3.1. Yield and G.C.–M.S. Analysis
As shown in
Table 2, the most considerable quantity of essential oils extracted, calculated based on the dry weight of the plant (
v/
w), was recorded in L8b (5.20%,
v/
w), followed by L9b (3.80%,
v/
w), L6 (1.70%,
v/
w) and L7 (1.30%,
v/
w). It is important to note that the yield values increase at the flowering stage more than before and after flowering. As reported by several authors, the EOs concentration in inflorescences is dependent on the variety and environmental conditions [
55,
56,
57].
In the present study, thirty-five compounds were identified with G.C.–M.S. analyses (
Table 3). A percentage of 67.50 to 100% of the total EOs composition of the flowers/leaves of
L. multiflora M were identified. It can be observed that the content (% of essential oil) of the determined compounds varies between EOs of the plants collected in the two provinces.
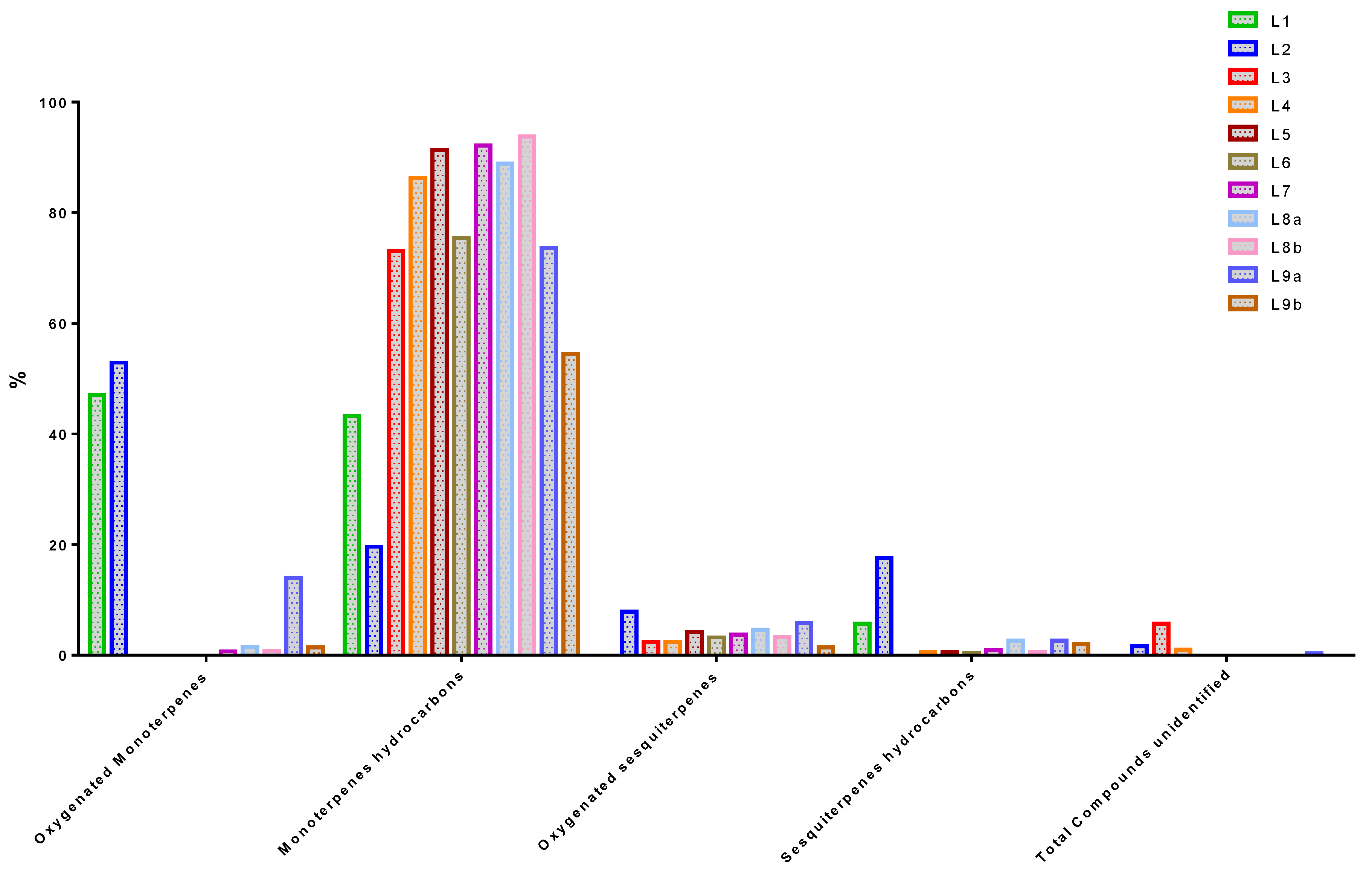
According to
Table 4 and
Figure 3, the main compound classes represented in all EOs were monoterpene hydrocarbons, oxygenated monoterpenes, sesquiterpene hydrocarbons, and oxygenated sesquiterpenes. Monoterpene hydrocarbons are the most common compounds in EOs, with a proportion ranging from 19.60% to 93.82%. L8b, L8a, L7, L4, L5 and L9b respectively are the samples that contain the most of them. Next, the monoterpenes oxygenated compounds which are present from 0.00% to 52.90% in EOs. The highest detected concentrations of these compounds are in L1/L2. They are followed by sesquiterpenes hydrocarbons present in proportions ranging from 0 to 17.60%. L1/L2 are the most concentrated in these compounds. Finally, oxygenated sesquiterpenes are the least present in the different oils. L2 and L9a are the samples that contain the most (7.90% and 5.80% respectively).
Limonene (p-mentha-1,8-diene) is the only monoterpene that appears in all EOs. It is the most predominant compound in the EOs of L4 to L8a et L9b with a percentage ranging from 25.95% to 40.70%. Moreover, it is more present in the plants sampled in M’banza Kongo (40.70%) and Cazengo (32.83%) than in Tomboco (25.95%). By comparing the chemical profiles in the table, the synthesis of Limonene showed an evident fluctuation during the different stages of plant growth. It is present throughout the phases but is slightly higher in samples taken before flowering than in those selected during and after flowering. These differences may be due to the type of harvested part of the plant, the physiological metabolism, phenology or environmental changes in the biotic and abiotic factors and the geographical aspect, which were not the same in this study [
59,
60,
61,
62].
