5-HT1A Serotonergic, α-Adrenergic and Opioidergic Receptors Mediate the Analgesic Efficacy of Vortioxetine in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Motor Coordination
Rota-Rod Test
2.2. Analgesic Activity
2.2.1. Analgesic Effect of Vortioxetine on Mechanical Noxious Stimuli-Induced Pain Behavior
2.2.2. Spinally Mediated Analgesic Effect of Vortioxetine on Thermal Noxious Stimuli-Induced Pain Behavior
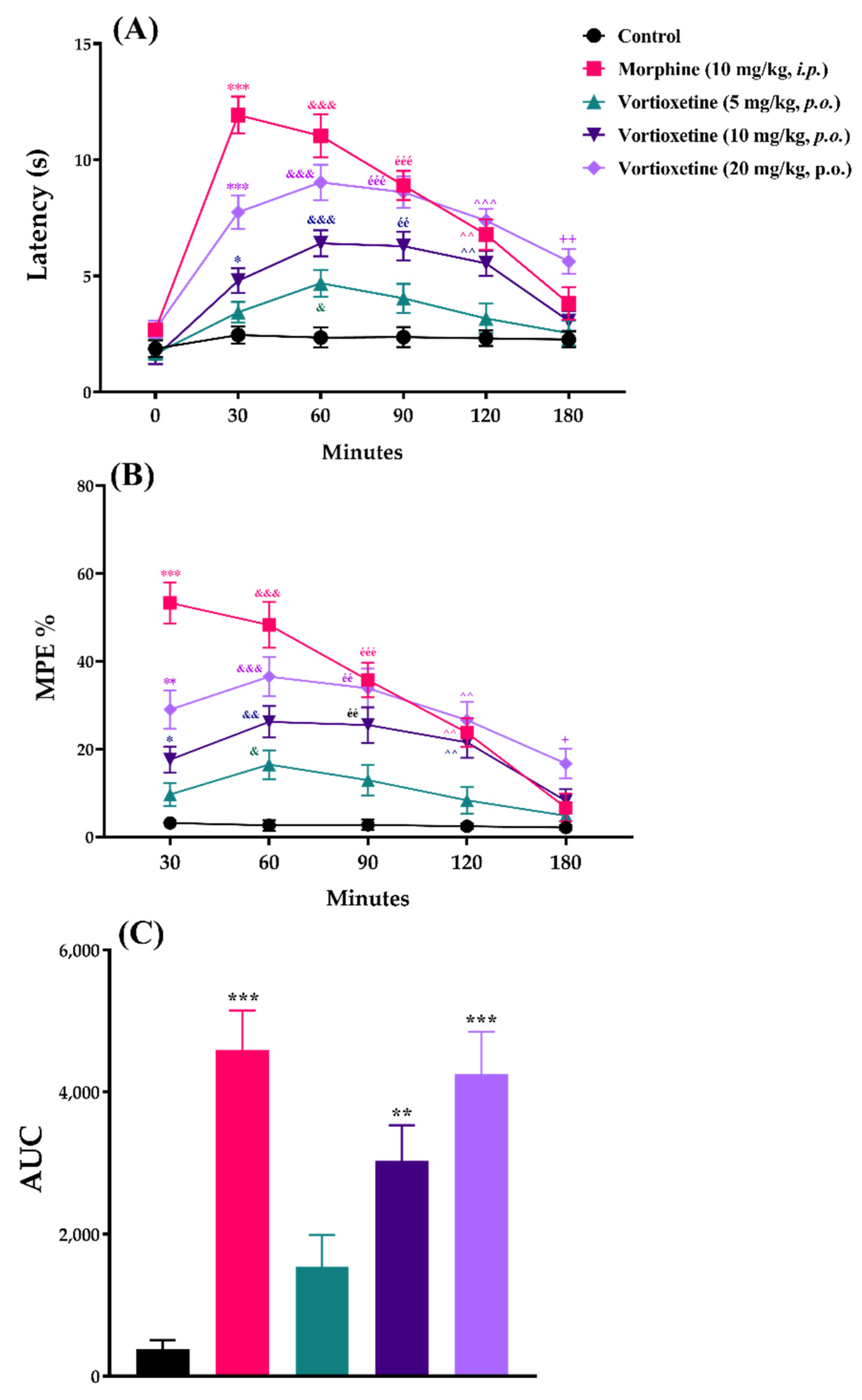
2.2.3. Supraspinally Mediated Analgesic Effect of Vortioxetine on Thermal Noxious Stimuli-Induced Pain Behavior
2.3. Mechanistic Studies
2.3.1. Involvement of Serotonergic System in the Analgesic Effect of Vortioxetine
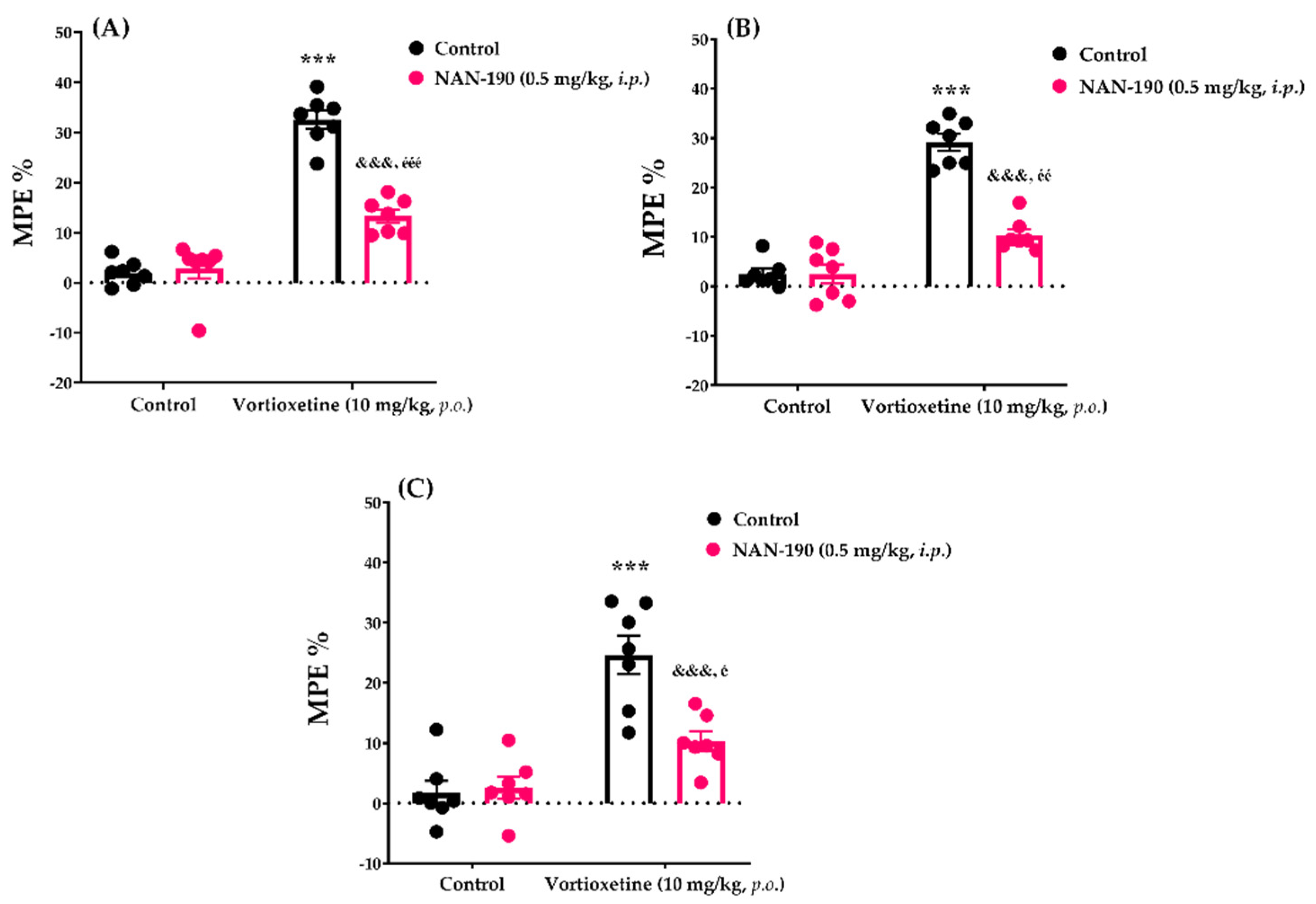
2.3.2. Involvement of Serotonergic 5HT1A Receptors in the Analgesic Effect of Vortioxetine
2.3.3. Involvement of Catecholaminergic System in the Analgesic Effect of Vortioxetine
2.3.4. Involvement of α-Adrenergic Receptors in the Analgesic Effect of Vortioxetine
2.3.5. Involvement of β-Adrenergic Receptors in the Analgesic Effect of Vortioxetine
2.3.6. Involvement of Opioid Receptors in the Analgesic Effect of Vortioxetine
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Administration Protocol
4.3. Motor Coordination Analysis
Rota-Rod Test
4.4. Analgesic Activity Analysis
4.4.1. Tail-Clip Test
4.4.2. Tail-Immersion Test
4.4.3. Hot-Plate Test
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garnock-Jones, K.P. Vortioxetine: A review of its use in major depressive disorder. CNS Drugs 2014, 28, 855–874. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Asin, K.E.; Artigas, F. Vortioxetine, a novel antidepressant with multimodal activity: Review of preclinical and clinical data. Pharm. Ther. 2015, 145, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Mørk, A.; Pehrson, A.; Brennum, L.T.; Nielsen, S.M.; Zhong, H.; Lassen, A.B.; Miller, S.; Westrich, L.; Boyle, N.J.; Sánchez, C.; et al. Pharmacological effects of Lu AA21004: A novel multimodal compound for the treatment of major depressive disorder. J. Pharm. Exp. Ther. 2012, 340, 666–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pehrson, A.L.; Cremers, T.; Bétry, C.; van der Hart, M.G.; Jørgensen, L.; Madsen, M.; Haddjeri, N.; Ebert, B.; Sanchez, C. Lu AA21004, a novel multimodal antidepressant, produces regionally selective increases of multiple neurotransmitters-a rat microdialysis and electrophysiology study. Eur. Neuropsychopharmacol. 2013, 23, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Moore, N.; Bang-Andersen, B.; Brennum, L.; Frederiksen, K.; Hogg, S.; Mork, A.; Stensbol, T.; Zhong, H.; Sanchez, C.; Smith, D. Lu AA21004: A novel potential treatment for mood disorders. Eur. Neuropsychopharmacol. 2008, 18, 321. [Google Scholar] [CrossRef]
