
Protective Effects of Thymoquinone, an Active Compound of Nigella sativa, on Rats with Benzo(a)pyrene-Induced Lung Injury through Regulation of Oxidative Stress and Inflammation

 , and
, and
Abstract
:1. Introduction
2. Results
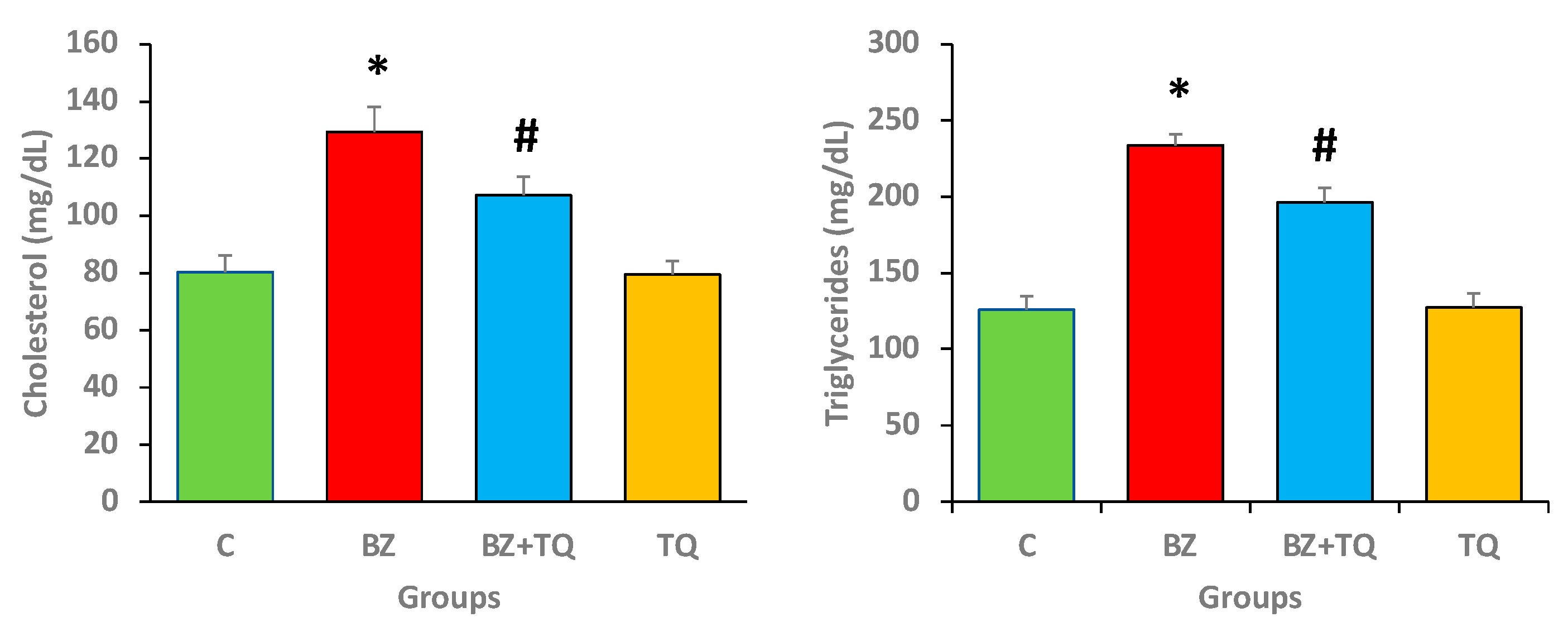
2.1. Effect of Thymoquinone on Lipid Profile Parameters
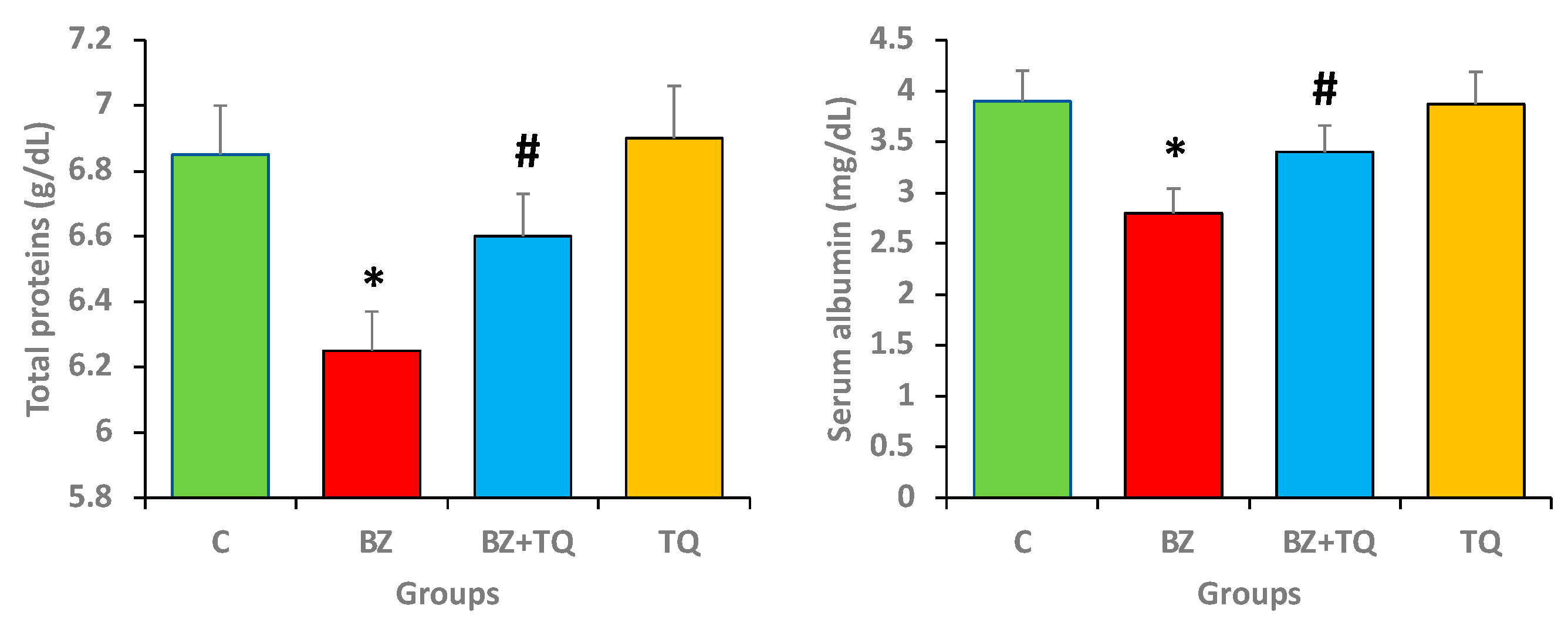
2.2. Effect of TQ on Total Protein and Albumin Levels
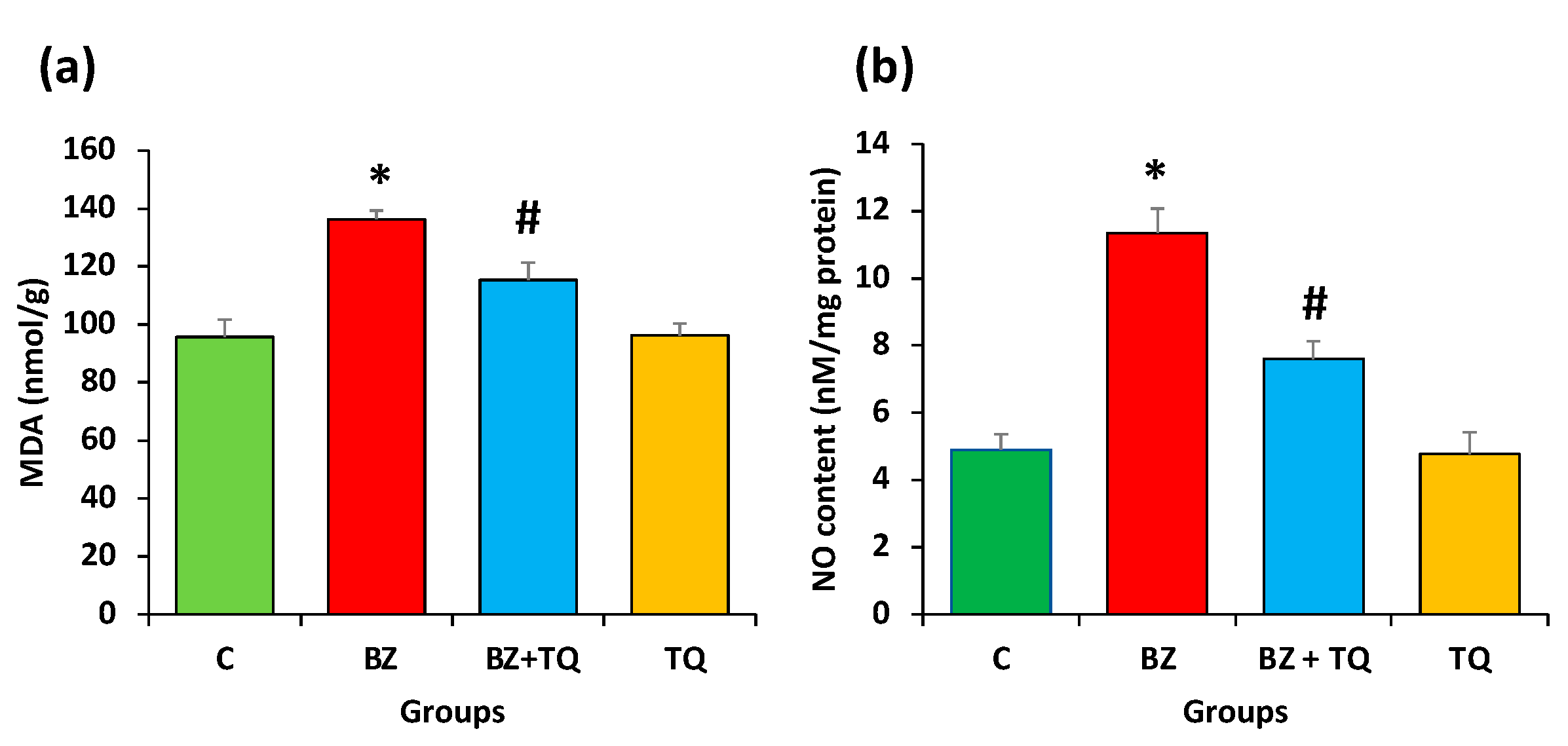
2.3. Effect of Thymoquinone on MDA and NO Levels
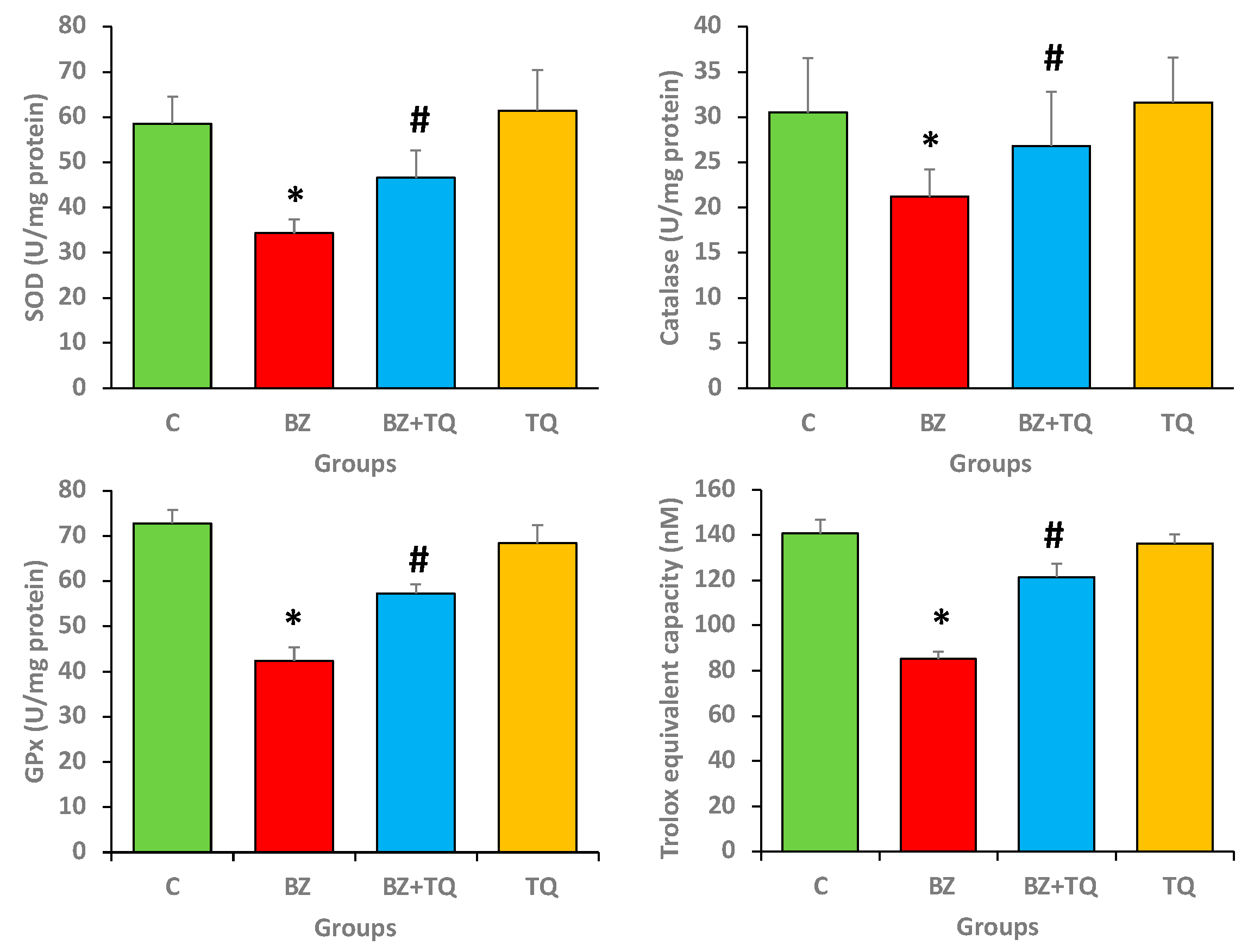
2.4. Effect of Thymoquinone on Antioxidant Enzyme Levels
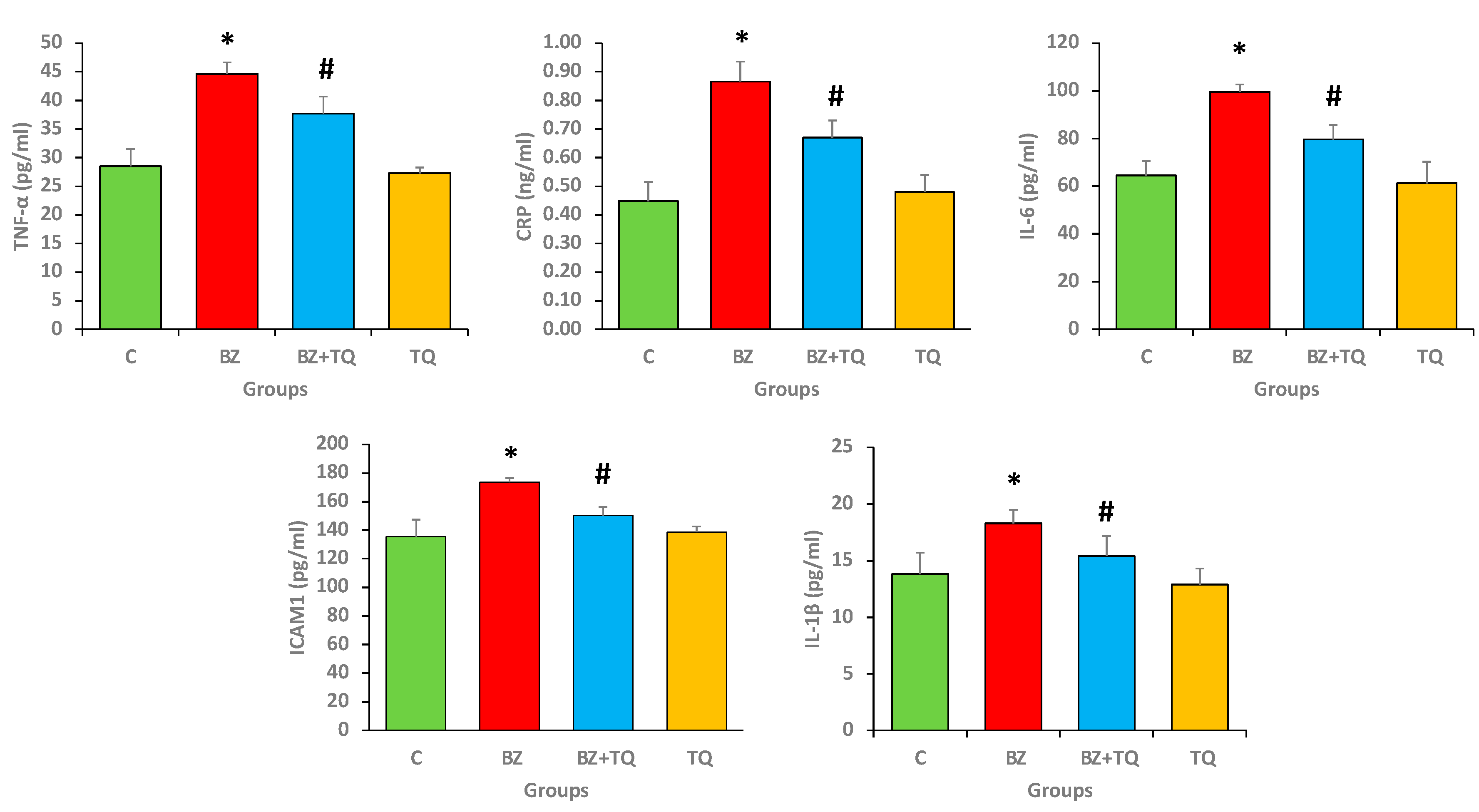
2.5. Effect of B(a)P and TQ on Anti-Inflammatory Markers
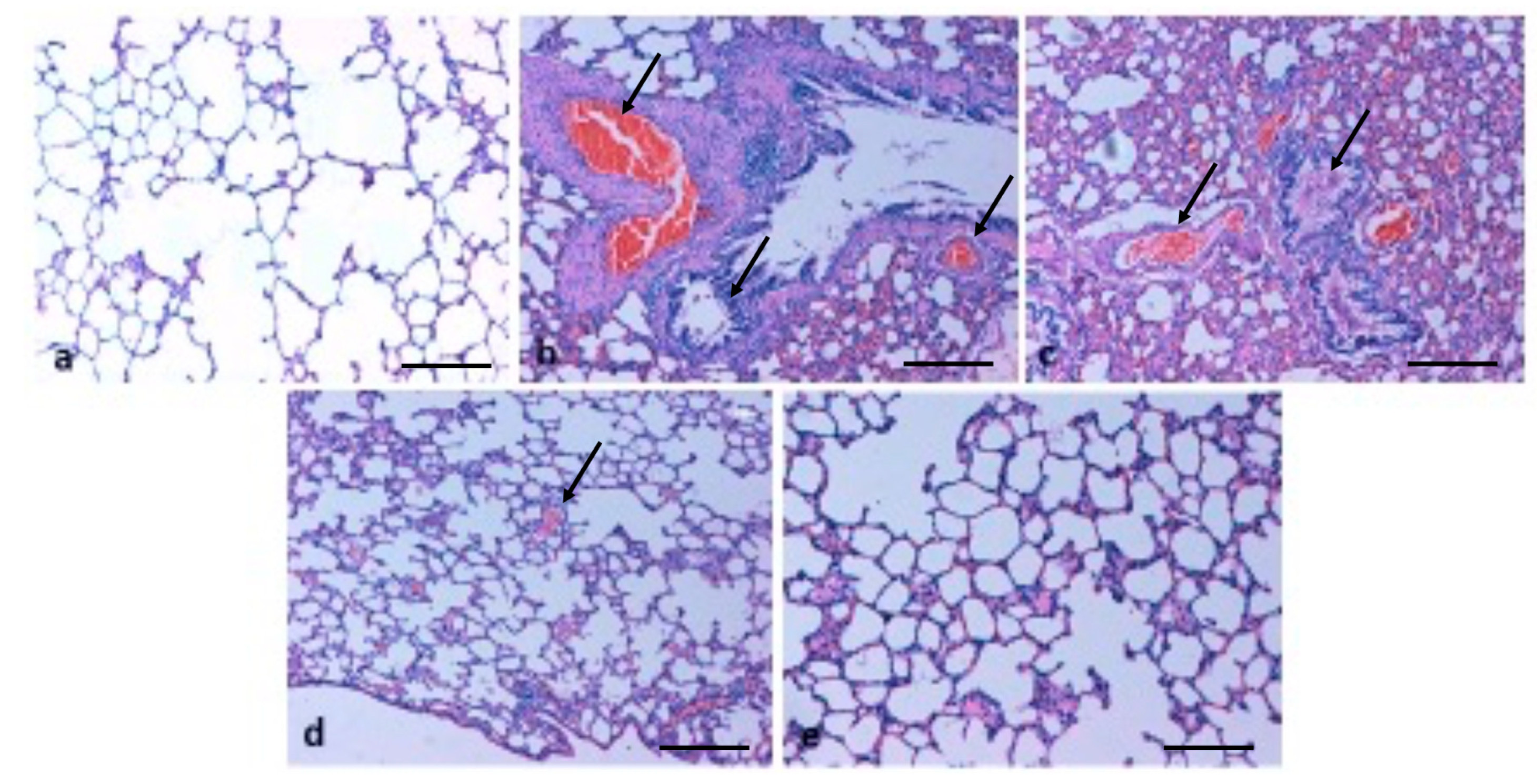
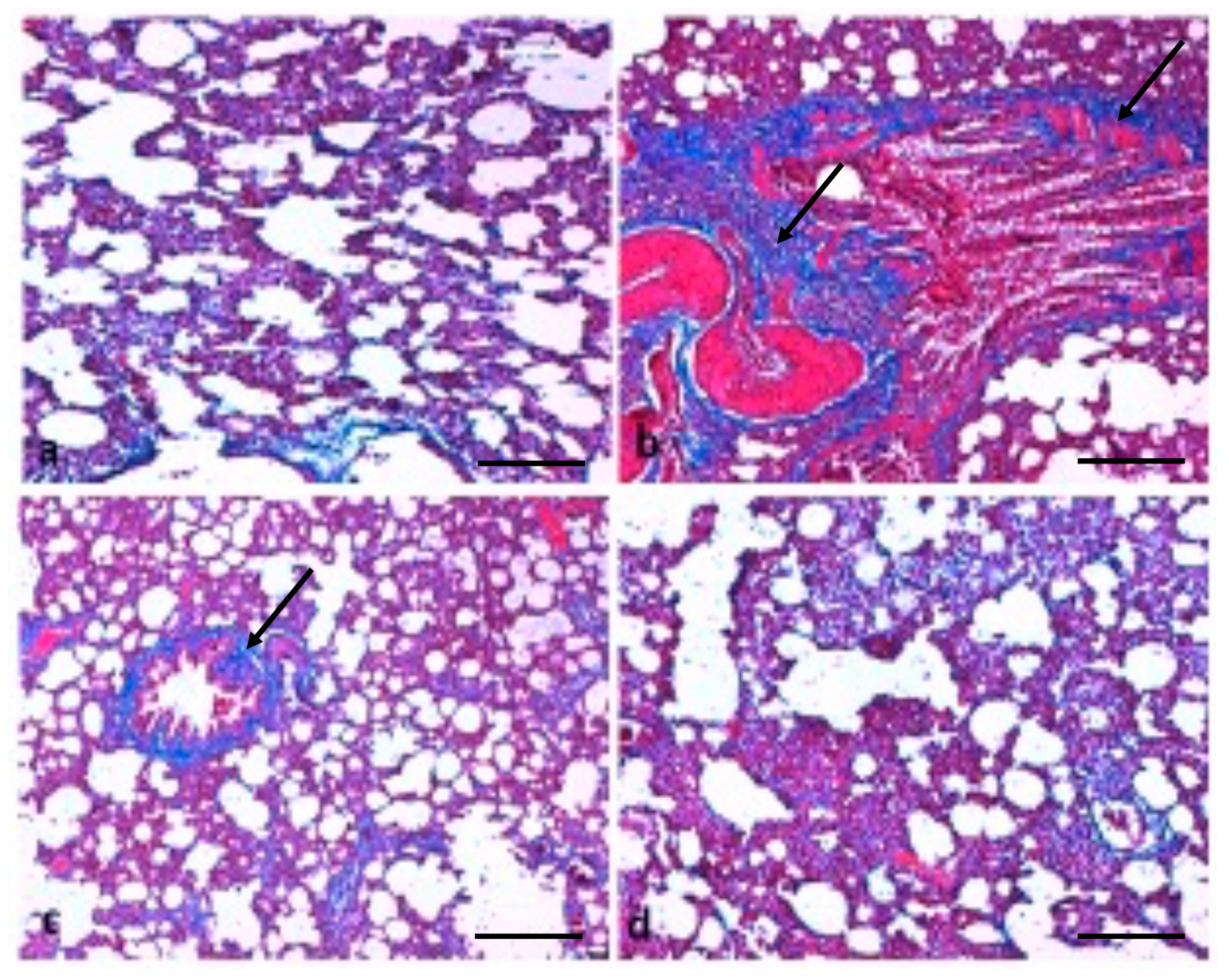
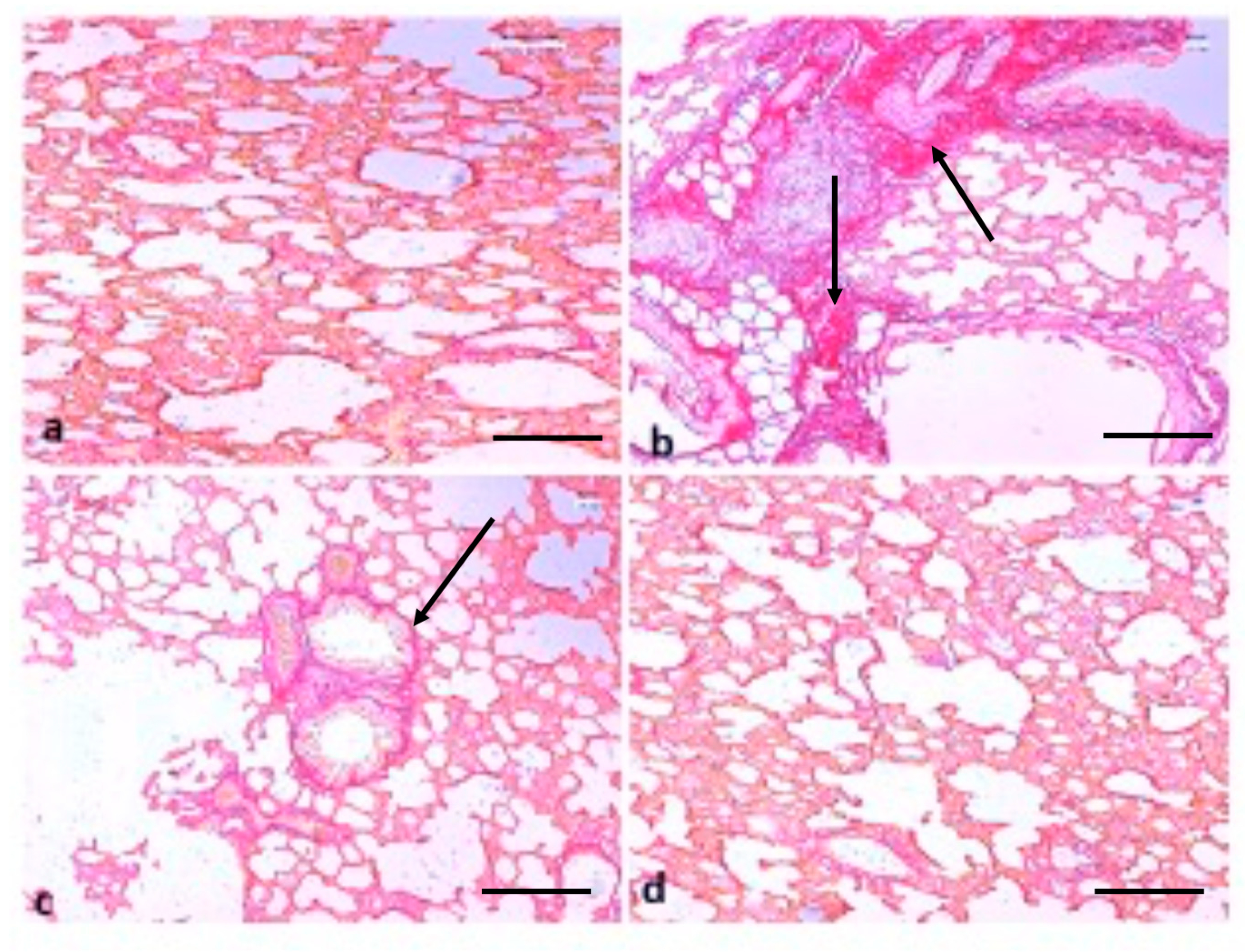
2.6. Effect of TQ on Lung Architecture
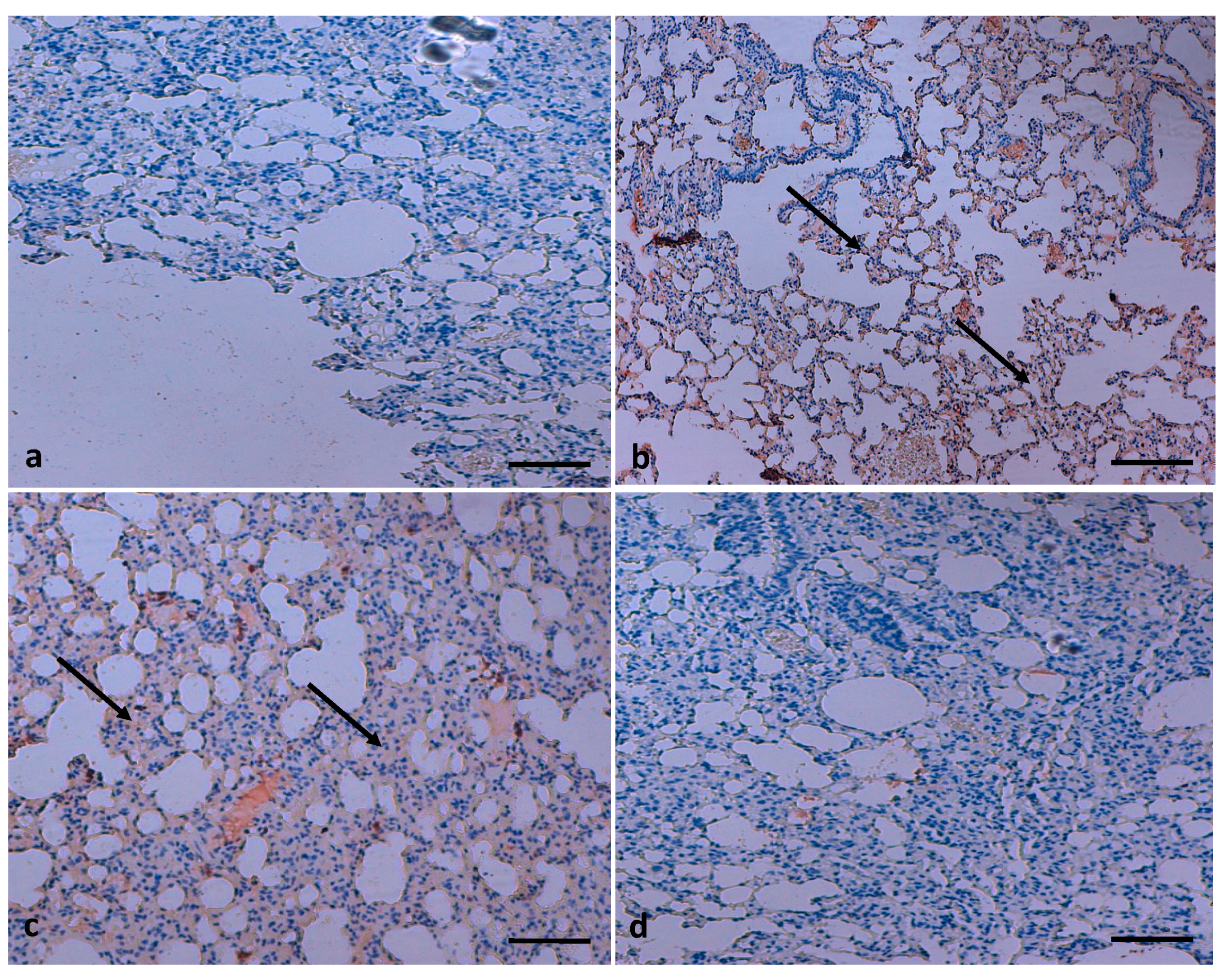
3. TQ Reduces NF-κB Expression in Lung Tissue
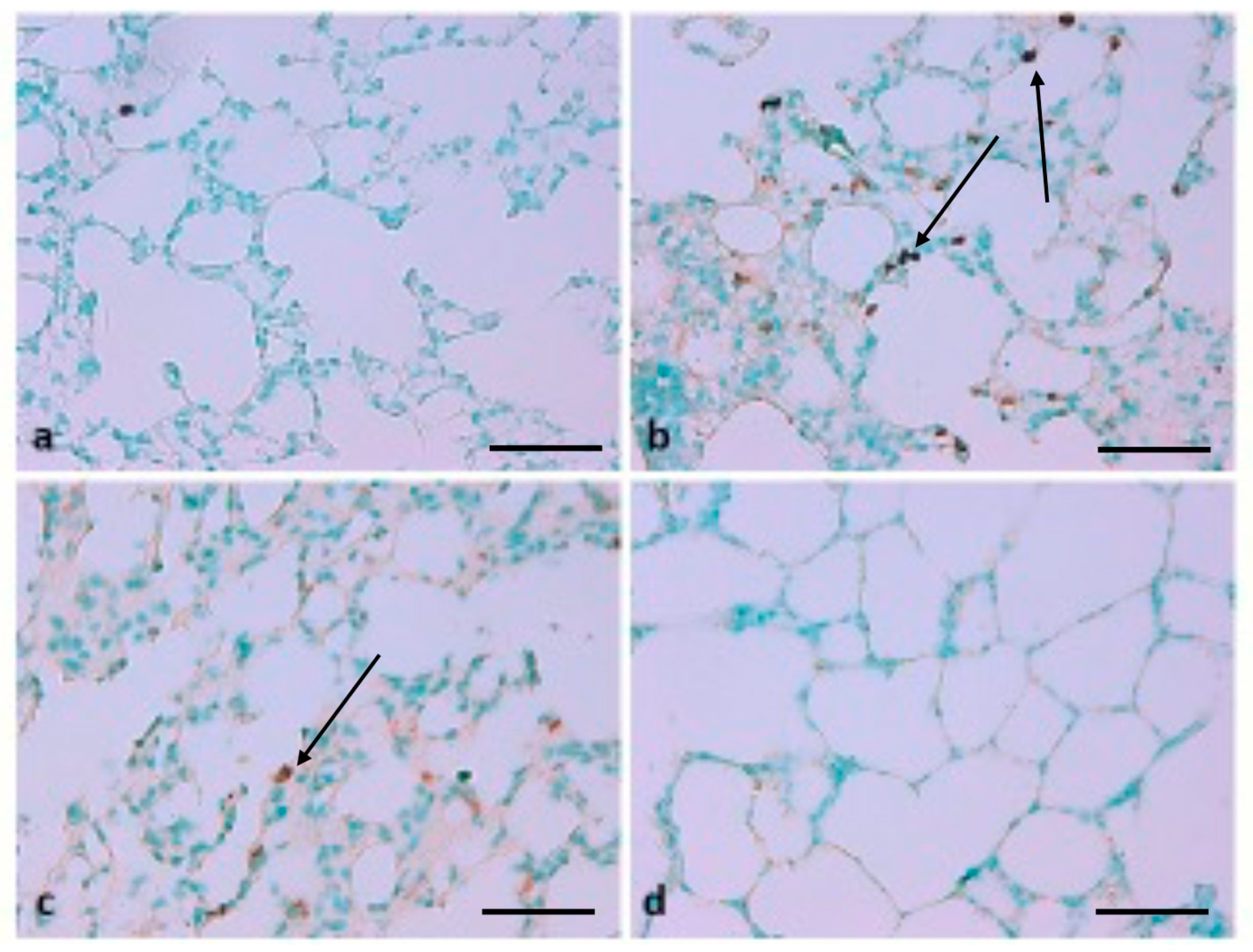
Effect of TQ on Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick End Labelling (TUNEL)-Positive Cells
4. Discussion
5. Materials and Methods
5.1. Chemicals
5.2. Animals
5.3. Animal Ethics
5.4. Animal Grouping and Treatment Plan
5.5. Measurement of Lipid Profile
5.6. Malondialdehyde (MDA) and NO Contents
5.7. Measurement of Antioxidant Enzymes
5.8. Total Antioxidant Capacity Measurement
5.9. Measurement of Inflammatory Markers
5.10. Haematoxylin and Eosin (H&E) Staining
5.11. Masson’s Trichrome Staining
5.12. Picro Sirius Red Staining
5.13. Terminal Deoxynucleotidyl Transferase-Mediated dUTP-Biotin Nick-End Labelling (TUNEL) Assay
5.14. Expressional Evaluation of NF-κB Protein
5.15. Statistical Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McCarthy, W.J.; Meza, R.; Jeon, J.; Moolgavkar, S.H. Chapter 6: Lung Cancer in Never Smokers: Epidemiology and Risk Prediction Models. Risk Anal. 2012, 32, S69–S84. [Google Scholar] [CrossRef] [Green Version]
