
Proteomic Analysis of Zeb1 Interactome in Breast Carcinoma Cells


 , ,
, , 

Abstract
:1. Introduction
2. Results and Discussion
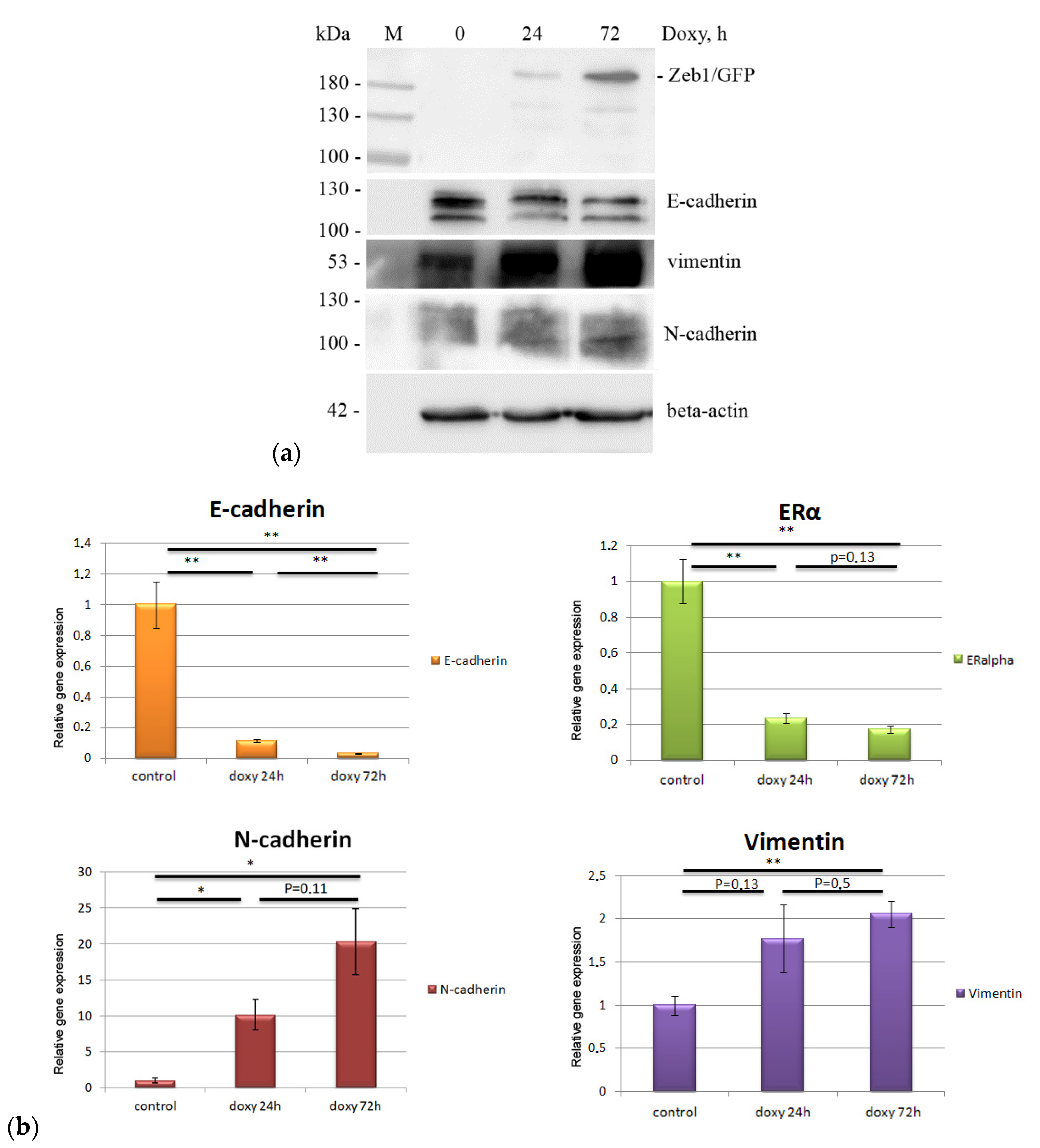
2.1. EMT Induction
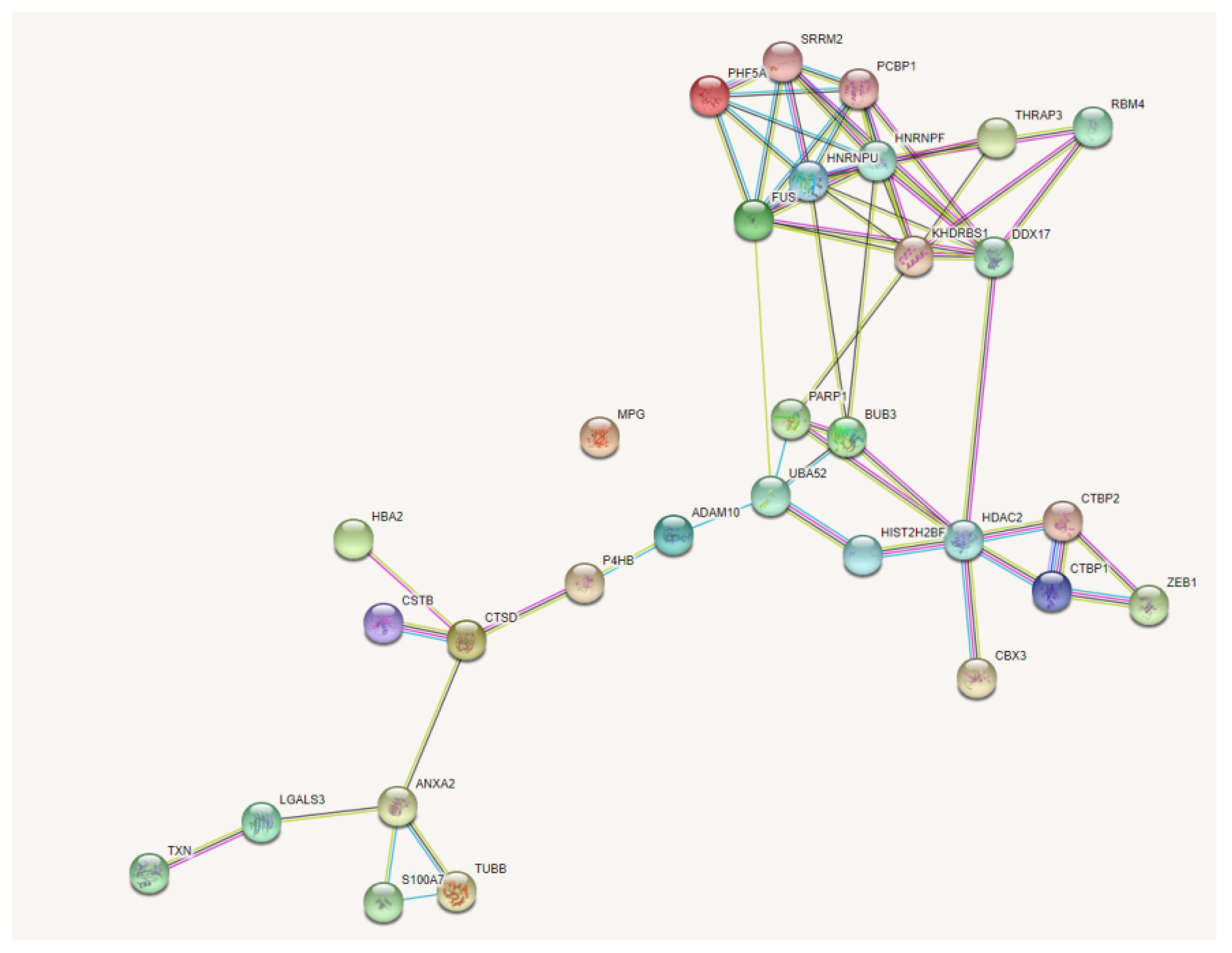
2.2. Isolation and Analysis of Zeb1 Interactome
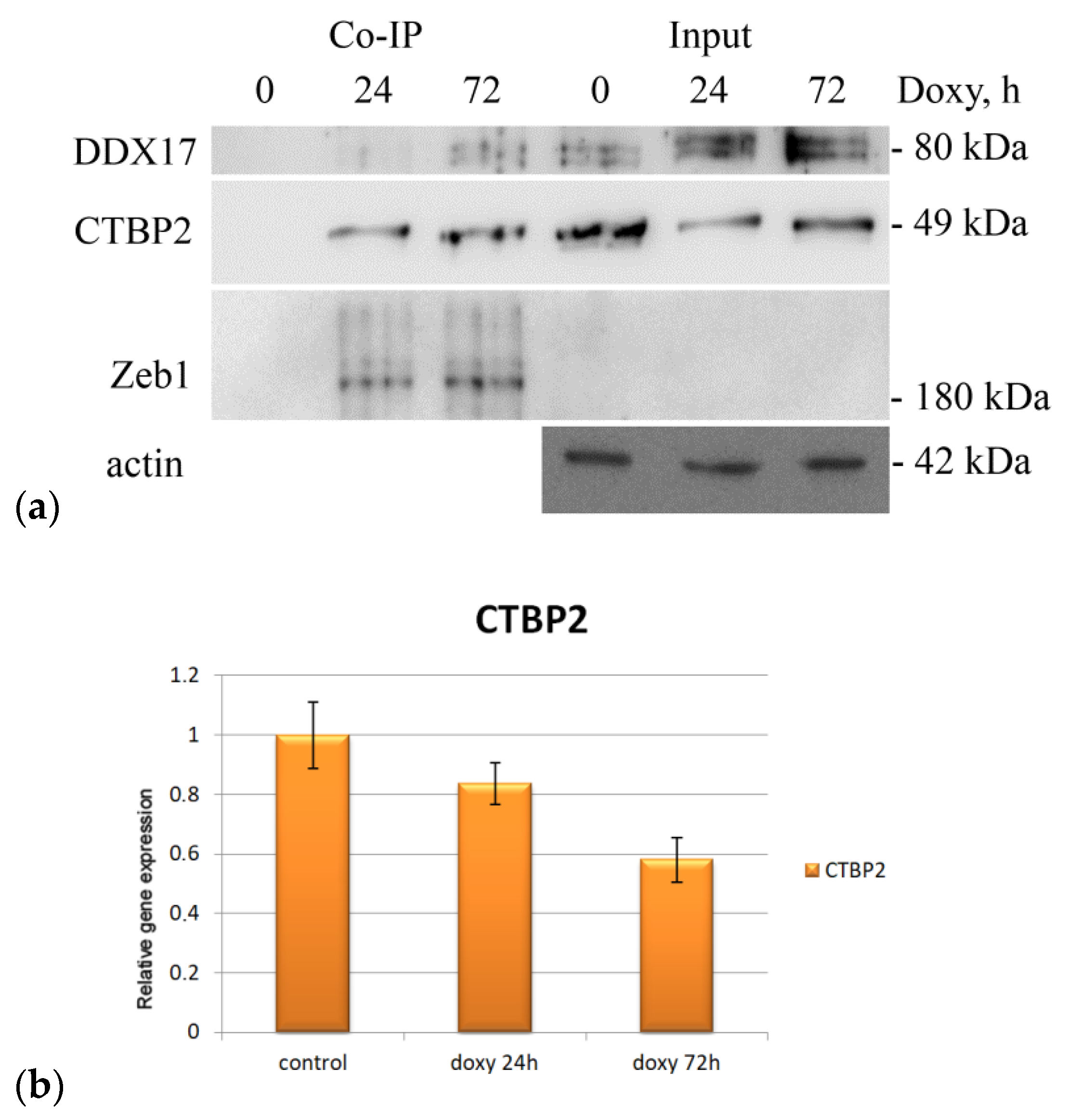
2.2.1. RNA Helicase DDX17
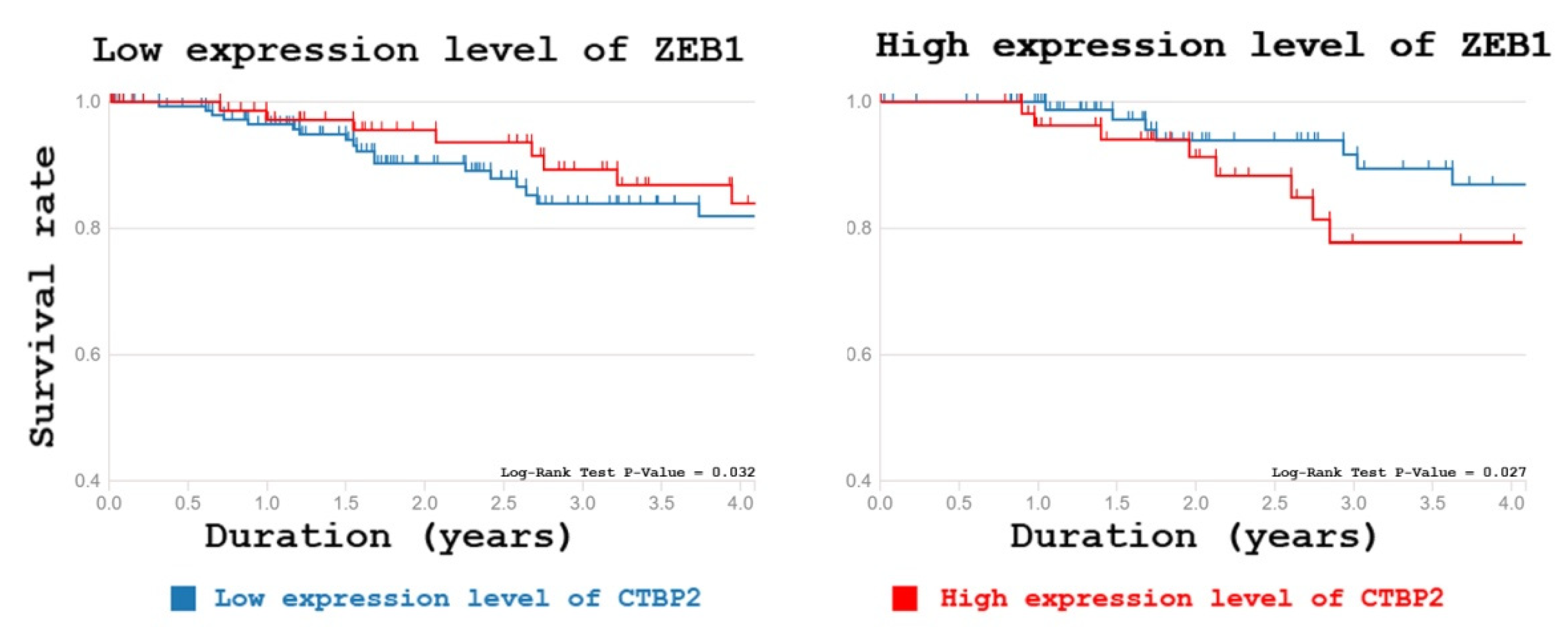
2.2.2. C-Terminal-Binding Protein 2
2.3. Mass Spectrometry Data Verification
3. Materials and Methods
3.1. Cell Culture
3.2. Wound Healing Assay
3.3. Co-Immunoprecipitation
3.4. Sample Preparation and LC-MALDI Mass Spectrometry
3.5. Western Blotting
3.6. Real-Time Polymerase Chain Reaction
3.7. Statistical Analysis
3.8. Bioinformatic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Pozdnyakov, D.Y.; Shuvalov, O.Y.; Barlev, N.A.; Mittenberg, A.G. Role of Zeb1 EMT-TF in metastasis and carcinogenesis. Tsitologiia 2019, 61, 915–925. (In Russian) [Google Scholar] [CrossRef]
- Pozdnyakov, D.Y.; Shuvalov, O.Y.; Barlev, N.A.; Mittenberg, A.G. Post-transcriptional regulation of Zeb1 EMT-TF activity in human breast carcinomae. Tsitologiia 2020, 62, 3–15. (In Russian) [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeone, P.; Trerotola, M.; Franck, J.; Cardon, T.; Marchisio, M.; Fournier, I.; Salzet, M.; Maffia, M.; Vergara, D. The multiverse nature of epithelial to mesenchymal transition. Semin. Cancer Biol. 2019, 58, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Roxanis, I. Occurrence and significance of epithelial-mesenchymal transition in breast cancer. J. Clin. Pathol. 