- Mørk, A.; Montezinho, L.P.; Miller, S.; Trippodi-Murphy, C.; Plath, N.; Li, Y.; Gulinello, M.; Sanchez, C. Vortioxetine (Lu AA21004), a novel multimodal antidepressant, enhances memory in rats. Pharmacol. Biochem. Behav. 2013, 105, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Pehrson, A.; Li, Y.; Haddjeri, N.; Gulinello, M.; Sanchez, C. Vortioxetine, a novel multimodal antidepressant, modulates GABA and glutamate neurotransmission via serotonergic mechanisms. Eur. Neuropsychopharmacol. 2013, 23, 196–197. [Google Scholar] [CrossRef]
- Ozbeyli, D.; Aykac, A.; Alaca, N.; Hazar-Yavuz, A.N.; Ozkan, N.; Sener, G. Protective effects of vortioxetine in predator scent stress model of post-traumatic stress disorder in rats: Role on neuroplasticity and apoptosis. J. Physiol. Pharm. 2019, 70, 557–571. [Google Scholar]
- Shah, A.; Northcutt, J. An open-label, flexible dose adaptive study evaluating the efficacy of vortioxetine in subjects with panic disorder. Ann. Gen. Psychiatry 2018, 17, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ögün, M.N.; Çetinkaya, A.; Beyazçiçek, E. The effect of vortioxetine on penicillin-induced epileptiform activity in rats. Arq. Neuropsiquiatr. 2019, 77, 412–417. [Google Scholar] [CrossRef]
- Berrocoso, E.; Mico, J.A. Role of serotonin 5-HT1A receptors in the antidepressant-like effect and the antinociceptive effect of venlafaxine in mice. Int. J. Neuropsychopharmacol. 2009, 12, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Bomholt, S.F.; Mikkelsen, J.D.; Blackburn-Munro, G. Antinociceptive effects of the antidepressants amitriptyline, duloxetine, mirtazapine and citalopram in animal models of acute, persistent and neuropathic pain. Neuropharmacology 2005, 48, 252–263. [Google Scholar] [CrossRef]
- Cartmell, S.M.; Gelgor, L.; Mitchell, D. A revised rotarod procedure for measuring the effect of antinociceptive drugs on motor function in the rat. J. Pharmacol. Methods 1991, 6, 149–159. [Google Scholar] [CrossRef]