Limonene has wide applications as a flavour additive in food, beverages, fragrances, cosmetics and household products [
63]. It is one of the most common terpenes in nature and a major constituent of numerous essential oils from Citrus. [
64]. In genus
Lippia, Limonene has been identified in
Lippia turbinata Griseb. (60.60%) [
65],
Lippia alnifolia (47.20%) [
66],
Lippia alba (Mill.) N.E. Br. ex Britton and P. Wilson (30.60 to 33.00%) [
67,
68],
Lippia junelliana (Moldenke) Trunk. (23.12%) [
65], and
Lippia citriodora Kunth (syn.
Aloysia citriodora Palau) (10.70%) [
67].
According to the results (
Table 3), Piperitenone would also follow Limonene in almost all EOs profiles. Among the eleven samples, it is present in eight EOs at a rate of 11.58% to 34.83%. Trans-tagetone is also detected in L4, L5, L6, L7 and L8b, with a proportion ranging from 8.50% to 12.34%. The two compounds Neral and Citral are only present in the L1 (25.3%, 21.70%) and L2 (24.40%, 28.30%) samples, respectively. We also notice the presence of p-cymene in the L1 (10.40%), Carvacrol in L9a (11.50%) and a significant concentration of Ipsenone in L4 (19.40%). Artemisia ketone has been detected in L6 (20.30%) and L9a (28.30%) and is not a common compound in the genus
Lippia. It is most often found in different species of the genus Artemisia [
69]. As for verbenone, it is present in large quantities in only L3 (55.90%). It is a natural organic compound classified as bicyclic monoterpenoids that are found naturally in a variety of plants. Verbenone is a compound that has an odour and/or flavour of camphor, celery and menthol. It can be found in many food items such as spearmint, cabbage, white cabbage, and rosemary, which makes verbenone a potential biomarker for the consumption of these food products [
70]. The anti-aggregation pheromone verbenone has been used to reduce the attack rates on pines by the mountain pine beetle,