2013, 66, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, B.P. Epithelial-mesenchymal Transition—A Hallmark of Breast Cancer Metastasis. Cancer Hallm. 2013, 1, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Dave, B.; Mittal, V.; Tan, N.M.; Chang, J.C. Epithelial-mesenchymal transition, cancer stem cells and treatment resistance. Breast Cancer Res. 2012, 14, 202. [Google Scholar] [CrossRef] [Green Version]
- Mallini, P.; Lennard, T.; Kirby, J.; Meeson, A. Epithelial-to-mesenchymal transition: What is the impact on breast cancer stem cells and drug resistance. Cancer Treat. Rev. 2014, 40, 341–348. [Google Scholar] [CrossRef]
- Barrallo-Gimeno, A.; Nieto, M.A. The Snail genes as inducers of cell movement and survival: Implications in development and cancer. Development 2005, 132, 3151–3161. [Google Scholar] [CrossRef] [Green Version]
- Eger, A.; Aigner, K.; Sonderegger, S.; Dampier, B.; Oehler, S.; Schreiber, M.; Berx, G.; Cano, A.; Beug, H.; Foisner, R. DeltaEF1 is a transcriptional repressor of E-cadherin and regulates epithelial plasticity in breast cancer cells. Oncogene 2005, 24, 2375–2385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirakihara, T.; Saitoh, M.; Miyazono, K. Differential regulation of epithelial and mesenchymal markers by deltaEF1 proteins in epithelial mesenchymal transition induced by TGF-beta. Mol. Biol. Cell. 2007, 18, 3533–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef] [Green Version]
- Tseng, L.M.; Hsu, N.C.; Chen, S.C.; Lu, Y.S.; Lin, C.H.; Chang, D.Y.; Li, H.; Lin, Y.C.; Chang, H.K.; Chao, T.C.; et al. Distant metastasis in triple-negative breast cancer. Neoplasia 2013, 60, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Pastushenko, I.; Brisebarre, A.; Sifrim, A.; Fioramonti, M.; Revenco, T.; Boumahdi, S.; van