- Piccardo, M.T.; Stella, A.; Valerio, F. Is the smokers exposure to environmental tobacco smoke negligible? Environ. Health 2010, 9, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, C.D.; Kim, J.W.; Ha, J.H.; Kim, S.J.; Lee, S.H.; Kim, I.K. Chemopreventive effect of phosphodieasterase-4 inhibition in ben-zo(a)pyrene-induced murine lung cancer model. Exp. Lung Res. 2014, 40, 500–506. [Google Scholar] [CrossRef]
- Jahangir, T.; Sultana, S. Benzo(a)pyrene-induced genotoxicity: Attenuation by farnesol in a mouse model. J. Enzym. Inhib. Med. Chem. 2008, 23, 888–894. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.P.; Ramos, K.S. Impact of cellular metabolism on the biological effects of benzo[a]pyrene and related hydrocarbons†. Drug Metab. Rev. 2001, 33, 1–35. [Google Scholar] [CrossRef]
- Wang, S.; Zang, W.; Yang, Y.; Zhang, Q.; Zhao, M.; Gao, Z. Tanshinone IIA and Baicalin inhibiting the formation of benzo [a] pyrene and benzo [a] pyrene induced cytotoxicity: Correlation with scavenging free radical. Environ. Toxicol. Pharmacol. 2013, 36, 403–410. [Google Scholar] [CrossRef]
- Chung, C.-K.; Jung, M.-E. Ethanol Fraction of Aralia elata Seemann Enhances Antioxidant Activity and Lowers Serum Lipids in Rats When Administered with Benzo(.ALPHA.)pyrene. Biol. Pharm. Bull. 2003, 26, 1502–1504. [Google Scholar] [CrossRef] [Green Version]
- Yuvaraj, S.; Premkumar, V.G.; Vijayasarathy, K.; Gangadaran, S.G.D.; Sachdanandam, P. Ameliorating effect of coenzyme Q10, riboflavin and niacin in tamoxifen-treated postmenopausal breast cancer patients with special reference to lipids and lipoproteins. Clin. Biochem. 2007, 40, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Moldoveanu, B.; Otmishi, P.; Jani, P.; Walker, J.; Sarmiento, X.; Guardiola, J.; Saad, M.; Yu, J. Inflammatory mechanisms in the lung. J. Inflamm. Res. 2008, 2, 1–11. [Google Scholar]
