
An Exploration of Seaweed Polysaccharides Stimulating Denitrifying Bacteria for Safer Nitrate Removal



Abstract
:1. Introduction
2. Results
2.1. Chemical Analysis of the Three Seaweed Polysaccharides
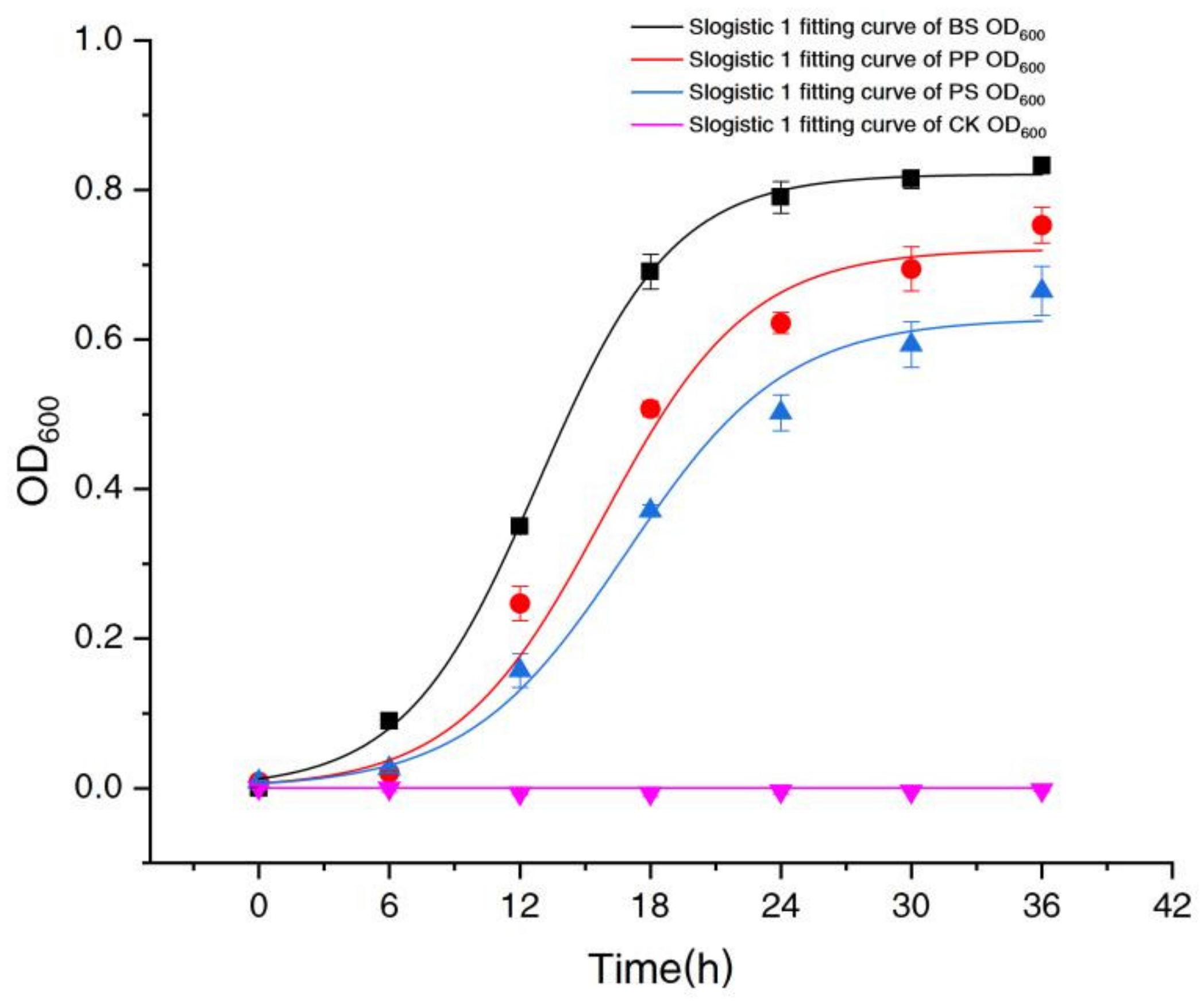
2.2. Comparison of the Growth and Denitrification Performance of Three Denitrifying Bacteria