- Kasap, M.; Can, Ö.D. Opioid system mediated anti-nociceptive effect of agomelatine in mice. Life Sci. 2016, 15, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Guilloux, J.P.; Mendez-David, I.; Pehrson, A.; Guiard, B.P.; Repérant, C.; Orvoën, S.; Gardier, A.M.; Hen, R.; Ebert, B.; Miller, S.; et al. Antidepressant and anxiolytic potential of the multimodal antidepressant vortioxetine (Lu AA21004) assessed by behavioural and neurogenesis outcomes in mice. Neuropharmacology 2013, 73, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuena, A.R.; Maftei, D.; Alemà, G.S.; Dal Moro, F.; Lattanzi, R.; Casolini, P.; Nicoletti, F. Multimodal antidepressant vortioxetine causes analgesia in a mouse model of chronic neuropathic pain. Mol. Pain 2018, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bars, D.; Gozariu, M.; Cadden, S.W. Animal models of nociception. Pharmacol. Rev. 2001, 53, 597–652. [Google Scholar] [PubMed]
- Shajib, S.; Rashid, R.B.; Ming, L.C.; Islam, S.; Sarker, M.R.; Nahar, L.; Sarker, S.D.; Datta, B.K.; Rashid, M.A. Polymethoxyflavones from Nicotiana plumbaginifolia (Solanaceae) exert antinociceptive and neuropharmacological effects in mice. Front. Pharmacol. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Micov, A.M.; Tomić, M.A.; Todorović, M.B.; Vuković, M.J.; Pecikoza, U.B.; Jasnic, N.I.; Djordjevic, J.D.; Stepanović-Petrović, R.M. Vortioxetine reduces pain hypersensitivity and associated depression-like behavior in mice with oxaliplatin-induced neuropathy. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 103, 109975. [Google Scholar] [CrossRef] [PubMed]
- Bang-Andersen, B.; Faldt, A.; Moerk, A.; Lopez De Diego, H.; Holm, R.; Stensboel, T.B.; Ringgaard, L.M.; Mealy, M.J.; Rock, M.H.; Brodersen, J.; et al. 1-[2-(2,4-dimethylphenylsulfanyl)-phenyl] Piperazine as a Compound with Combined Serotonin Reuptake, 5-HT3 and 5-HT1A Activity for the Treatment of Cognitive Impairment. WO 2007/144005 A1, 21 December 2007. [Google Scholar]
- Moore, N.; Stensboel, T.B. 1-[2-(2,4-dimethylphenylsulfanyl)-phenyl] Piperazine as a Compound with Combined Serotonin Reuptake, 5-HT3 and 5-HT1A Activity for the Treatment of Pain or Residual Symptoms in Depression Relating to Sleep and Cognition. WO 2008/113359 A2, 25 September 2008. [Google Scholar]
- Holden, J.E.; Jeong, Y.; Forrest, J.M. The endogenous opioid system and clinical pain management. AACN Clin. Issues 2005, 16, 291–301. [Google Scholar] [CrossRef]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [CrossRef]
- Yaksh, T.L. Pharmacology of spinal adrenergic systems which modulate spinal nociceptive processing. Pharmacol. Biochem. Behav. 1985, 22, 845–858. [Google Scholar] [CrossRef]