- Benihoud, K.; Esselin, S.; Descamps, D.; Jullienne, B.; Salone, B.; Bobé, P.; Bonardelle, D.; Connault, E.; Opolon, P.; Saggio, I.; et al. Erratum: Respective roles of TNF-α and IL-6 in the immune response-elicited by adenovirus-mediated gene transfer in mice. Gene Ther. 2007, 14, 551. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Luedde, T.; Schwabe, R.F. NF-kB in the liver-linking injury, fibrosis and hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Luo, W. Exploration of pro-apoptotic effect of Thymoquinone on oral squamous cell carcinoma cells through PI3K/Akt signaling pathway. Cell. Mol. Biol. 2019, 65, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Pelegrin, S.; Galtier, F.; Chalançon, A.; Gagnol, J.P.; Barbanel, A.M.; Pélissier, Y.; Larroque, M.; Lepape, S.; Faucanié, M.; Gabillaud, I. Effects of Nigella sativa seeds (black cumin) on insulin secretion and lipid profile: A pilot study in healthy volunteers. Br. J. Clin. Pharmacol. 2019, 85, 1607–1611. [Google Scholar] [CrossRef] [PubMed]
- Zeinvand-Lorestani, H.; Nili-Ahmadabadi, A.; Balak, F.; Hasanzadeh, G.; Sabzevari, O. Protective role of thymo-quinone against paraquat-induced hepatotoxicity in mice. Pestic. Biochem. Physiol. 2018, 148, 16–21. [Google Scholar] [CrossRef]
- Guan, D.; Li, Y.; Peng, X.; Zhao, H.; Mao, Y.; Cui, Y. Thymoquinone protects against cerebral small vessel disease: Role of antioxidant and anti-inflammatory activities. J. Boil. Regul. Homeost. Agents 2018, 32, 225–231. [Google Scholar]
- Suddek, G.M. Protective role of thymoquinone against liver damage induced by tamoxifen in female rats. Can. J. Physiol. Pharmacol. 2014, 92, 640–644. [Google Scholar] [CrossRef]
- Nagi, M.N.; Alam, K.; Badary, O.A.; Al-Shabanah, O.A.; Al-Sawaf, H.A.; Al-Bekairi, A.M. Thymoquinone protects against carbon tetrachloride hepatotoxicity in mice via an antioxidant mechanism. Biochem. Mol. Biol. Int. 1999, 47, 153–159. [Google Scholar]