2.2.1. Comparison of the Growth of Three Denitrifying Bacteria
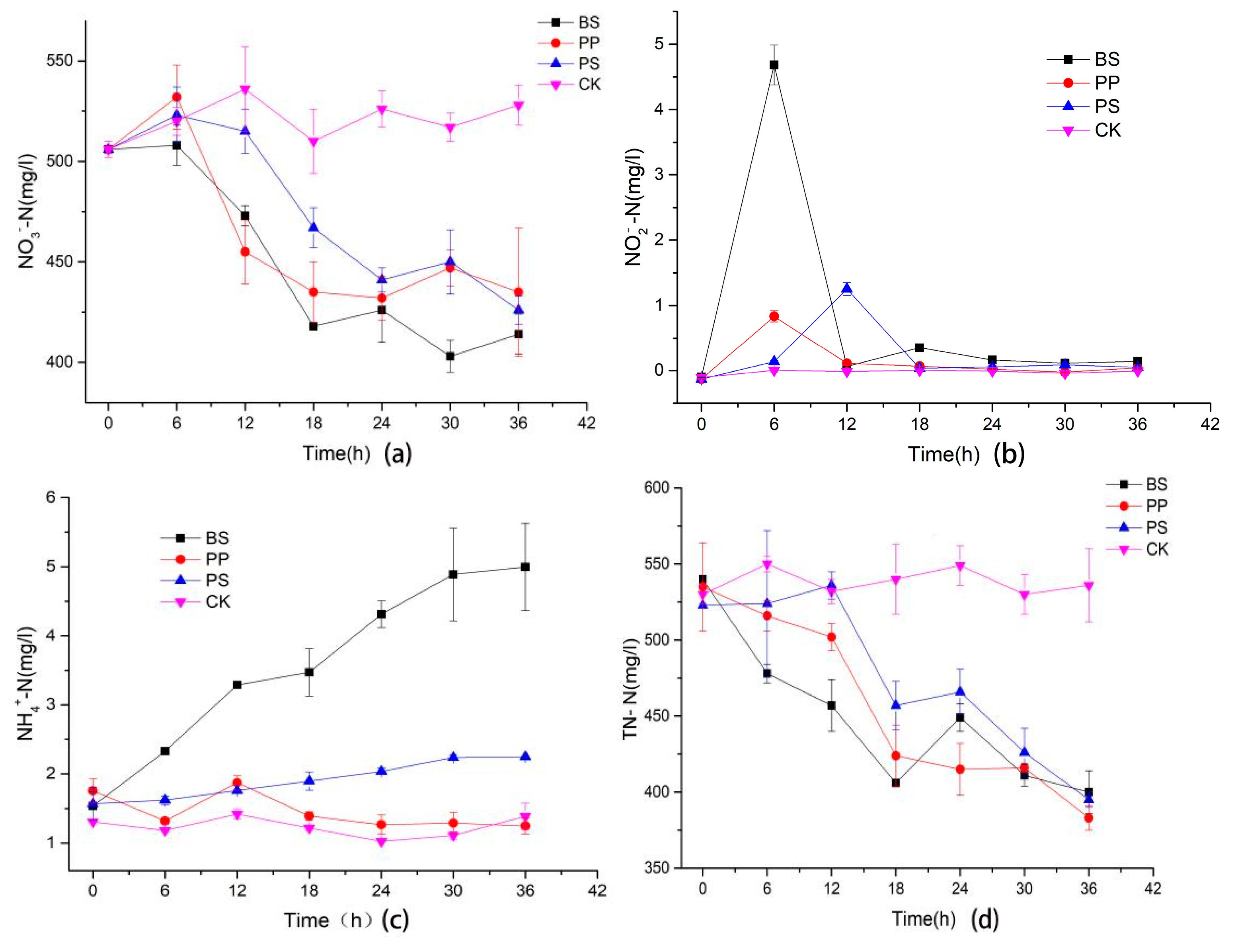
2.2.2. The Changes of Inorganic