- Koe, B.K.; Weissman, A. p-Chlorophenylalanine: A specific depletor of brain serotonin. J. Pharmacol. Exp. Ther. 1996, 154, 499–516. [Google Scholar]
- Sommer, C. Serotonin in pain and pain control. In Handbook of Behavioral Neurobiology of Serotonin, 1st ed.; Müller, C., Jacobs, B.L., Eds.; Academic Press: Cambridge, MA, USA, 2010; Volume 21, p. 462. [Google Scholar]
- Cortes-Altamirano, J.L.; Olmos-Hernandez, A.; Jaime, H.B.; Carrillo-Mora, P.; Bandala, C.; Reyes-Long, S.; Alfaro-Rodríguez, A. Review: 5-HT1, 5-HT2, 5-HT3 and 5-HT7 receptors and their role in the modulation of pain response in the central nervous system. Curr. Neuropharmacol. 2018, 16, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Marlier, L.; Teilhac, J.R.; Cerruti, C.; Privat, A. Autoradiographic mapping of 5-HT1, 5-HT1A, 5-HT1B and 5-HT2 receptors in the rat spinal cord. Brain Res. 1991, 550, 15–23. [Google Scholar] [CrossRef]
- Daval, G.; Vergé, D.; Basbaum, A.I.; Bourgoin, S.; Hamon, M. Autoradiographic evidence of serotonin 1 binding sites on primary afferent fibers in the dorsal horn of the rat spinal cord. Neurosci. Lett. 1987, 83, 71–76. [Google Scholar] [CrossRef]
- Jeong, H.J.; Mitchell, V.A.; Vaughan, C.W. Role of 5-HT1 receptor subtypes in the modulation of pain and synaptic transmission in rat spinal superficial dorsal horn. Br. J. Pharm. 2012, 165, 1956–1965. [Google Scholar] [CrossRef] [Green Version]
- Nadeson, R.; Goodchild, C.S. Antinociceptive role of 5-HT1A receptors in rat spinal cord. Br. J. Anaesth. 2002, 88, 679–684. [Google Scholar] [CrossRef] [Green Version]
- Gjerstad, J.; Tjѳlsen, A.; Hole, K. The effect of 5-HTIA receptor stimulation on nociceptive dorsal horn neurones in rats. Eur. J. Pharm. 1996, 318, 315–321. [Google Scholar] [CrossRef]
- Eide, P.K.; Joly, N.M.; Hole, K. The role of spinal cord 5-HT1A and 5-HT1B receptors in the modulation of a spinal nociceptive reflex. Brain Res. 1990, 536, 195–200. [Google Scholar] [CrossRef]
- Newman-Tancredi, A.; Bardin, L.; Auclair, A.; Colpaert, F.; Depoortère, R.; Varney, M.A. NLX-112, a highly selective 5-HT 1A receptor agonist, mediates analgesia and antidepressant-like activity in rats via spinal cord and prefrontal cortex 5-HT 1A receptors, respectively. Brain Res. 2018, 1688, 1–7. [Google Scholar] [CrossRef]
- Sałat, K.; Kołaczkowski, M.; Furgała, A.; Rojek, A.; Sniecikowska, J.; Varney, M.A.; Newman-Tancredi, A. Antinociceptive, antiallodynic and antihyperalgesic effects of the 5-HT1A receptor selective agonist, NLX-112 in mouse models of pain. Neuropharmacology 2017, 125, 181–188. [Google Scholar] [CrossRef]