Keymeulen, A.; Brown, D.; Moers, V.; Lemaire, S.; et al. Identification of the tumour transition states occurring during EMT. Nature 2018, 556, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Tulchinsky, E.; Demidov, O.; Kriajevska, M.; Barlev, N.A.; Imyanitov, E. EMT: A mechanism for escape from EGFR-targeted therapy in lung cancer. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 29–39. [Google Scholar] [CrossRef]
- Caramel, J.; Ligier, M.; Puisieux, A. Pleiotropic Roles for ZEB1 in Cancer. Cancer Res. 2018, 78, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.S.; Gao, W.; Chan, J.Y. Transcription regulation of E-cadherin by zinc finger E-box binding homeobox proteins in solid tumors. Biomed. Res. Int. 2014, 2014, 921564. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Jordaan, G.; Srivastava, M.K.; Dubinett, S.; Sharma, S.; Sharma, S. Effect of epigenetic histone modifications on E-cadherin splicing and expression in lung cancer. Am. J. Cancer Res. 2013, 3, 374–389. [Google Scholar]
- Zhang, J.; Zhou, C.; Jiang, H.; Liang, L.; Shi, W.; Zhang, Q.; Sun, P.; Xiang, R.; Wang, Y.; Yang, S. ZEB1 induces ER-α promoter hypermethylation and confers antiestrogen resistance in breast cancer. Cell Death Dis. 2017, 8, e2732. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janknecht, R. Multi-talented DEAD-box proteins and potential tumor promoters: p68 RNA helicase (DDX5) and its paralog, p72 RNA helicase (DDX17). Am. J. Transl. Res. 2010, 2, 223–234. [Google Scholar] [PubMed]
