Different Reactivity of Raw Starch from Diverse Potato Genotypes


Abstract
:1. Introduction
2. Results
2.1. HMF Synthesis
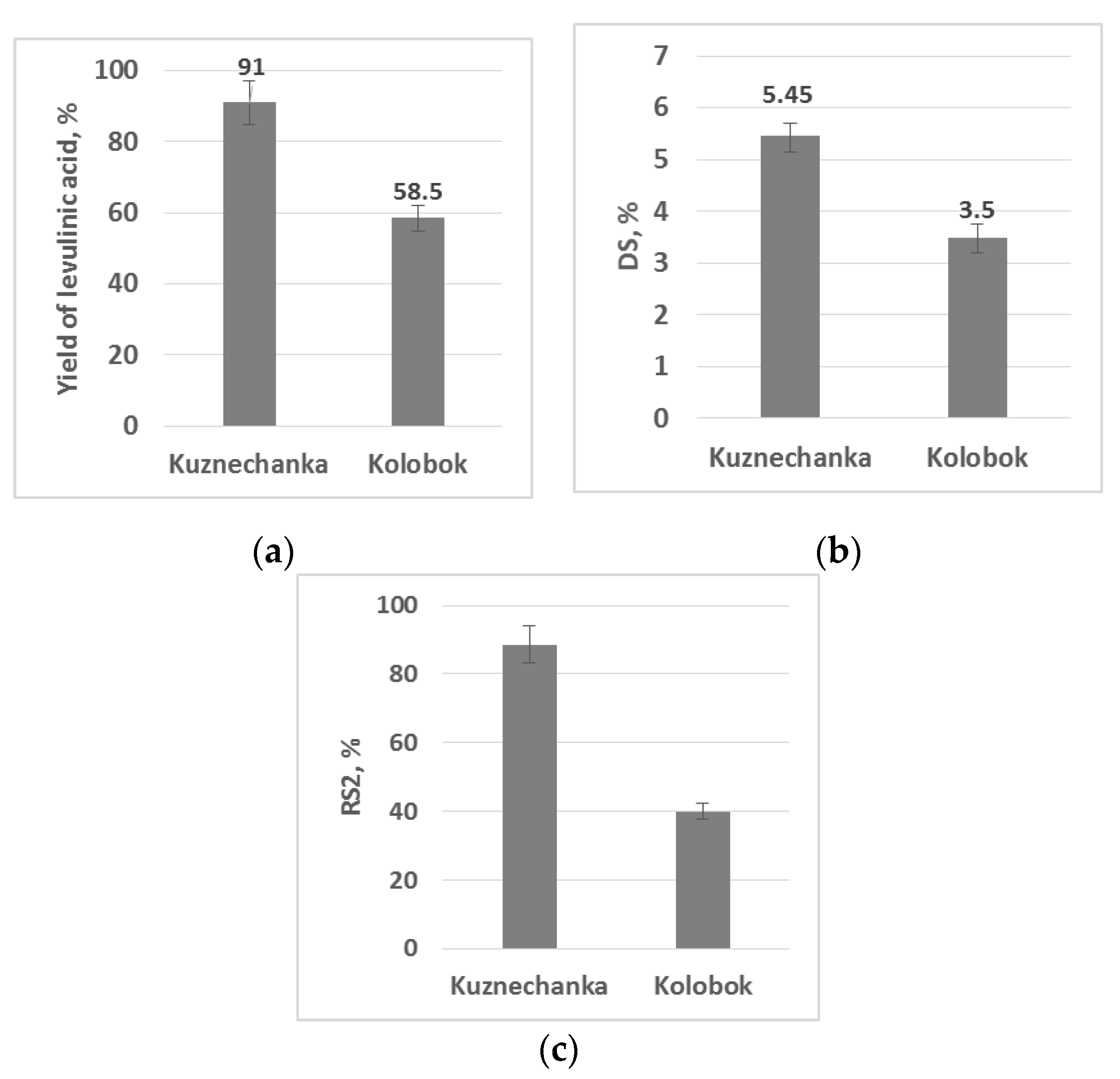
2.2. Starch from Potato Genotypes Kolobok and Kuznechanka in (Bio)Chemical Transformations
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Starch Isolation
4.3. Chemicals and Equipment
4.4. Starch to HMF
4.5. Starch to Levulinic Acid
4.6. Acylation of Starch
4.7. Resistant Starch
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Khlestkin, V.K.; Peltek, S.E.; Kolchanov, N.A. Review of direct chemical and biochemical transformations of starch. Carbohydr. Polym. 2018, 181, 460–476. [Google Scholar] [CrossRef] [PubMed]
- Khlestkin, V.K.; Erst, T.V. A practical guide to the starch granules’ morphology study by microscopy. Vavilovskii Zhurnal Genet. Sel. 2017, 21, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Khlestkin, V.K.; Erst, T.V.; Rozanova, I.V.; Efimov, V.M.; Khlestkina, E.K. Genetic loci determining potato starch yield and granule morphology revealed by genome-wide association study (GWAS). PeerJ 2020, 8, e10286. [Google Scholar] [CrossRef] [PubMed]
- Khlestkin, V.K.; Rozanova, I.V.; Efimov, V.M.; Khlestkina, E.K. Starch phosphorylation associated SNPs found by genome-wide association studies in the potato (Solanum tuberosum L.). BMC Genet. 2019, 20 (Suppl. 1). [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Schols, H.A.; Voragen, A.G.J. Differently sized granules from acetylated potato and sweet potato starches differ in the acetyl substitution pattern of their amylose populations. Carbohydr. Polym. 2004, 56, 219–226. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, J.; Suurs, P.; Schols, H.A.; Voragen, A.G.J. Granule size affects the acetyl substitution on amylopectin populations in potato and sweet potato starches. Carbohydr. Polym. 2005, 62, 333–337. [Google Scholar] [CrossRef]
- De Vetten, N.C.M.H.; Heeres, P. Method for Modifying the Size and/or Morphology of Starch Granules—EP 1473307A1; European Patent Office: Munich, Germany, 2004. [Google Scholar]