Nitrogen Concentration in Fermentation Liquid of Bacillus Subtilis (BS), Pseudomonas Stutzeri (PS) and Pseudomonas Putida (PP)
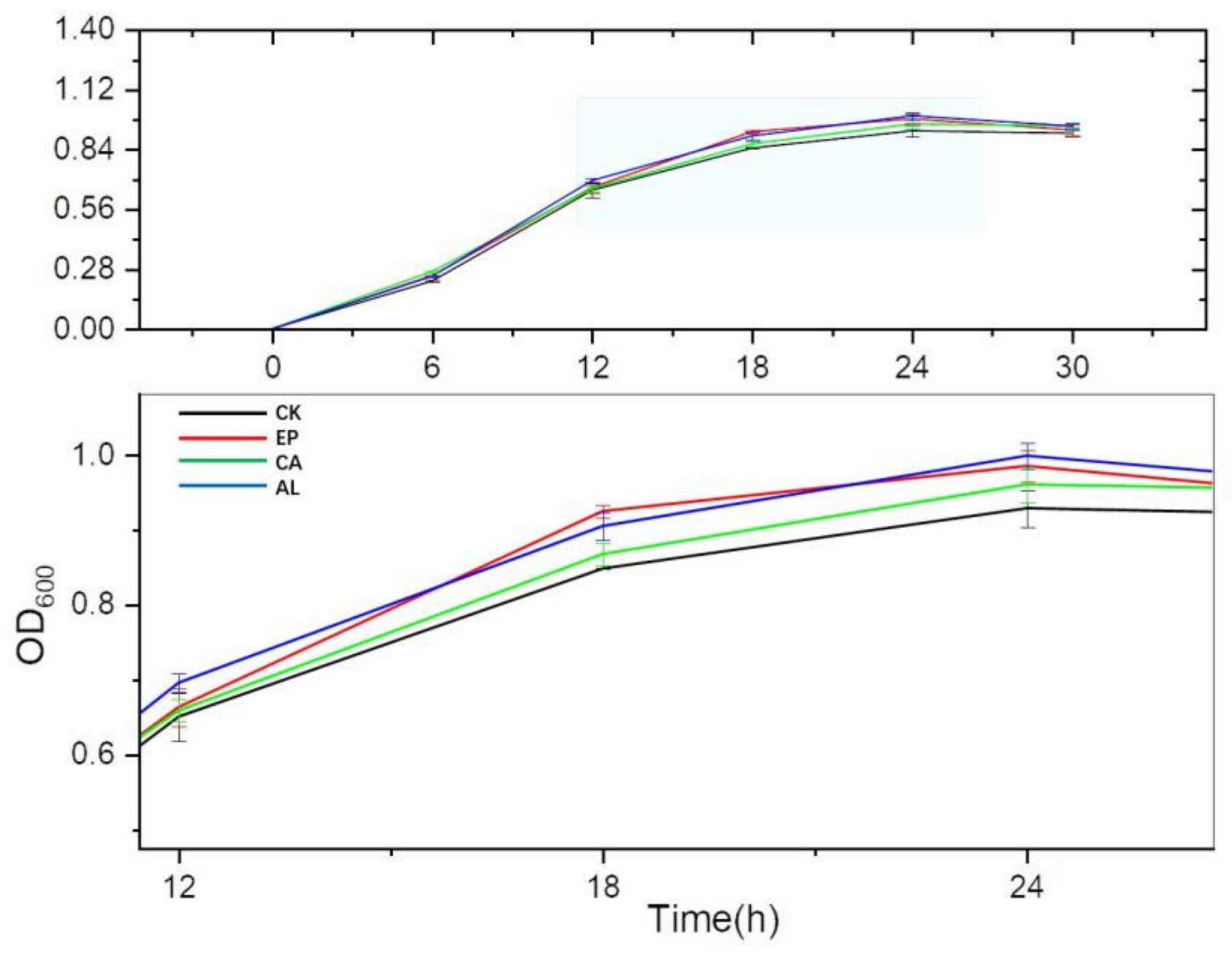