- Umar, S.; Zargan, J.; Umar, K.; Ahmad, S.; Katiyar, C.K.; Khan, H.A. Modulation of the oxidative stress and inflammatory cytokine response by thymoquinone in the collagen induced arthritis in Wistar rats. Chem. Interact. 2012, 197, 40–46. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Parvardeh, S.; Asl, M.N.; Sadeghnia, H.R.; Ziaee, T. Effect of thymoquinone and Nigella sativa seeds oil on lipid peroxidation level during global cerebral ischemia-reperfusion injury in rat hippocampus. Phytomedicine 2007, 14, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; AlRumaihi, F.; Alsahli, M.A.; Alhommrani, M.F.; Khan, A.; Rahmani, A.H. Curcumin, an Active Constituent of Turmeric Spice: Implication in the Prevention of Lung Injury Induced by Benzo(a) Pyrene (BaP) in Rats. Molecules 2020, 25, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, A.; Ali, R.; Ali, N.; Kazim Hasan, S.; Barnwal, P.; Mohammad Afzal, S.; Vafa, A.; Sultana, S. Methanolic bark extract of Acacia catechu ameliorates benzo(a)pyrene induced lung toxicity by abrogation of oxidative stress, inflammation, and apop-tosis in mice. Environ. Toxicol. 2017, 32, 1566–1577. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.; Karin, M. Immunity, Inflammation, and Cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Oberdörster, G. Pulmonary effects of inhaled ultrafine particles. Int. Arch. Occup. Environ. Health 2000, 74, 1–8. [Google Scholar] [CrossRef]
- Multhoff, G.; Molls, M.; Radons, J. Chronic Inflammation in Cancer Development. Front. Immunol. 2012, 2, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Vliet, A.; Eiserich, J.P.; Cross, E.C. Nitric oxide: A pro-inflammatory mediator in lung disease? Respir. Res. 2000, 1, 1–72. [Google Scholar] [CrossRef] [Green Version]
- Qamar, W.; Khan, A.Q.; Khan, R.; Lateef, A.; Tahir, M.; Rehman, M.U.; Ali, F.; Sultana, S. Benzo (a) pyrene-induced pulmonary in-flammation, edema, surfactant dysfunction, and injuries in rats: Alleviation by farnesol. Exp. Lung Res. 2012, 38, 19–27. [Google Scholar] [CrossRef]