- Singh, N.; Kaur, L. Morphological, thermal, rheological and retrogradation properties of potato starch fractions varying in granule size. J. Sci. Food Agric. 2004, 84, 1241–1252. [Google Scholar] [CrossRef]
- Zhou, G.; Willett, J.L.; Craig, C.J.; Wu, Y.V. Effect of starch granule size on viscosity of starch-filled poly(hydroxy ester ether) composites. J. Polym. Environ. 2002, 8, 145–150. [Google Scholar] [CrossRef]
- Wang, C.; Tang, C.-H.; Fu, X.; Huang, Q.; Zhang, B. Granular size of potato starch affects structural properties, octenylsuccinic anhydride modification and flowability. Food Chem. 2016, 212, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Khlestkin, V.K.; Peltek, S.E.; Kolchanov, N.A. Target genes for development of potato (Solanum tuberosum L.) cultivars with desired starch properties. Sel’skokhozyaistvennaya Biol. 2017, 52, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.-B.; Lee, J.-W.; Choi, Y.-H.; Park, S.-M.; Chung, C.-H. Simple process for production of hydroxymethylfurfural from raw biomasses of girasol and potato tubers. Biomass Bioenergy 2012, 39, 484–488. [Google Scholar] [CrossRef]
- Fuentes-Zaragoza, E.; Sánchez-Zapata, E.; Sendra, E.; Sayas, E.; Navarro, C.; Fernández-López, J.; Pérez-Alvarez, J.A. Resistant starch as prebiotic: A review. Starch-Stärke 2011, 63, 406–415. [Google Scholar] [CrossRef]
- Vatanasuchart, N.; Niyomwit, B.; Wongkrajang, K. Resistant starch contents and the in vitro starch digestibility of Thai starchy foods. Kasetsart J. Nat. Sci. 2009, 43, 178–186. [Google Scholar]
- Tester, R.F.; Karkalas, J. The Effects of Environmental Conditions on the Structural Features and Physico-chemical Properties of Starches. Starch-Stärke 2001, 53, 513–519. [Google Scholar] [CrossRef]
- Sheets, R.L. Amylolysis of Eight Different Starches with Four Different Species of Alpha-Amylase. Master’s Thesis, Iowa State University, Ames, IA, USA, 2016. Available online: https://lib.dr.iastate.edu/etd/15188 (accessed on 4 January 2021).