Dendroctonus ponderosae Hopkins. In northern America, it the most damaging insect pest of lodgepole pine Pinus contorta Douglas ex Loudon [
71,
72]. In genus
Lippia, it has been detected in the essential oil of
Lippia citriodora (15.64%) and
Lippia alba (Mill.) N.E. Brown (21.74%) [
73,
74].
It is interesting to note that although
Lippia species have grown under the same edaphic and climatic conditions, there is a difference in the chemical profiles observed in plants groups from the same geographical areas. Three outgoing lots can be distinguished: L2 (Citral/Neral/Elemol), L3 (Verbenone/Ocimenone/Limonene), and L9a (Artemisia ketone/Limonene/Piperitenone). This phenotypic variability within the same species from the same region can be explained by the spatial influence of the environment on the plant [
56,
75]. The importance of spatial distributions in pollination, competition, herbivory, nutrient cycling and other ecological processes in the variability of the content and quality of secondary metabolites is well documented [
56,
57,
76,
77,
78]. Therefore, detection and measurement of the spatial pattern are relevant for understanding phenotypic variability along a geographical gradient within the same community or the same population [
79]. However, phenology and genetic or developmental stage of plant organs should also be considered [
80,
81]
According to previous studies, a high chemical polymorphism has also been observed at the level of African
Lippia species. In contrast to the Angolan species, high concentrations of 1,8-cineole and Sabinene were found in oils from Nigeria [
82], Togo [
83], Ivory Coast [
29], Benin [
84] and Ghana [
18]. A Geraniol, Geranial/Neral, Thymol, Linalool, Tagetone/Ipsenol, Epoxy-myrcene, p-cymene/Thymol/Ethyl acetate, Myrtenol/Linalool/1,8-cineole, and Nerolidol chemotypes were also described [
29,
37,
82,
83,