- Yetkin, N.A.; Büyükoğlan, H.; Sönmez, M.F.; Tutar, N.; Gülmez, I.; Yilmaz, I. The protective effects of thymoquinone on lung damage caused by cigarette smoke. Biotech. Histochem. 2019, 95, 268–275. [Google Scholar] [CrossRef]
- Suddek, G.M.; Ashry, N.A.; Gameil, N.M. Thymoquinone attenuates cyclophosphamide-induced pulmonary injury in rats. Inflammopharmacology 2013, 21, 427–435. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Almatroudi, A.; Babikr, A.Y.; Khan, A.A.; Alsahli, M.A. Thymoquinone, an Active Constituent of Black Seed Attenuates CCl4 Induced Liver Injury in Mice via Modulation of Antioxidant Enzymes, PTEN, P53 and VEGF Protein. Open Access Maced. J. Med. Sci. 2019, 7, 311–317. [Google Scholar] [CrossRef] [Green Version]
- El Gazzar, M.; El Mezayen, R.; Marecki, J.C.; Nicolls, M.R.; Canastar, A.; Dreskin, S.C. Anti-inflammatory effect of thymoquinone in a mouse model of allergic lung inflammation. Int. Immunopharmacol. 2006, 7, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Nawar, W.W. Lipids. In Food Chemistry; Fennema, O.R., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1996; pp. 225–314. [Google Scholar]
- Shahid, A.; Ali, R.; Ali, N.; Kazim Hasan, S.; Rashid, S.; Majed, F.; Sultana, S. Attenuation of genotoxicity, oxidative stress, apoptosis and inflammation by rutin in benzo(a)pyrene exposed lungs of mice: Plausible role of NF-kappaB, TNF-alpha and Bcl-2. J. Complement. Integr. Med. 2016, 13, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Pourgholamhossein, F.; Sharififar, F.; Rasooli, R.; Pourgholi, L.; Nakhaeipour, F.; Samareh-Fekri, H.; Iranpour, M.; Mandegary, A. Thymoquinone effectively alleviates lung fibrosis induced by paraquat herbicide through down-regulation of pro-fibrotic genes and inhibition of oxidative stress. Environ. Toxicol. Pharmacol. 2016, 45, 340–345. [Google Scholar] [CrossRef] [PubMed]