- Valhondo, M.; Marco, I.; Martín-Fontecha, M.; Vázquez-Villa, H.; Ramos, J.A.; Berkels, R.; Lauterbach, T.; Benhamú, B.; López-Rodríguez, M.L. New serotonin 5-HT1A receptor agonists endowed with antinociceptive activity in vivo. J. Med. Chem. 2013, 56, 7851–7861. [Google Scholar] [CrossRef]
- Bardin, L.; Tarayre, J.P.; Malfetes, N.; Koek, W.; Colpaert, F.C. Profound, non-opioid analgesia produced by the high-efficacy 5-HT(1A) agonist F 13640 in the formalin model of tonic nociceptive pain. Pharmacology 2003, 67, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Giordano, J.; Rogersa, L.V. Putative mechanisms of buspirone-induced antinociception in the rat. Pain 1992, 50, 365–372. [Google Scholar] [CrossRef]
- Widerlöv, E.; Lewander, T. Inhibition of the in vivo biosynthesis and changes of catecholamine levels in rat brain after alpha-methyl-p-tyrosine; time- and dose–response relationships. Naunyn Schmiedebergs Arch. Pharm. 1978, 304, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Mayorga, A.J.; Dalvi, A.; Page, M.E.; Zimov-Levinson, S.; Hen, R.; Lucki, I. Antidepressant-like behavioral effects in 5-hydroxytryptamine (1A) and 5-hydroxytryptamine (1B) receptor mutant mice. J. Pharm. Exp. Ther. 2001, 298, 1101–1107. [Google Scholar]
- Baba, H.; Shirnoji, K.; Yoshirnura, M. Norepinephrine facilitates inhibitory transmission in substantia gehtinosa of adult rat spinal cord (Part 1): Effects on axon terminals of GABAergic and glycinergic neurons. Anesthesiology 2000, 92, 473–484. [Google Scholar] [CrossRef]
- Fukui, M.; Nakagawa, T.; Minami, M.; Satoh, M. Involvement of β2-adrenergic and µ-opioid receptors in antinociception produced by intracerebroventricular administration of α, β-methylene-ATP. Jpn. J. Pharm. 2001, 86, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Pertovaara, A. Noradrenergic pain modulation. Prog. Neurobiol. 2006, 80, 53–83. [Google Scholar] [CrossRef]
- Sonohata, M.; Furue, H.; Katafuchi, T.; Yasaka, T.; Doi, A.; Kumamoto, E.; Yoshimura, M. Actions of noradrenaline on substantia gelatinosa neurones in the rat spinal cord revealed by in vivo patch recording. J. Physiol. 2004, 555, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Sudo, R.T.; do Amaral, R.V.; da Silva Monteiro, C.E.D.S.; Pitta, I.D.R.; Lima, M.D.C.; Montes, G.C.; Ririe, D.G.; Hayashida, K.; Zapata-Sudo, G. Antinociception induced by a novel α2A adrenergic receptor agonist in rodents acute and chronic pain models. Eur. J. Pharm. 