- Dutertre, M.; Gratadou, L.; Dardenne, E.; Germann, S.; Samaan, S.; Lidereau, R.; Driouch, K.; de la Grange, P.; Auboeuf, D. Estrogen regulation and physiopathologic significance of alternative promoters in breast cancer. Cancer Res. 2010, 70, 3760–3770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Guo, X.; Lv, F.; Xu, Y.; Gao, G. p72 DEAD box RNA helicase is required for optimal function of the zinc-finger antiviral protein. Proc. Natl. Acad. Sci. USA 2008, 105, 4352–4357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, M.; Triboulet, R.; Mohseni, M.; Schlegelmilch, K.; Shrestha, K.; Camargo, F.D.; Gregory, R.I. Hippo signaling regulates microprocessor and links cell-density-dependent miRNA biogenesis to cancer. Cell 2014, 156, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Shen, X.; Yang, W.; Zhang, Y.; Liu, C.; Huang, T. ZEB1 stimulates breast cancer growth by up-regulating hTERT expression. Biochem. Biophys. Res. Commun. 2018, 495, 2505–2511. [Google Scholar] [CrossRef]
- Wang, J.; Lee, S.; Teh, C.E.; Bunting, K.; Ma, L.; Shannon, M.F. The transcription repressor, ZEB1, cooperates with CtBP2 and HDAC1 to suppress IL-2 gene activation in T cells. Int. Immunol. 2009, 21, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xiao, Z.; Zheng, J.; Wu, J.; Hu, X.L.; Yang, X.; Shen, Q. ZEB1 Represses Neural Differentiation and Cooperates with CTBP2 to Dynamically Regulate Cell Migration during Neocortex Development. Cell Rep. 2019, 27, 2335–2353. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.J.; Kuppuswamy, M.; Vijayalingam, S.; Chinnadurai, G. Interaction of ZEB and histone deacetylase with the PLDLS-binding cleft region of monomeric C-terminal binding protein 2. BMC Mol Biol. 2009, 10, 89. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Sun, Y.; Li, H.; Shao, Y.; Zhao, D.; Yu, W.; Fu, J. C-terminal binding protein-2 promotes cell proliferation and migration in breast cancer via suppression of p16INK4A. Oncotarget 2017, 8, 26154–26168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yang, C.S.; Varelas, X.; Monti, S. Altered RNA editing in 3’ UTR perturbs microRNA-mediated regulation of oncogenes and tumor-suppressors. Sci. Rep. 2016, 6, 23226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birts, C.N.; Harding, R.; Soosaipillai, G.; Halder, T.; Azim-Araghi, A.; Darley, M.; Cutress, R.I.; Bateman, A.C.; Blaydes, J.P. Expression of CtBP family protein isoforms in breast cancer and their role in chemoresistance. Biol. Cell 2010, 103, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Hao, D.; Wang, L.; Li, J.; Meng, Y.; Li, P.; Wang, Y.; Zhang, C.; Zhou, H.; Gardner, K.; et al. CtBP promotes metastasis of breast cancer through repressing cholesterol and activating TGF-β signaling. Oncogene 2019, 38, 2076–2091. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.S.; Park, S.; Yi, D.I.; Shin, J.H.; Hernandez, S.G.; Hewitt, S.M.; Nicklaus, M.C.; Peach, M.L.; Guasch, L.; Tang, B.; et al. Epigenetic re-wiring of breast cancer by pharmacological targeting of C-terminal binding protein. Cell Death Dis. 2019, 10, 689. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Sekiya, M.; Kainoh, K.; Matsuda, T.; Iwasaki, H.; Osaki, Y.; Sugano, Y.; Suzuki, H.; Takeuchi, Y.; Miyamoto, T.; et al. Transcriptional co-repressor CtBP2 orchestrates epithelial-mesenchymal transition through a novel transcriptional holocomplex with OCT1. Biochem. Biophys. Res. Commun. 2020, 523, 354–360. [Google Scholar] [CrossRef]