- El-Khouly, D.; El-Bakly, W.M.; Awad, A.S.; El-Mesallamy, H.O.; El-Demerdash, E. Thymoquinone blocks lung injury and fibrosis by attenuating bleomycin-induced oxidative stress and activation of nuclear factor Kappa-B in rats. Toxicology 2012, 302, 106–113. [Google Scholar] [CrossRef]
- Boskabady, M.; Khazdair, M.R.; Bargi, R.; Saadat, S.; Memarzia, A.; Roshan, N.M.; Hosseini, M.; Askari, V.R.; Boskabady, M.H. Thymoquinone Ameliorates Lung Inflammation and Pathological Changes Observed in Lipopolysaccharide-Induced Lung Injury. Evid. Based Complement. Altern. Med. 2021, 2021, 6681729. [Google Scholar] [CrossRef] [PubMed]
- Islam, J.; Shree, A.; Afzal, S.M.; Vafa, A.; Sultana, S. Protective effect of Diosmin against benzo(a)pyrene-induced lung injury in Swiss Albino Mice. Environ. Toxicol. 2020, 35, 747–757. [Google Scholar] [CrossRef]
- Anandakumar, P.; Kamaraj, S.; Jagan, S.; Ramakrishnan, G.; Asokkumar, S.; Naveenkumar, C.; Raghunandhakumar, S.; Vanitha, M.K.; Devaki, T. The Anticancer Role of Capsaicin in Experimentallyinduced Lung Carcinogenesis. J. Pharmacopunct. 2015, 18, 19–25. [Google Scholar] [CrossRef]
- Al-Gabri, N.; Ali, A.-M.; Al-Attar, E.-S.; Hamed, M. Pathological Study on the Role of Thymoquinone in Experimentally Induced Acute Lung Injury in Rats. Zagazig Veter. J. 2017, 45, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Kanter, M. Thymoquinone attenuates lung injury induced by chronic toluene exposure in rats. Toxicol. Ind. Health 2010, 27, 387–395. [Google Scholar] [CrossRef]
- Zin, W.A.; Silva, A.G.L.S.; Magalhães, C.B.; Carvalho, G.M.C.; Riva, D.R.; Lima, C.C.; Leal-Cardoso, J.H.; Takiya, C.M.; Valença, S.S.; Saldiva, P.H.N.; et al. Eugenol attenuates pulmonary damage induced by diesel exhaust particles. J. Appl. Physiol. 