85]. These studies thus demonstrate that the Angolan species is atypical in its chemical composition compared to its neighbours.
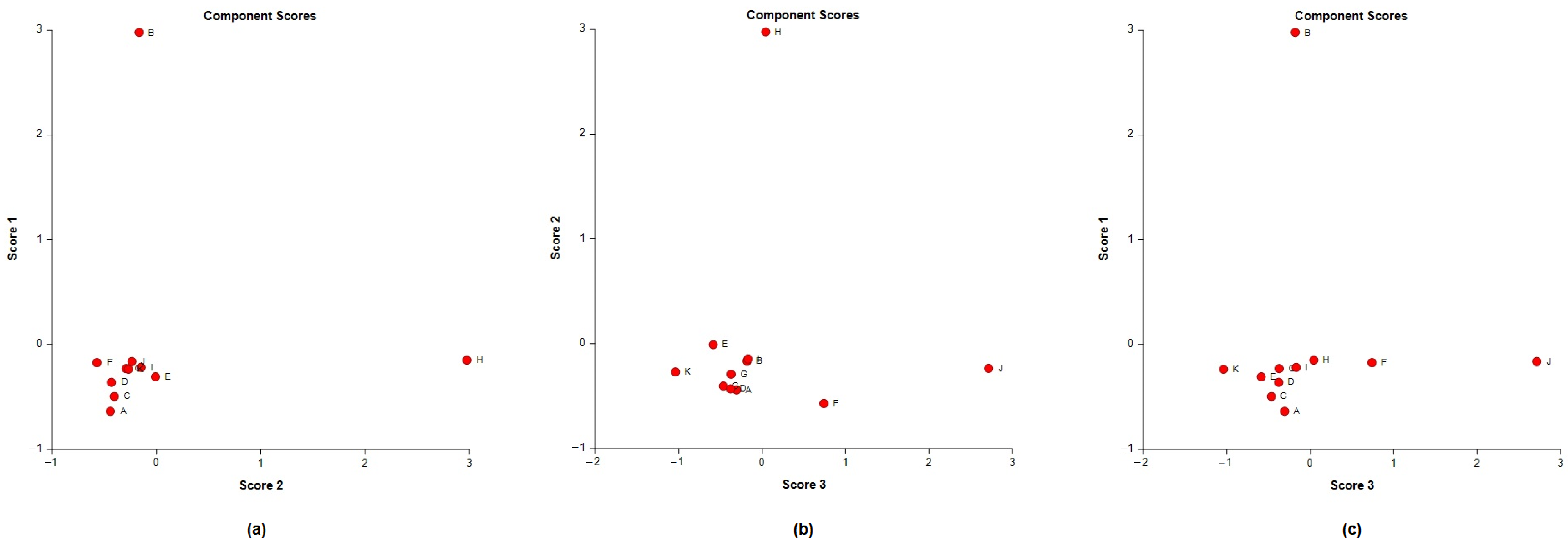
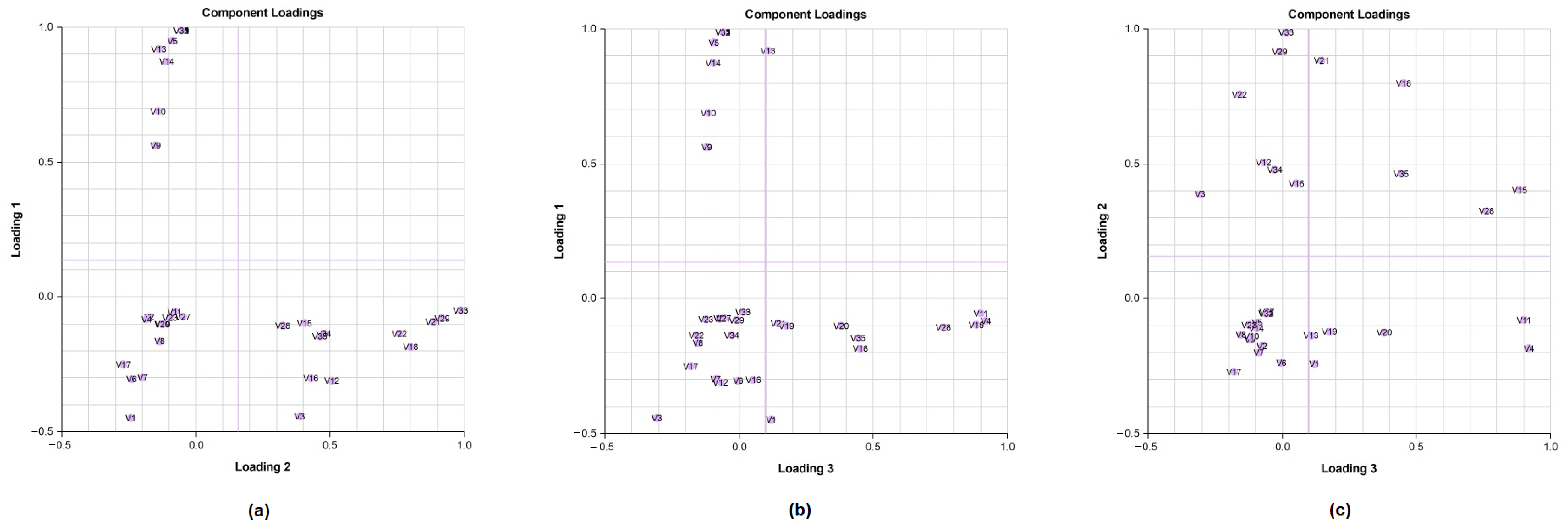
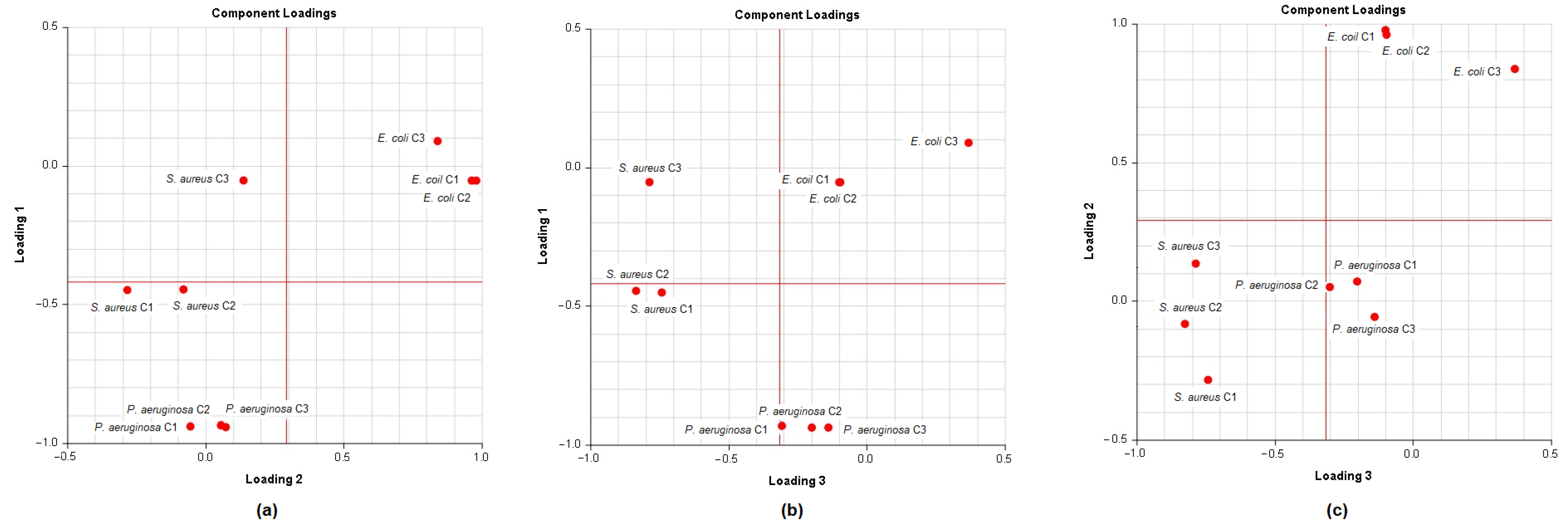
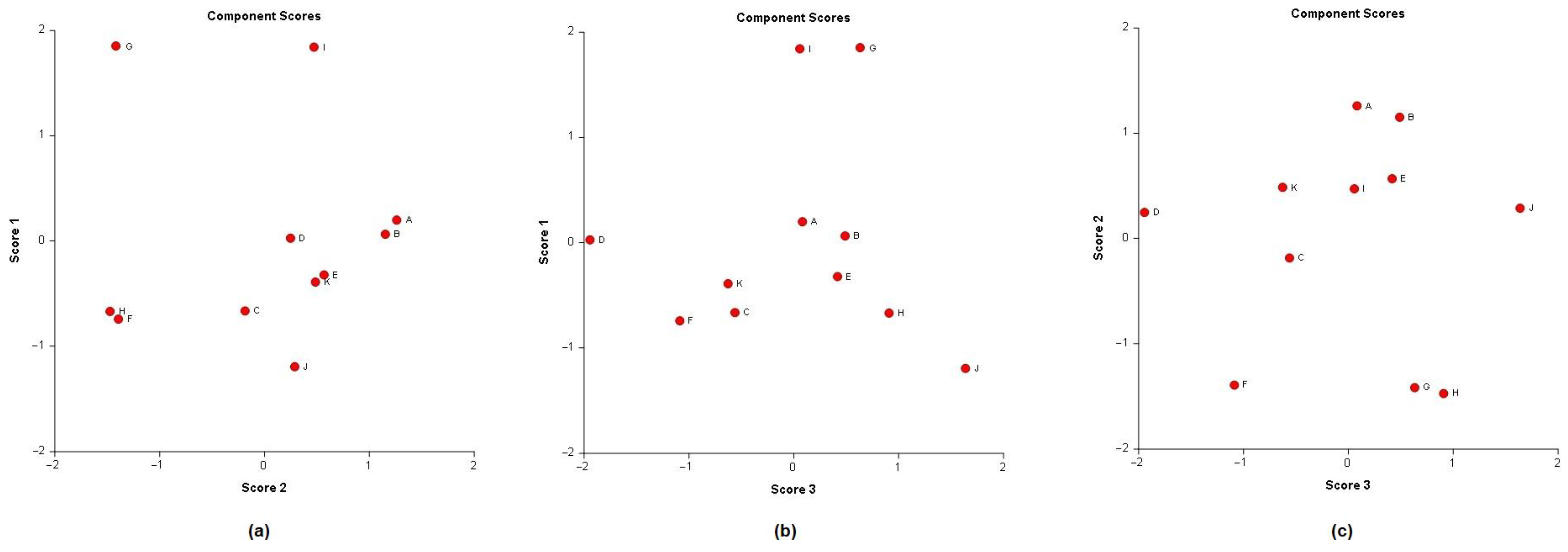
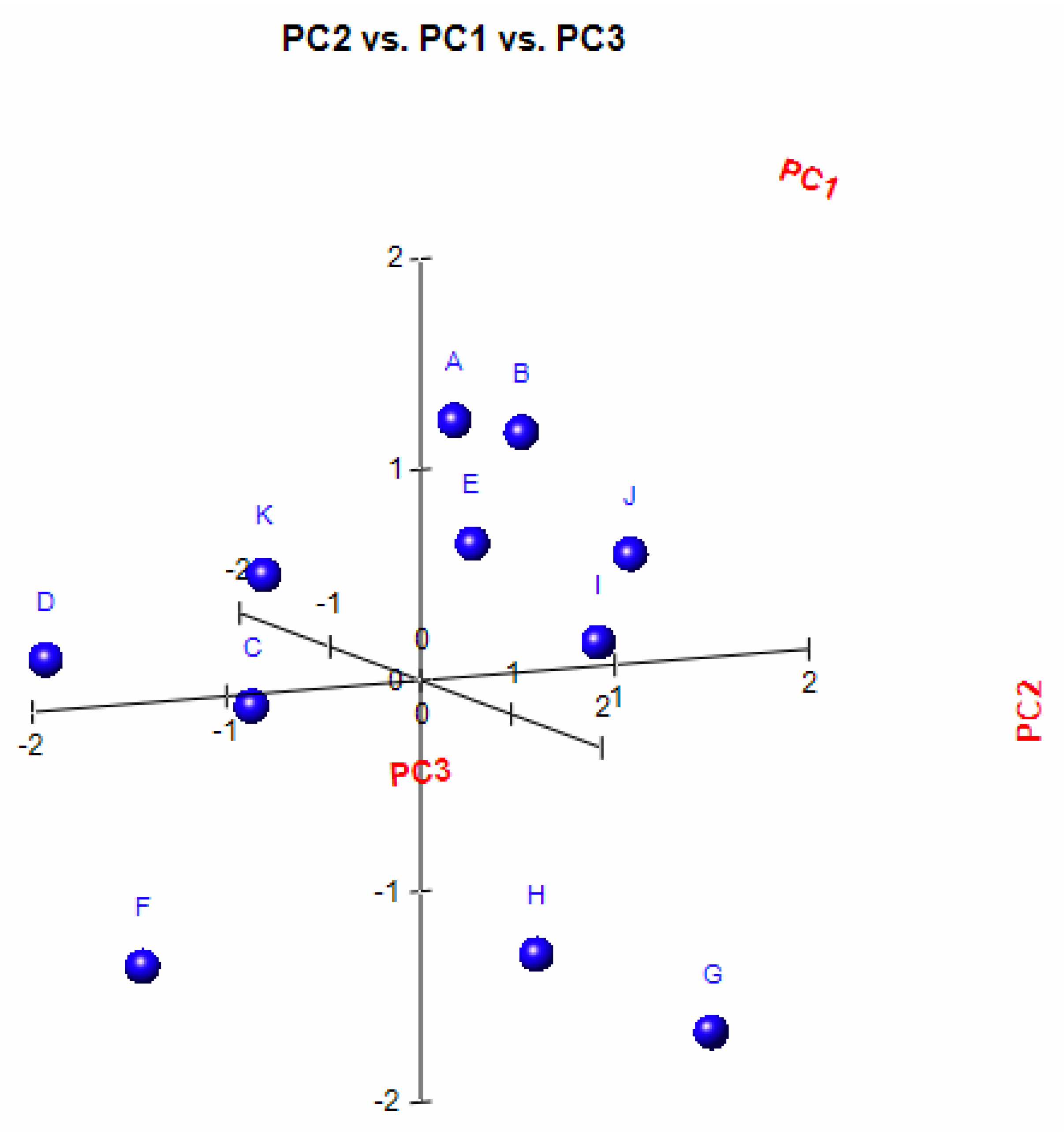
A principal component analysis (PCA) was conducted on the chemical content data for each
Lippia oil to obtain a basic overview of the data structure and to identify similarities and specific grouping patterns. Data raw (number of principal components (PCs) and per cent variance of the first three PCs) for eight PCA models are presented in
Table S1. Mutual projections of factor scores and their loadings for the first three PCs have been presented in
Figure 4,
Figure 5 and
Figure 6. We obtained the following variance percentage for the principal components: 26.37% (PC1), 18.70% (PC2), and 11.21% (PC3) which resulted in a model that would explain approximately 56.28% of the total variance (
Figure 4). The scoring plots (
Figure 5) distinguishes three oils that come out of the sample batch (L2, L8a, L9a). The loading plots (
Figure 6) implies that the most influential compounds for discriminating L2 from other oils are γ-terpinene, Germacrene D, Elemol, Citral, Neral, Hedycaryol, and Zingiberene. The presence of a high concentration of Carvacrol and Artemisia Ketone separates L9a from the samples. Humulene oxide, Carvone, Trans-carveol, Trans-carvyl acetate and Caryophyllene oxide also contribute to the separation of L8a from the plants. Other oils (L1/L3/L4/L5/L6/L7/L8b/L9b), however, are joined into a single group by their common chemical composition, including Limonene, α-humulene, Ipsenone, Myrcene, p-cymene, Trans-tagetone, β-caryophyllene, Piperitenone and p-mentha-trans-2,8-dien-1-ol. We can also distinguish loading plot of Verbenone, Carveyl acetate and Vervenol in this group.