- Zheng, X.; Song, T.; Dou, C.; Jia, Y.; Liu, Q. CtBP2 is an independent prognostic marker that promotes GLI1 induced epithelial-mesenchymal transition in hepatocellular carcinoma. Oncotarget 2015, 6, 3752–3769. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Xiang, Z.; Xiong, F.; Yan, B.; Huang, Q. C-terminal binding protein-2 is a prognostic marker for lung adenocarcinomas. Medicine 2020, 99, e20076. [Google Scholar] [CrossRef]
- Soule, H.D.; Vazquez, J.; Long, A.; Albert, S.; Brennan, M. A human cell line from a pleural effusion derived from a breast carcinoma. J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef]
- Wan Makhtar, W.R.; Browne, G.; Karountzos, A.; Stevens, C.; Alghamdi, Y.; Bottrill, A.R.; Mistry, S.; Smith, E.; Bushel, M.; Pringle, J.H.; et al. Short stretches of rare codons regulate translation of the transcription factor ZEB2 in cancer cells. Oncogene 2017, 36, 6640–6648. [Google Scholar] [CrossRef] [Green Version]
- Manshouri, R.; Coyaud, E.; Kundu, S.T.; Peng, D.H.; Stratton, S.A.; Alton, K.; Bajaj, R.; Fradette, J.J.; Minelli, R.; Peoples, M.D.; et al. ZEB1/NuRD complex suppresses TBC1D2b to stimulate E-cadherin internalization and promote metastasis in lung cancer. Nat. Commun. 2019, 10, 5125. [Google Scholar] [CrossRef]
- Zhang, P.; Wei, Y.; Wang, L.; Debeb, B.G.; Yuan, Y.; Zhang, J.; Yuan, J.; Wang, M.; Chen, D.; Sun, Y.; et al. ATM-mediated stabilization of ZEB1 promotes DNA damage response and radioresistance through CHK1. Nat. Cell Biol. 2014, 16, 864–875. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′-3′ Primer Sequence |