2012, 112, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, B.; Chakraborty, S.; Ghosh, D.; Raha, S.; Sen, P.C.; Jana, K. Benzo(a)pyrene Induced p53 Mediated Male Germ Cell Apop-tosis: Synergistic Protective Effects of Curcumin and Resveratrol. Front. Pharmacol. 2016, 7, 245. [Google Scholar] [CrossRef] [Green Version]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.W. NF-κB in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef]
- Shi, Y.; Dai, J.; Liu, H. Naringenin inhibits allergen-induced airway inflammation and airway responsiveness and inhibits NF-kB activity in a murine model of asthma. Can. J. Physiol. Pharmacol. 2009, 87, 729–735. [Google Scholar] [CrossRef]
- Hassan, S.K.; Mousa, A.M.; El-Sammad, N.M.; Abdel-Halim, A.H.; Khalil, W.K.; Elsayed, E.A.; Anwar, N.; Linscheid, M.W.; Moustafa, E.S.; Hashim, A.N.; et al. Antitumor activity of Cuphea ignea extract against benzo(a)pyrene-induced lung tumorigenesis in Swiss Albino mice. Toxicol. Rep. 2019, 6, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Klopfleisch, R. Multiparametric and semiquantitative scoring systems for the evaluation of mouse model histopathology—A systematic review. BMC Veter. Res. 2013, 9, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for Valid Histopathologic Scoring in Research. Veter. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Hevel, J.M.; Marletta, M.A. Nitric-oxide synthase assays. Methods Enzym. 1994, 233, 250–258. [Google Scholar]
- Husain, N.E.O.S.; Babiker, A.Y.; Albutti, A.S.; Alsahli, M.A.; Aly, S.M.; Rahmani, A.H. Clinicopathological Significance of Vimentin and Cytokeratin Protein in the Genesis of Squamous Cell Carcinoma of Cervix. Obstet. Gynecol. Int. 2016, 2016, 8790120. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, A.; Alzohairy, M.; Khadri, H.; Mandal, A.K.; Rizvi, A.M. Expressional evaluation of Vascular Endothelial Growth Factor (VEGF) protein in urinary bladder carcinoma patients exposed to cigarette smoke. Int. J. Clin. Exp. Pathol. 2012, 5, 195–202. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Number | Short Name | Description |