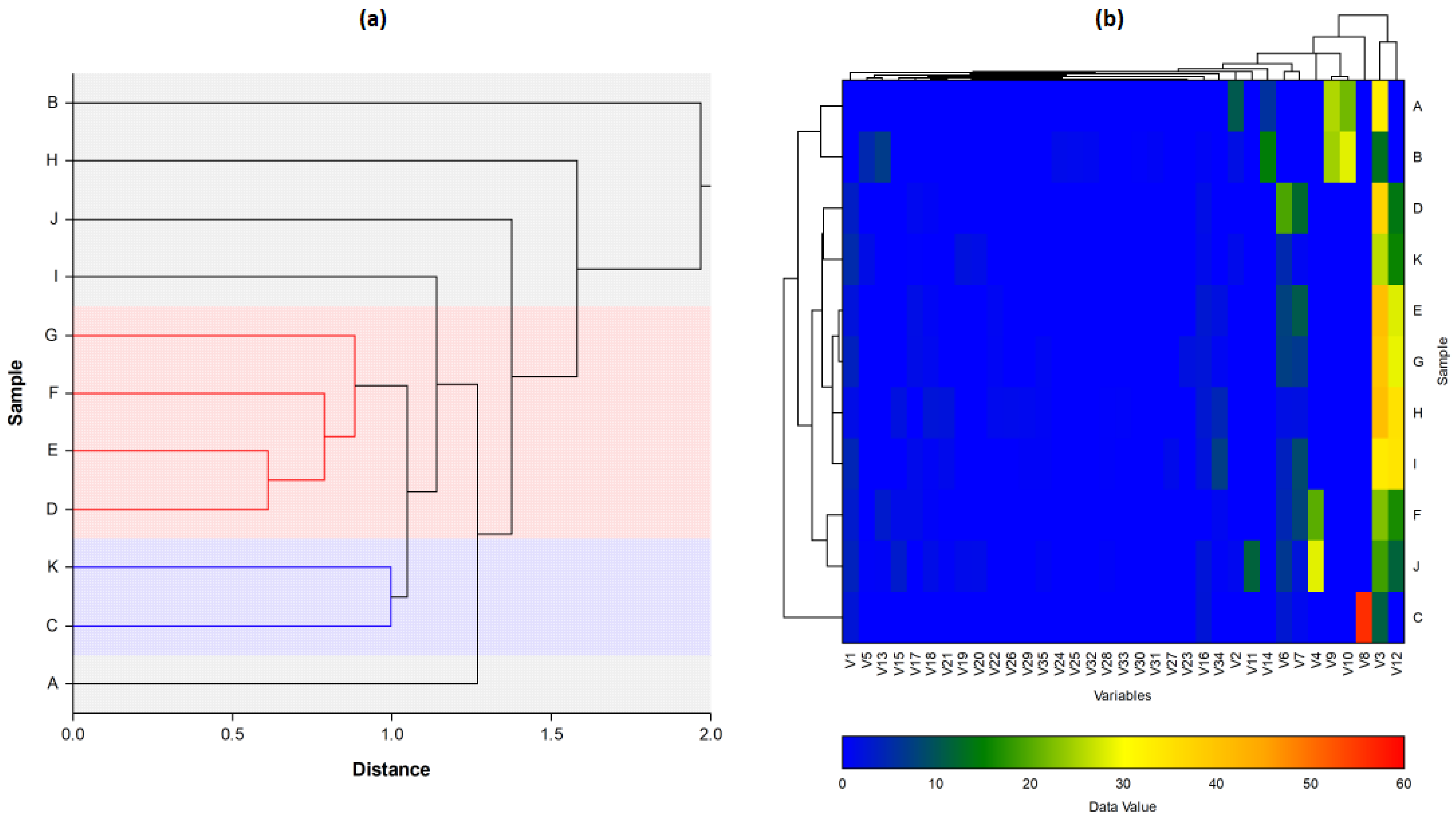
For further characterisation of the
Lippia EOs, the hierarchical cluster analysis (HCA), followed by the heat-map using the abundance of individual volatiles were performed (
Figure 7. The total HCA dendrogram (
Figure 7a) showed that the volatile profile of L4/L5/L6/L7 and L3/L9b were close to each other. Also, L2, L8a, and L9a are isolated from the others by their chemical profiles. The oil distribution is the same as shown by the PCs. The HCA results generated using the abundance of the volatile compounds (
Figure 7b) also showed variability between the populations from the two provinces, but also intraspecific variability between sub-groups within a population. This phenomenon has also been observed in several species of
Lippia, such as
L. origanoides,
L. alba,
L. lupulina,
L. velutina,
L. sidoides,
L. salviifolia, and
L. grata [
56,
86]. Furthermore, it should also be noted that for the L8/L9 plants, the EO chemical composition of their leaves (L8a/L9a) is different from that of their flowers (L8b/L9b). Indeed, the functioning of each organ can be independent of the others through authentic genetic machinery [
87,
88]. The chemical composition of an essential oil varies considerably not only according to individual genetic variability but also according to the phenological stage and organ of the plant.