2017, 815, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Chabot-Doré, A.J.; Schuster, D.J.; Stone, L.S.; Wilcox, G.L. Analgesic synergy between opioid and α2-adrenoceptors. Br. J. Pharm. 2015, 172, 388–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fields, H. State-dependent opioid control of pain. Nat. Rev. Neurosci. 2004, 5, 565–575. [Google Scholar] [CrossRef]
- Zhao, Z.Q.; Gao, Y.J.; Sun, Y.G.; Zhao, C.S.; Gereau, R.W., 4th; Chen, Z.F. Central serotonergic neurons are differentially required for opioid analgesia but not for morphine tolerance or morphine reward. Proc. Natl. Acad. Sci. USA 2007, 104, 14519–14524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, B.B. Catecholamines, symapthomimetic drugs, and adrenergic receptor antagonists. In Goodman & Gilman’s The Pharmacological Basis of Therapeutics, 10th ed.; Hardman, J.G., Limbird, L.E., Gilman, A.G., Eds.; McGraw-Hill: New York, NY, USA, 2001; p. 245. [Google Scholar]
- Sear, J.W. Antihypertensive drugs and vasodilators. In Pharmacology and Physiology for Anesthesia Foundations and Clinical Application, 2nd ed.; Hemmings, H.C., Egan, T.D., Eds.; Saunders: Philadelphia, PA, USA, 2019; pp. 535–555. [Google Scholar]
- Gruetter, C.A. Phentolamine. In xPharm: The Comprehensive Pharmacology Reference, 1st ed.; Enna, S.J., Bylund, D.B., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 1–5. [Google Scholar]
- Vardanyan, R.; Hruby, V. Drugs for treatment of erectile dysfunction. In Synthesis of Best-Seller Drugs, 1st ed.; Vardanyan, R., Hruby, V., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 783–800. [Google Scholar]
- Foong, J.P.; Bornstein, J.C. 5-HT antagonists NAN-190 and SB 269970 block alpha2-adrenoceptors in the guinea pig. Neuroreport 2009, 20, 325–330. [Google Scholar] [CrossRef]
- Tinajero, J.C.; Fabbri, A.; Dufau, M.L. Serotonergic inhibition of rat Leydig cell function by propranolol. Endocrinology 1993, 133, 257–264. [Google Scholar] [CrossRef]
- Demir Özkay, U.; Can, O.D. Anti-nociceptive effect of vitexin mediated by the opioid system in mice. Pharmacol. Biochem. Behav. 2013, 109, 23–30. [Google Scholar] [CrossRef]
- De Freitas Pires, A.; Assreuy, A.M.; Lopes, É.A.; Celedônio, N.R.; Soares, C.E.; Rodrigues, N.V.; Sousa, P.L.; Benevides, R.G.; Nagano, C.S.; Cavada, B.S.; et al. Opioid-like antinociceptive effects of oral administration of a lectin purified from the seeds of Canavalia brasiliensis. Fundam. Clin. Pharm. 2013, 27, 201–209. [Google Scholar] [CrossRef]