|---|---|---|
| Group I | C | The rats received normal saline solution |
| Group II | BZ | B(a)P in corn oil (50 mg/kg b.w.) was administered orally [45] thrice a week for 8 consecutive weeks |
| Group III | BZ + TQ | Co-treatment with B(a)P (50 mg/kg b.w.) and TQ (50 mg/kg b.w.), where TQ was given orally before B(a)P administration |
| Group IV | TQ | The rats received TQ (50 mg/kg b.w.) three times/week |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alzohairy, M.A.; Khan, A.A.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. Protective Effects of Thymoquinone, an Active Compound of Nigella sativa, on Rats with Benzo(a)pyrene-Induced Lung Injury through Regulation of Oxidative Stress and Inflammation. Molecules 2021, 26, 3218. https://doi.org/10.3390/molecules26113218
Alzohairy MA, Khan AA, Alsahli MA, Almatroodi SA, Rahmani AH. Protective Effects of Thymoquinone, an Active Compound of Nigella sativa, on Rats with Benzo(a)pyrene-Induced Lung Injury through Regulation of Oxidative Stress and Inflammation. Molecules. 2021; 26(11):3218. https://doi.org/10.3390/molecules26113218
Chicago/Turabian StyleAlzohairy, Mohammad A., Amjad Ali Khan, Mohammed A. Alsahli, Saleh A. Almatroodi, and Arshad Husain Rahmani. 2021. "Protective Effects of Thymoquinone, an Active Compound of Nigella sativa, on Rats with Benzo(a)pyrene-Induced Lung Injury through Regulation of Oxidative Stress and Inflammation" Molecules 26, no. 11: 3218. https://doi.org/10.3390/molecules26113218
APA StyleAlzohairy, M. A., Khan, A. A., Alsahli, M. A., Almatroodi, S. A., & Rahmani, A. H. (2021). Protective Effects of Thymoquinone, an Active Compound of Nigella sativa, on Rats with Benzo(a)pyrene-Induced Lung Injury through Regulation of Oxidative Stress and Inflammation. Molecules, 26(11), 3218. https://doi.org/10.3390/molecules26113218