3.2. Antimicrobial Vapour Phase Test
The antibacterial activity of
Lippia multiflora Moldenke EOs has been tested using a vapour evaluation technique. It exclusively allows us to detect the antimicrobial power of EOs volatile components against three bacterial strains
Staphylococcus aureus,
Escherichia coli and
Pseudomonas aeruginosa. The results are reported in
Table 5.
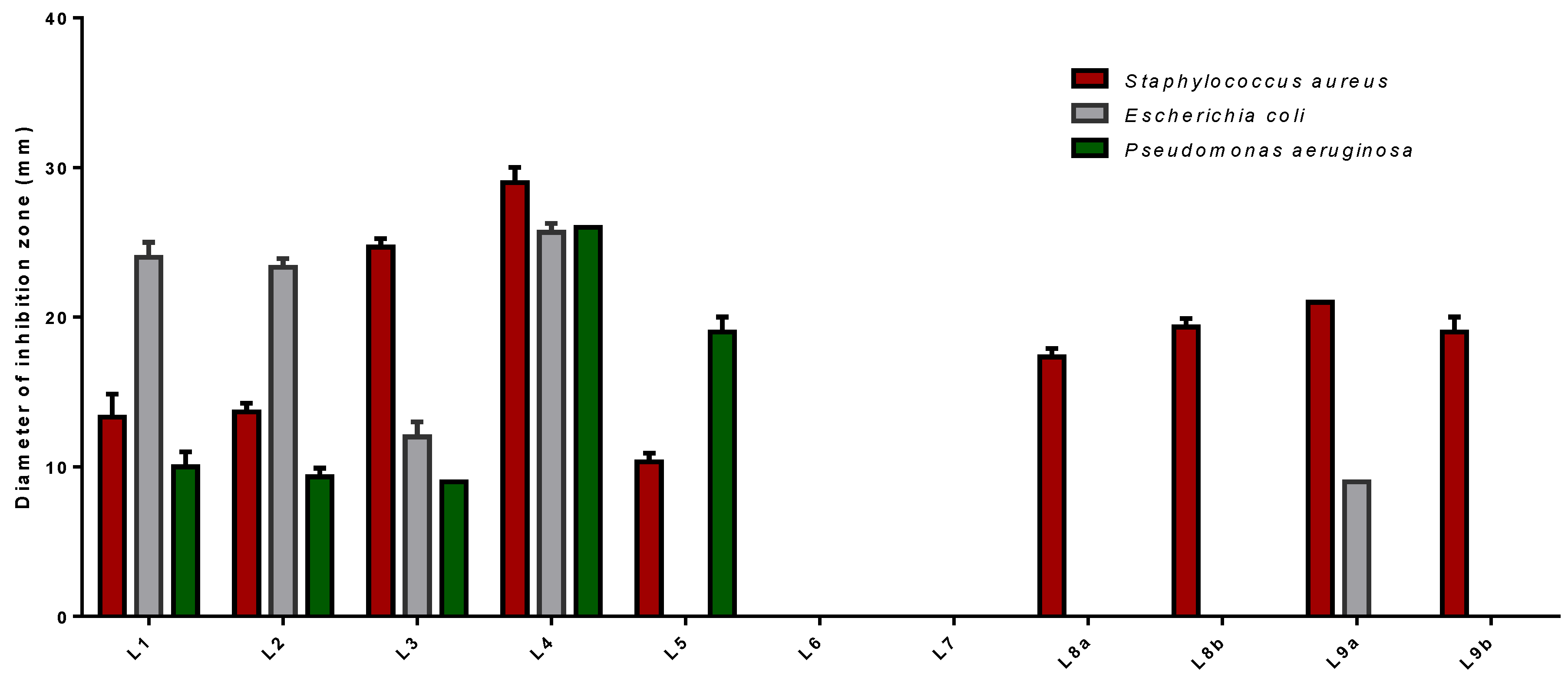
The volatile oil of L1/L2/L3/L4/L5/L9a significantly inhibits the growth of tested microorganisms although they were more potent against
S. aureus than against other strains of bacteria (
Figure 8). Among those, L4 gave the strongest inhibition activity against all investigated pathogens with diameter values in the range of 25.67 ± 0.58 mm to 29.00 ± 1.00 mm. It seems that the presence of a high concentration of Limonene (37.20%), Ipsenone (19.40%), Piperitenone (14.00%), and Trans-tagetone (12.34%), in its oils was responsible for this higher antibacterial activity. Other essential oils L8a (17.33 ± 0.58 mm), L8b (19.33 ± 0.58 mm) and L9b (19.00 ± 1.00 mm) exhibited antimicrobial only against
P. aeruginosa. However, L6/L7 are inefficient.
It is worthy to note from the results of the study that
P. aeruginosa is the most resistant to the action of EOs. This bacterium is an opportunistic human pathogen responsible for several different nosocomial infections [
89]. Through the presence of intrinsic resistance mechanisms and the development of irreversible mutations and adaptations,
P. aeruginosa can survive and multiply in several environments and over a wide range of temperatures [
90]. It is resistant to all types of conventional antibiotics, as it has an exceptional capacity to develop biofilms that are difficult to control [
91]. The indiscriminate misuse of antimicrobial drugs would lead to an increase in bacterial resistance in chronic disease [
92]. As a result, this bacterium is now a global health threat [
92,
93]. One approach to overcome this problem is to use essential plant oils that may represent a promising source for new resistance modifying agents [
93]. In our study, most of the oils were effective against
P. aeruginosa. However, the antibacterial activity depends on the volatility of the chemical compounds present in these EOs [
94]. Indeed, the vapour generated by essential oils has a more significant antimicrobial effect than that of direct contact with these oils in liquid form. Lipophilic molecules in the aqueous phase could associate and form micelles which limit the fixation of EOs to microorganisms [
95]. According to Kloucek et al., there is variability in the phenomenon of essential oils’ volatility. Each EO presents a mixture of chemical compounds that have specific volatility. Each compound diffuses into the environment with its specific speed according to its molecular weight. The vapours interact together to reach an equilibrium that would allow the destruction of microorganisms, but only in a closed environment [
96].
3.3. Well Diffusion Agar Technique
The essential oil from the different
L. multiflora Mold. collections were tested for their antibacterial activity against
Staphylococcus aureus,
Escherichia coli and
Pseudomonas aeruginosa. Three concentrations (C
1, C
2, C
3) of each EO were tested using the wells diffusion agar technique. The results in terms of inhibition zone diameter (mm) are summarised in
Table 6.
