comparative antioxidant capacities of phenolic compounds measured by various tests. Food Chem. 2009, 113, 1226–1233. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Shrivastava, A.; Gupta, V.B. Methods for the determination of limit of detection and limit of quantitation of the analytical methods. Chron. Young Sci. 2011, 2, 21. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
| Harvesting Stage | Plant Organs | Extracts | IC50 (μg/mL) | Inhibition P-gp Function (%) |
|---|---|---|---|---|
| Flowering | Leaves | Lf-EtOH | 381.6 ± 7.5 | 35 |
| Lf-EtOAc | 252.5 ± 10.1 | 23 | ||
| Lf-n-BuOH | 302.0 ± 8.0 | 19 | ||
| Vegetative stage | Lv-EtOH | 392.8 ± 6.7 | 58 | |
| Lv-EtOAc | 313.0 ± 12.1 | 45 | ||
| Lv-n-BuOH | 496.5 ± 6.7 | 27 | ||
| Flowering | Stems | Sf-EtOH | 228.2 ± 8.7 | 84 |
| Vegetative stage | Sv-EtOH | 355.7 ± 7.6 | 65 | |
| Flowering | Flowers | F-EtOH | 263.5 ± 8.2 | 85 |
| F-EtOAc | 259.5 ± 10.6 | - | ||
| F-n-BuOH | 315.5 ± 9.9 | - |
| Harvesting Stage | Plant Organs | Extracts | FRAP (μM of Ascorbic Acid Equivalents) | DPPH IC50 (μg/mL) | Lipid Peroxidation Inhibition IC50 (µg/mL) |
|---|---|---|---|---|---|
| Flowering | Leaves | Lf-EtOH | 88.21 ± 3.77 a | 22.65 ± 1.73 a | 16.52 ± 2.19 a |
| Lf-EtOAc | 90.52 ± 3.61 a | 39.27 ± 2.04 a | 21.63 ± 1.78 a | ||
| Lf-n-BuOH | 63.51 ± 2.62 a | 53.12 ± 0.92 a | 38.91 ± 3.52 a | ||
| Vegetative stage | Lv-EtOH | 279.42 ± 8.48 b | 151.90 ± 3.71 b | 41.66 ± 2.87 b | |
| Lv-EtOAc | 234.70 ± 7.93 b | 138.11 ± 2.67 b | 46.23 ± 3.12 b | ||
| Lv-n-BuOH | 231.56 ± 10.14 b | 84.24 ± 3.81 b | 51.60 ± 2.04 b | ||
| Flowering | Stems | Sf-EtOH | 88.43 ± 3.96 a | 39.23 ± 1.92 a | 37.21 ± 2.55 a |
| Vegetative stage | Sv-EtOH | 197.21 ± 3.30 b | 32.72 ± 7.23 a | 48.32 ± 3.87 b | |
| Flowering | Flowers | F-EtOH | 169.91 ± 7.63 a | 29.45 ± 1.99 a | 21.35 ± 2.66 a |
| F-EtOAc | 167.21 ± 5.34 a | 38.77 ± 2.64 b | 32.34 ± 2.01 b | ||
| F-n-BuOH | 91.66 ± 3.21 b | 54.13 ± 2.88 c | 30.27 ± 3.81 b | ||
| Sakuranetin | 319.38 ± 6.65 | - | - | ||
| 7-methoxyaromadendrin | 238.45 ± 2.11 | - | - | ||
| Ascorbic acid | - | 21.97 ± 0.84 | - | ||
| Trolox-C | 10.64 ± 1.02 | ||||
| Harvesting Stage | Plant Organs | Extracts | Concentration (μg/mL) | |
|---|---|---|---|---|
| Sakuranetin | 7-Methoxyaromadendrin | |||
| Flowering | Leaves | Lf-EtOH | Below LOQ | 1.27 |
| Lf-EtOAc | 46.70 | 16.8 | ||
| Lf-n-BuOH | Below LOQ | Below LOQ | ||
| Vegetative stage | Lv-EtOH | 13.20 | 2.90 | |
| Lv-EtOAc | 87.02 | 30.2 | ||
| Lv-n-BuOH | Below LOQ | Below LOQ | ||
| Flowering | Stems | Sf-EtOH | Below LOQ | Below LOQ |
| Vegetative stage | Sv-EtOH | Below LOQ | Below LOQ | |
| Flowering | Flowers | F-EtOH | Below LOQ | Below LOQ |
| F-EtOAc | 29.90 | 6.30 | ||
| F-n-BuOH | Below LOQ | Below LOQ | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García, T.H.; Rocha, C.Q.d.; Delgado-Roche, L.; Rodeiro, I.; Ávila, Y.; Hernández, I.; Cuellar, C.; Lopes, M.T.P.; Vilegas, W.; Auriemma, G.; et al. Influence of the Phenological State of in the Antioxidant Potential and Chemical Composition of Ageratina havanensis. Effects on the P-Glycoprotein Function. Molecules 2020, 25, 2134. https://doi.org/10.3390/molecules25092134
García TH, Rocha CQd, Delgado-Roche L, Rodeiro I, Ávila Y, Hernández I, Cuellar C, Lopes MTP, Vilegas W, Auriemma G, et al. Influence of the Phenological State of in the Antioxidant Potential and Chemical Composition of Ageratina havanensis. Effects on the P-Glycoprotein Function. Molecules. 2020; 25(9):2134. https://doi.org/10.3390/molecules25092134
Chicago/Turabian StyleGarcía, Trina H., Claudia Quintino da Rocha, Livan Delgado-Roche, Idania Rodeiro, Yaiser Ávila, Ivones Hernández, Cindel Cuellar, Miriam Teresa Paz Lopes, Wagner Vilegas, Giulia Auriemma, and et al. 2020. "Influence of the Phenological State of in the Antioxidant Potential and Chemical Composition of Ageratina havanensis. Effects on the P-Glycoprotein Function" Molecules 25, no. 9: 2134. https://doi.org/10.3390/molecules25092134
APA StyleGarcía, T. H., Rocha, C. Q. d., Delgado-Roche, L., Rodeiro, I., Ávila, Y., Hernández, I., Cuellar, C., Lopes, M. T. P., Vilegas, W., Auriemma, G., Spengler, I., & Rastrelli, L. (2020). Influence of the Phenological State of in the Antioxidant Potential and Chemical Composition of Ageratina havanensis. Effects on the P-Glycoprotein Function. Molecules, 25(9), 2134. https://doi.org/10.3390/molecules25092134