- Hess, S.; Padoani, C.; Scorteganha, L.C.; Holzmann, I.; Malheiros, A.; Yunes, R.A.; Delle Monache, F.; de Souza, M.M. Assessment of mechanisms involved in antinociception caused by myrsinoic acid B. Biol. Pharm. Bull. 2010, 33, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Fajemiroye, J.O.; Galdino, P.M.; Alves, S.F.; de Paula, J.A.; de Paula, J.R.; Ghedini, P.C.; Costa, E.A. Involvement of 5-HT1A in the anxiolytic-like effect of dichloromethane fraction of Pimenta pseudocaryophyllus. J. Ethnopharmacol. 2012, 141, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Can, Ö.D.; Turan, N.; Demir Özkay, Ü.; Öztürk, Y. Antidepressant-like effect of gallic acid in mice: Dual involvement of serotonergic and catecholaminergic systems. Life Sci. 2017, 190, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.; Frishtick, R.; Volis, I.; Rubovitch, V.; Pick, C.G.; Weizman, R. The antinociceptive properties of reboxetine in acute pain. Eur. Neuropsychopharmacol. 2009, 19, 735–739. [Google Scholar] [CrossRef]
- Schreiber, S.; Backer, M.M.; Herman, I.; Shamir, D.; Boniel, T.; Pick, C.G. The antinociceptive effect of trazodone in mice is mediated through both mu-opioid and serotonergic mechanisms. Behav. Brain Res. 2000, 114, 51–56. [Google Scholar] [CrossRef]
- Schreiber, S.; Backer, M.M.; Pick, C.G. The antinociceptive effect of venlafaxine in mice is mediated through opioid and adrenergic mechanisms. Neurosci. Lett. 1999, 273, 85–88. [Google Scholar] [CrossRef]
- Boschi, G.; Desiles, M.; Reny, V.; Rips, R.; Wrigglesworth, S. Antinociceptive properties of thyrotropin releasing hormone in mice: Comparison with morphine. Br. J. Pharm. 1983, 79, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Salam, O.M.; El-Batran, S. Pharmacological investigation of trimetazidine in models of inflammation, pain and gastric injury in rodents. Pharmacology 2005, 75, 122–132. [Google Scholar] [CrossRef]
- de Oliveira, D.R.; da Silva, D.M.; Florentino, I.F.; de Brito, A.F.; Fajemiroye, J.O.; da Silva, D.P.B.; da Rocha, F.F.; Costa, E.A.; Galdino, P.M. Monoamine involvement in the antidepressant-like effect of β-caryophyllene. CNS Neurol. Disord. Drug Targets 2018, 17, 309–320. [Google Scholar] [CrossRef]
- Chen, H.S.; Gu, L.J.; Yang, Y.X.; Guo, J.Y. GABA and 5-HT systems are involved in the anxiolytic effect of Gan-Mai-Da-Zao decoction. Front. Neurosci. 2019, 12, 1043. [Google Scholar] [CrossRef]