In order to analyse the correlation between the chemical composition of the different essential oils and their antibacterial activity, the mean values of the diameters of the inhibition zones were subjected to a PCA and HCA analysis. First, we proceeded to the classification of essential oils according to the values of the diameters of the inhibition zones (
Table 6): we have EO non-effective for a diameter equal to or less than 8 mm; effective (+) for a diameter between 8 and 14 mm; highly effective for a diameter between 14 and 20 mm and extremely effective for a diameter equal to or greater than 20 mm [
97,
98]. The heat-map correlation of the
Lippia samples according to their antibacterial activity was then performed. Data raw (number of principal components (PCs) and per cent variance of the first three PCs) of PCA models are presented in
Table S2. Mutual projections of factor scores and their loadings for the first three PCs have been presented in
Figure 9 and
Figure 10. Starting with 11 variables, the PCA calculated three components having eigenvalues greater than one and representing together 87.97% of the total variance. PC1 accounted for 33.91%, PC2 for 30.02% and PC3 for 24.04% of the variance (
Figure 11).
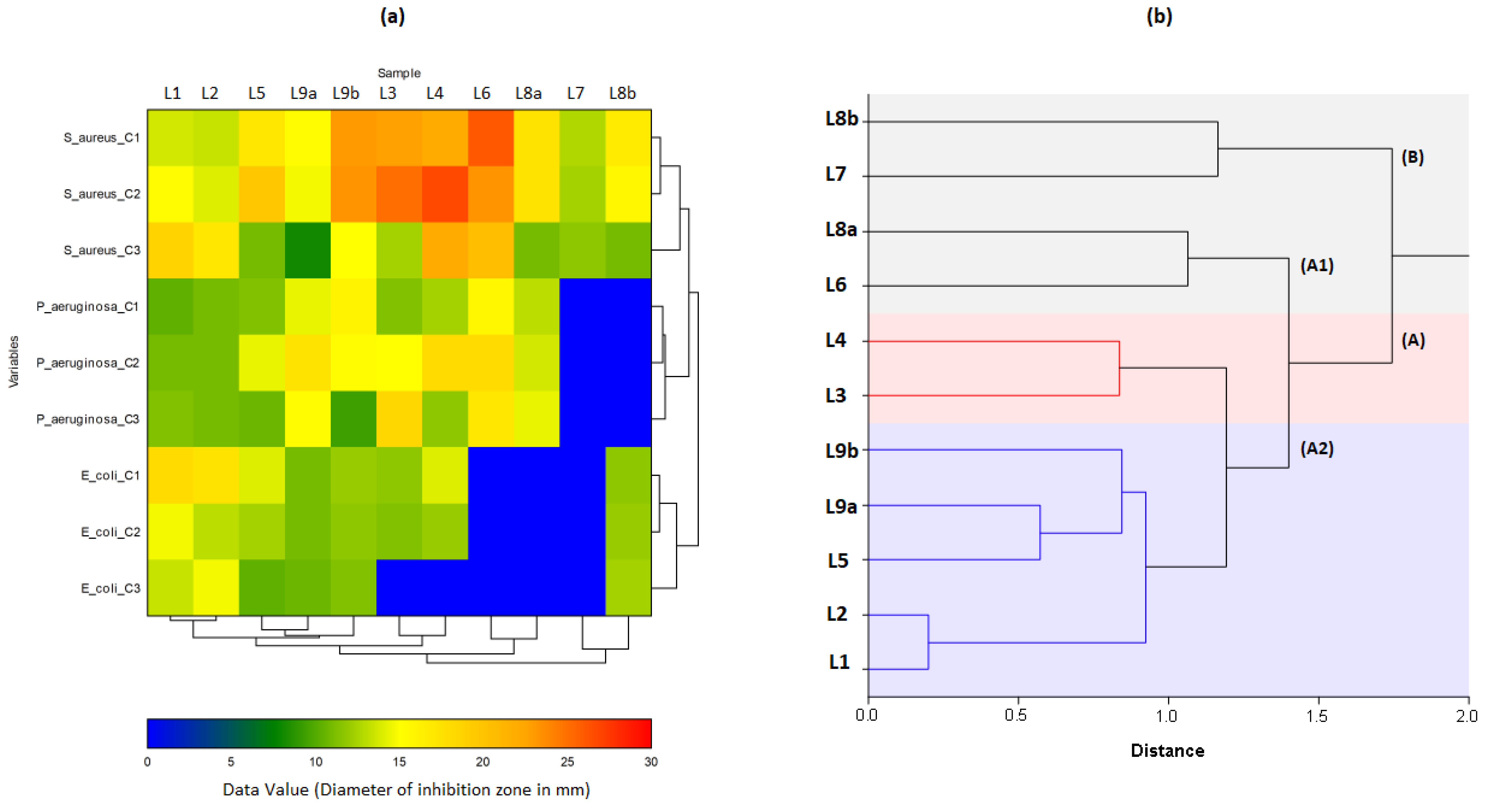
In the heat map (
Figure 12a), almost all the essential oils of the plants used in the study showed an effect against bacteria. The best results recorded were those against
S. aureus, followed by
P. aeruginosa and
E. coli, but they remain much lower than those produced by Gentamicin A (
Table 6). L6C
1/L3C
2/L1C
3 observed the most important effects against
S. aureus, L1C
1/L1C
2/L2C
3 against
E. coli, and L9bC
1/L4C
2/L3C
3 against
P. aeruginosa. It is also noted that all tested concentrations of L6/L7/L8a were ineffective against
E. coli. Similarly, for L7/L8a against
P. aeruginosa. These samples were also recorded as ineffective by the vapour technique previously performed (
Section 3.2).