|---|---|
| E-cadherin | F: CTTCTGCTGATCCTGTCTGATG |
| R: TGCTGTGAAGGGAGATGTATTG | |
| GAPDH | F: GTCTCCTCTGACTTCAACAGCG |
| R: ACCACCCTGTTGCTGTAGCCAA | |
| ZEB1 | F: GGCATACACCTACTCAACTACGG |
| R: TGGGCGGTGTAGAATCAGAGTC | |
| CtBP2 | F: CGTTCTCAGAGCTGGGATGC |
| R: TCTGCTGTGCCATACGTCAG | |
| N-cadherin | F: AGCCCGGTTTCATTTGAGGG |
| R: TTGAGGGCATTGGGATCGTC | |
| Vimentin | F: TGTCCAAATCGATGTGGATGTTTC |
| R: TTGTACCATTCTTCTGCCTCCTG | |
| ERα | F: TGATGAAAGGTGGGATACGA |
| R: AAGGTTGGCAGCTCTCATGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parfenyev, S.E.; Shabelnikov, S.V.; Pozdnyakov, D.Y.; Gnedina, O.O.; Adonin, L.S.; Barlev, N.A.; Mittenberg, A.G. Proteomic Analysis of Zeb1 Interactome in Breast Carcinoma Cells. Molecules 2021, 26, 3143. https://doi.org/10.3390/molecules26113143
Parfenyev SE, Shabelnikov SV, Pozdnyakov DY, Gnedina OO, Adonin LS, Barlev NA, Mittenberg AG. Proteomic Analysis of Zeb1 Interactome in Breast Carcinoma Cells. Molecules. 2021; 26(11):3143. https://doi.org/10.3390/molecules26113143
Chicago/Turabian StyleParfenyev, Sergey E., Sergey V. Shabelnikov, Danila Y. Pozdnyakov, Olga O. Gnedina, Leonid S. Adonin, Nickolai A. Barlev, and Alexey G. Mittenberg. 2021. "Proteomic Analysis of Zeb1 Interactome in Breast Carcinoma Cells" Molecules 26, no. 11: 3143. https://doi.org/10.3390/molecules26113143
APA StyleParfenyev, S. E., Shabelnikov, S. V., Pozdnyakov, D. Y., Gnedina, O. O., Adonin, L. S., Barlev, N. A., & Mittenberg, A. G. (2021). Proteomic Analysis of Zeb1 Interactome in Breast Carcinoma Cells. Molecules, 26(11), 3143. https://doi.org/10.3390/molecules26113143