- Santos, A.R.; Gadotti, V.M.; Oliveira, G.L.; Tibola, D.; Paszcuk, A.F.; Neto, A.; Spindola, H.M.; Souza, M.M.; Rodrigues, A.L.; Calixto, J.B. Mechanisms involved in the antinociception caused by agmatine in mice. Neuropharmacology 2005, 248, 1021–1034. [Google Scholar] [CrossRef]
- Ghelardini, C.; Galeotti, N.; Bartolini, A. Caffeine induces central cholinergic analgesia. Naunyn Schmiedebergs Arch. Pharm. 1997, 356, 590–595. [Google Scholar] [CrossRef] [Green Version]
- Dunham, N.W.; Miya, T.S. A note on a simple apparatus for detecting neurological deficit in rats and mice. J. Am. Pharm. Assoc. 1957, 46, 208–209. [Google Scholar] [CrossRef] [PubMed]
- Woolfe, G.; MacDonald, A.D. The evaluation of analgesic action of pethidine hydrochloride (Demerol). J. Pharm. Exp. Ther. 1944, 80, 300–307. [Google Scholar]
- Argoff, C. Mechanisms of pain transmission and pharmacologic management. Curr. Med. Res. Opin. 2011, 27, 2019–2031. [Google Scholar] [CrossRef] [PubMed]
- Gilron, I.; Bailey, J.M.; Tu, D.; Holden, R.R.; Weaver, D.F.; Houlden, R.L. Morphine, gabapentin, or their combination for neuropathic pain. N. Engl. J. Med. 2005, 352, 1324–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, D.J.; Tritschler, L.; Guilloux, J.P.; Gardier, A.M.; Sanchez, C.; Gaillard, R. Pharmacological properties of vortioxetine and its pre-clinical consequences. Encephale 2016, 42, 1S12–1S23. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turan Yücel, N.; Kandemir, Ü.; Demir Özkay, Ü.; Can, Ö.D. 5-HT1A Serotonergic, α-Adrenergic and Opioidergic Receptors Mediate the Analgesic Efficacy of Vortioxetine in Mice. Molecules 2021, 26, 3242. https://doi.org/10.3390/molecules26113242
Turan Yücel N, Kandemir Ü, Demir Özkay Ü, Can ÖD. 5-HT1A Serotonergic, α-Adrenergic and Opioidergic Receptors Mediate the Analgesic Efficacy of Vortioxetine in Mice. Molecules. 2021; 26(11):3242. https://doi.org/10.3390/molecules26113242
Chicago/Turabian StyleTuran Yücel, Nazlı, Ümmühan Kandemir, Ümide Demir Özkay, and Özgür Devrim Can. 2021. "5-HT1A Serotonergic, α-Adrenergic and Opioidergic Receptors Mediate the Analgesic Efficacy of Vortioxetine in Mice" Molecules 26, no. 11: 3242. https://doi.org/10.3390/molecules26113242
APA StyleTuran Yücel, N., Kandemir, Ü., Demir Özkay, Ü., & Can, Ö. D. (2021). 5-HT1A Serotonergic, α-Adrenergic and Opioidergic Receptors Mediate the Analgesic Efficacy of Vortioxetine in Mice. Molecules, 26(11), 3242. https://doi.org/10.3390/molecules26113242