The dendrogram from HCA (
Figure 12b) presents two distinct main clusters, A and B. Cluster B could correspond to the L7/L8b group, in which L7C
3/L8bC
3 were observed to be effective (+) against
S. aureus (11.67 ± 1.53 mm/11.00 ± 1.00 mm), while L7C
1/C
2/C
3, L8bC
1/C
2/C
3 showed negative results against
P. aeruginosa. With the formation of subgroups in HCA and the results of loading and scores plots in PCA, it can be observed that oils are effectively divided according to their efficacy against the three bacteria. There are subgroups of L1/L2, which have shown better efficacy against
E. coli, and subgroups L7/L8b, L5/L9a/L9b, L3/L4 and L6/L8a, which are relatively more effective against
S. aureus and
P. aeruginosa.
Two distinct groups can be observed in cluster A (
Figure 12): A1 and A2. In gr. A1, we first observe subgroups (1) of L1/L2, whose members L1C
1/L2C
1 are effective (+) and L1C
2/L1C
3 highly effective (++) against
S. aureus. There are also L1C
1/L1C
2, which are effective (+) against
P. aeruginosa. and L2C
1/L2C
3 very effective (++) against
E.coli. Sub-groups (2) L5/L9a/L9b, of which L5C
1/L5C
2/L9aC
1/L9aC
1 are highly effective (++) against
S. aureus, while L5C
1/L5C
2/L5C
3/L9aC
1/C
2/L9aC
3 are effective (+) against
E. coli. On the other hand,
P. aeruginosa is sensitive (+) to L5C
3/L9bC
3, and highly sensitive (++) to L5C
2/L9aC
2/L9bC
1 samples. In subgroup (3) of L3/L4, the L3C
1/L3C
2/L4C
1/L4C
2 are extremely effective (+++) against
S. aureus. At the same time, L3C
1/L3C
2/L4C
1/L4C
2 are effective (+) and L3C
3/L4C
3 non-effective against
E. coli. There are also L3C
1/L4C
1, which are effective (+) and L3C
2/L4C
2 highly effective (++) against
P. aeruginosa. Finally, gr. A2 of L6/L8a showed negative results against
E coli (L6C
1/C
2/C
3, L8aC
1/C
2/C
3), but they are highly effective (++) against
P. aeruginosa (L6C
3/L8C
3).According to our study, the activity of the EOs against gram-negative bacteria was greater than that against gram-positive bacteria. The results were not in line with previous research. Indeed, gram-negative bacteria are more resistant to essential oils than Gram-positive bacteria. Their outer membrane has a thick layer of impermeable peptidoglycan. It makes it difficult for antimicrobial agents to pass through and confers rigidity and resistance to gram-negative bacteria [
99,
100,
101]. The variability of the antibacterial activity of the different
Lippia collections is probably due to the synergistic effect that could occur in the oil and thus potentiate or not its biological activity. According to several studies, the antibacterial activity depends on the quantity of the chemical constituent in the EOs, i.e., at low concentrations they can interfere with the enzymes involved in energy production and at higher concentrations they can denature proteins [
102,
103].
In our study, the plants that have shown the highest activity against the three bacteria strains are the ones that contain more Limonene. This one is reported to have antimicrobial properties against a wide range of bacteria [
104,
105,
106,
107,
108] related to its chemical nature. Indeed, Limonene belongs to the family of cyclic monoterpene hydrocarbons known for their effectiveness against bacteria. Their mechanism of action consists of an accumulation in the plasma membrane and dissipation of the protons motive force causing a loss of membrane integrity [
109,
110]. On the other hand, the results of our study also allow us to suggest that the antibacterial activity of the different
Lippia samples could not be attributed solely to Limonene. Common compounds such as Piperitenone, Ketone artemisia Ipsenone, Ocimenone, Citral, Neral, Elemol and other minor compounds have been detected. They could play a critical role by producing a synergistic and additives effect. Their antimicrobial activity is not attributable to a specific mechanism, but there are several targets in the cell [
101].
In the last part of this study, we were able to determine the Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) values following standard guidelines published by the National Committee for Clinical Laboratory Standards to characterise antibiotic activity [
111]. The MIC/MBC ratio was then calculated to determine the antibacterial power of the EOs studied. The interpretation of the results was based on the classification of Shanmughapriya et al. (2010). EO is considered to be bactericidal when the ratio is less than or equal to 2 and bacteriostatic when it is greater than 2 [
112]. The results are summarised in
Table 7.
There are bacteriostatic (ration greater or equal to 2), bactericidal (whose ratio is between 1 and 2) and very bactericidal oils (whose ration is equal or less than 1) according to the calculated ratios. In the results of activity against
S. aureus, different levels of activity were found with MIC values ranging from 42.52 ± 20.05 to 113.38 μg/mL and MBC values ranging from 47.24 ± 13.36 μg/mL to 170.07 ± 80.16μg/mL (see
Table 7). L7 is the only oil that is bacteriostatic against bacteria. L1/L2/L6/L8a/L8b/L9b EOs are bactericidal, whereas L3/L4/L5/L9a are very bactericidal. For
E. coli, the MIC values are from 85.04 ± 40.09 to 113.38 ± 0.00 μg/mL, and the MBC values are ranging from 85.04 ± 40.09 to 226.75 ± 0.00 μg/mL. The L6/L7/L8a are bacteriostatic, L3/L4/L6/L7/L8a bactericidal, and L1L2/L5/L8b/L9a/L9b very bactericidal EOs. The MIC values for
P. aeruginsa are between 42.52 ± 20.05 and 113.38 ± 0.00 μg/mL, and those for MBC are in the range 56.69 ± 0.00 and 226.75 ± 0.00μg/mL. The oils that have shown bacteriostatic effects are L4 and L7. The others, L5/L8b/L9a are bactericidal and L1/L2/L3/L6/L8a/L9b very bactericidal EOs.
Due to the wide variation in the chemical profiles of
L. multiflora Mold. essential oils, the antibacterial activity is also well diversified in the literature [
10,
32,
113,
114,
115]. According to the study by Bassolé et al. (2010),
Lippia from Burkina-Faso was the most effective among the plants tested against
S. aureus with a MIC = 1.2 ± 0 mg/mL [
32]. The Ivory Coast plant gave better antibacterial activity against
S. aureus and
P. aeruginosa with MIC = MBC = 0.9 mg/mL [
32,
114]. Moreover, MIC values range from 3 to 96 × 10
−3mL
−1 for
Lippia from Gabon and no antibacterial activity was observed for Nigerian plant against
S. aureus or
E. coli [
82,
116]. The antimicrobial activity of any essential oil is not attributable to just one mechanism but to several ones widely described in the literature [
89]. Factors such as climatic and environmental conditions, the origin of the plant, the plant’s adaptive metabolism, the harvesting season, the part of the plant involved in the extraction, the distillation conditions, the microbiological methods used, as well as the susceptibility of the bacterial strains would make it difficult to compare the results obtained by different groups of researchers [
117,
118].


 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}