- Bertolini, A.C.; Souza, E.S.; Nelson, J.E.; Huber, K.C. Composition and Reactivity of A- and B-type Starch Granules of Normal, Partial Waxy, and Waxy Wheat. Cereal Chem. 2003, 80, 544–549. [Google Scholar] [CrossRef]
- Svegmark, K.; Helmerson, K.; Nilsson, G.; Nilsson, P.-O.; Andersson, R.; Svensson, E. Comparison of potato amylopectin starches—Influence of year and variety. Carbohydr. Polym. 2002, 47, 331–340. [Google Scholar] [CrossRef]
- Yusupha, M.; Testera, R.F.; Ansella, R.; Snape, C.E. Composition and properties of starches extracted from tubers of different potato varieties grown under the same environmental conditions. Food Chem. 2003, 82, 283–289. [Google Scholar] [CrossRef]
- Lai, L.; Zhang, Y. The Production of 5-Hydroxymethylfurfural from Fructose in Isopropyl Alcohol: A Green and Efficient System. ChemSusChem 2011, 4, 1745–1748. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Dumont, M.-J. Levulinic Acid Production from Starch Using Microwave and Oil Bath Heating: A Kinetic Modeling Approach. Ind. Eng. Chem. Res. 2016, 55, 8941–8949. [Google Scholar] [CrossRef]
- Boruczkowska, H.; Boruczkowski, T.; Gubica, T.; Anioł, M.; Tomaszewska-Ciosk, E. Analysis of the chemical structure of insoluble products of enzymatic esterification of starch and transesterification of acetylated starch with oleic acid by solid-state CP/MAS 13C NMR. Starch-Stärke 2016, 68, 1180–1186. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Yield, % | Feret’s Diameter, µm | Circularity | Aspect Ratio | Phosphorus, % |
|---|---|---|---|---|---|
| Kuznechanka | 13.78 | 39.672 | 0.8493 | 1.31 | 0.080 |
| Kolobok | 11.62 | 32.527 | 0.8464 | 1.32 | 0.092 |
| Compounds | Compared Signals | Number of H-Atoms | Integral Units | Units per H-Atom | Ratio | Amount in NMR Sample, Mole | Yield in Reaction, mg | Yield, % 1 |
|---|---|---|---|---|---|---|---|---|
| HMDS | SiCH3 | 18 | 8.073 | 0.4485 | 1 | 9.41 × 10−6 | --- | --- |
| Levulinic acid | CH2 | 2 | 1.027 | 0.5135 | 1.15 | 10.77 × 10−6 | 62.5 | 97 |
| Compounds | Compared Signals | Number of H-Atoms | Integral Units | Units per H-Atom | Ratio | DS, % |
|---|---|---|---|---|---|---|
| Starch | CHOH | 1 | 17.5 | 17.5 | 1 | 5.7 |
| CH3 | 3 | 3 | 1 | 0.057 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khlestkin, V.; Eltsov, I. Different Reactivity of Raw Starch from Diverse Potato Genotypes. Molecules 2021, 26, 226. https://doi.org/10.3390/molecules26010226
Khlestkin V, Eltsov I. Different Reactivity of Raw Starch from Diverse Potato Genotypes. Molecules. 2021; 26(1):226. https://doi.org/10.3390/molecules26010226
Chicago/Turabian StyleKhlestkin, Vadim, and Ilia Eltsov. 2021. "Different Reactivity of Raw Starch from Diverse Potato Genotypes" Molecules 26, no. 1: 226. https://doi.org/10.3390/molecules26010226
APA StyleKhlestkin, V., & Eltsov, I. (2021). Different Reactivity of Raw Starch from Diverse Potato Genotypes. Molecules, 26(1), 226. https://doi.org/10.3390/molecules26010226