2.3. Effects of Seaweed Polysaccharides on the Growth and Denitrification of Bacillus subtilis (BS)
2.3.1. Effects of Seaweed Polysaccharides on the Growth of Bacillus subtilis (BS)
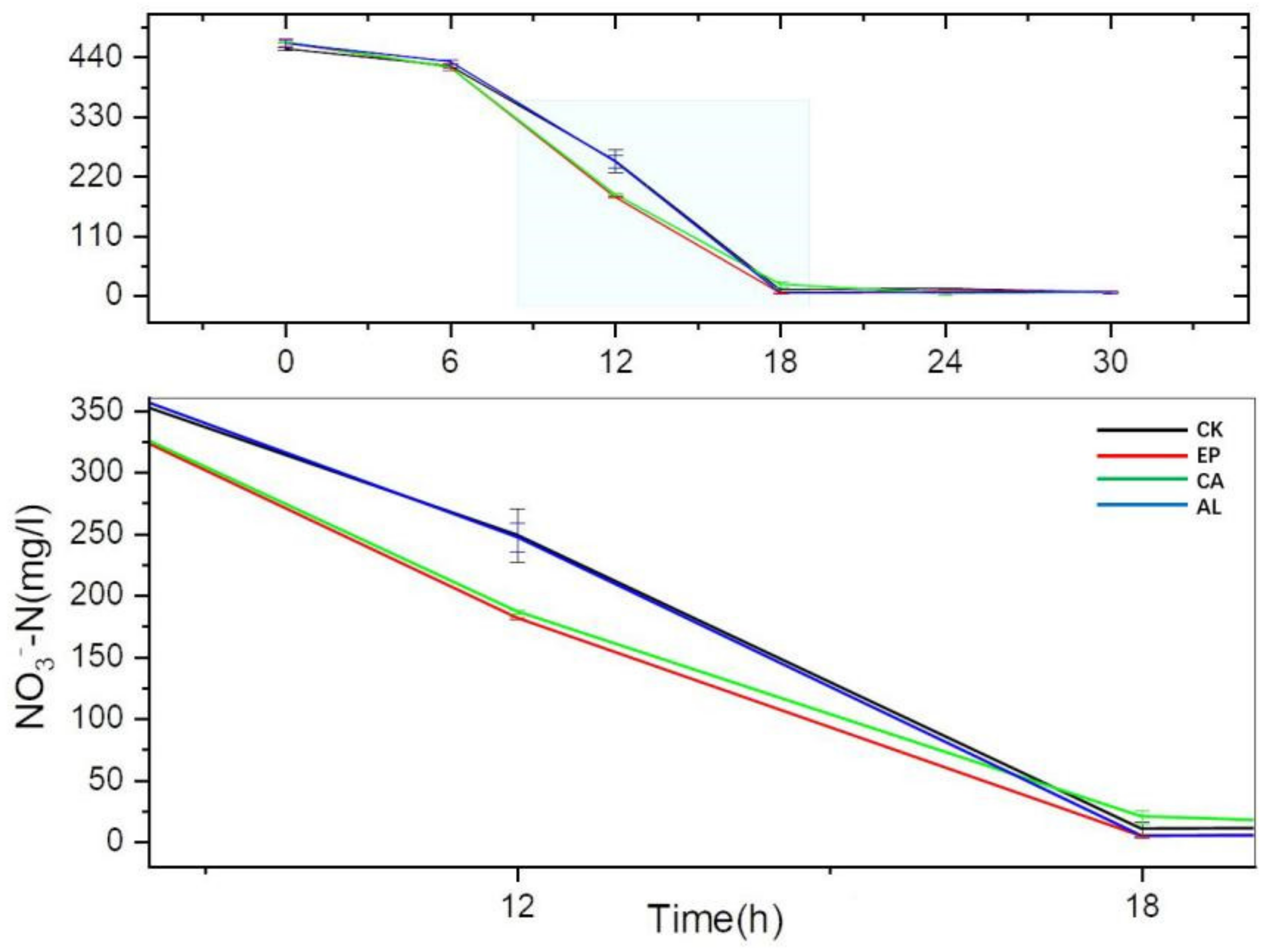
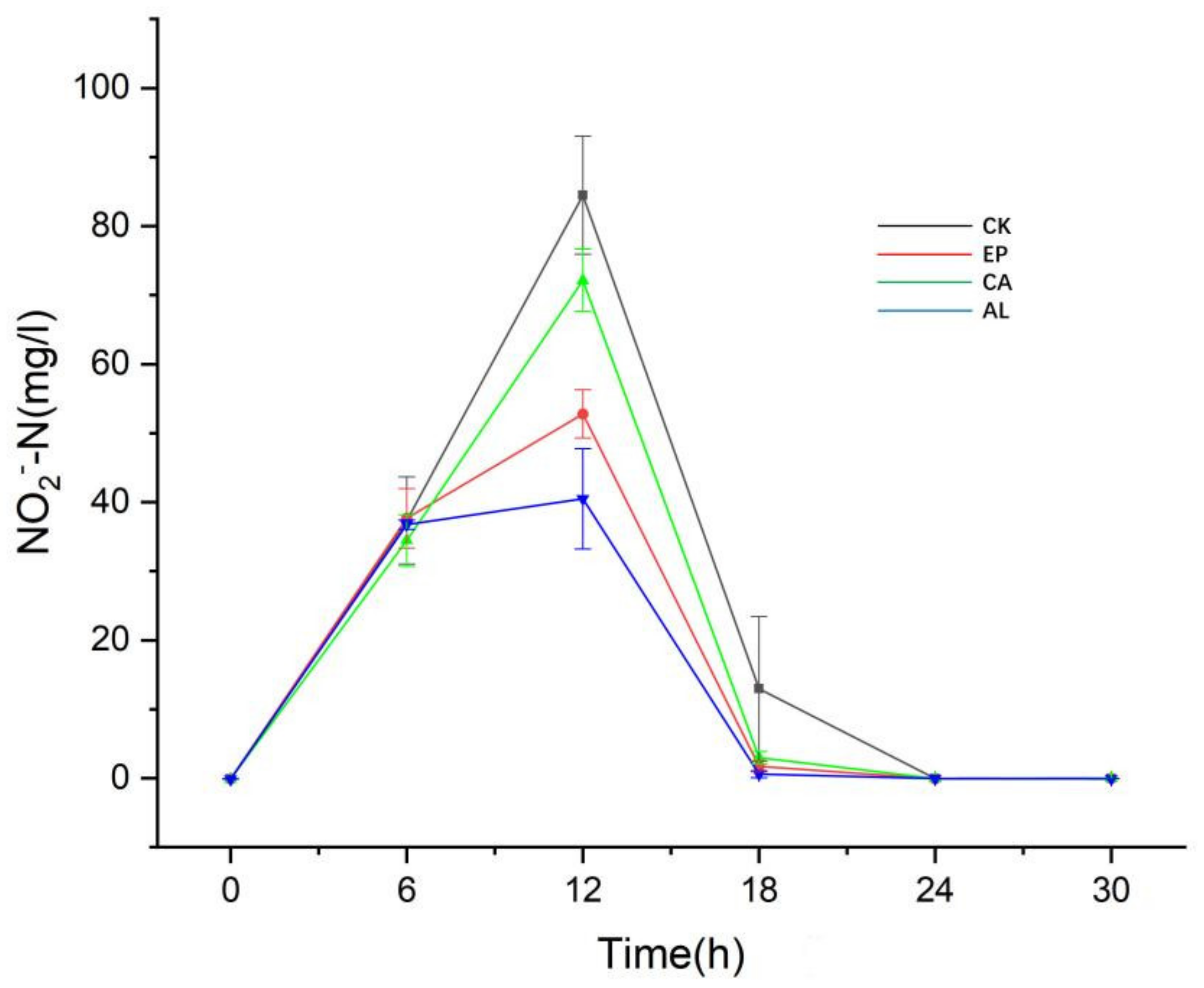
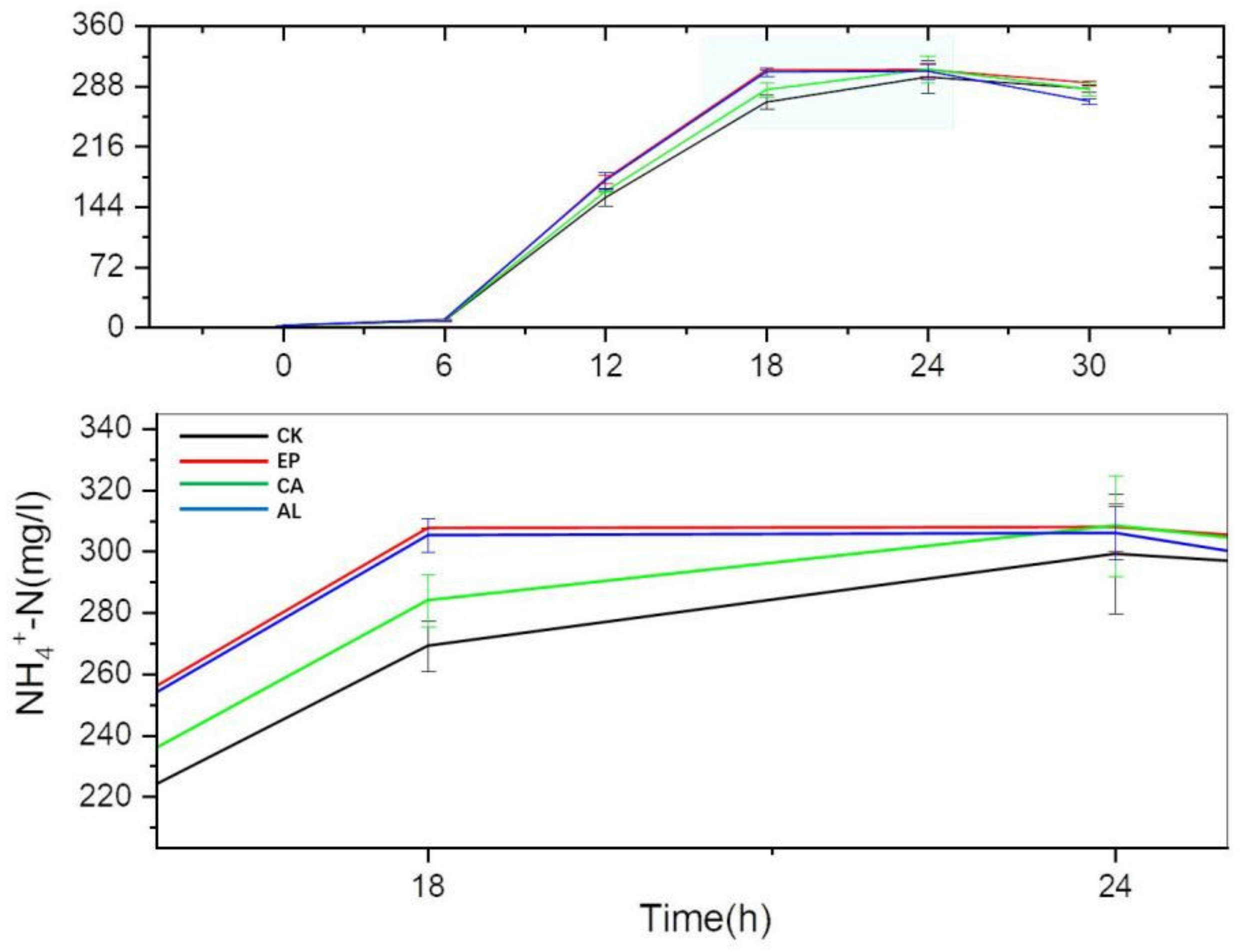
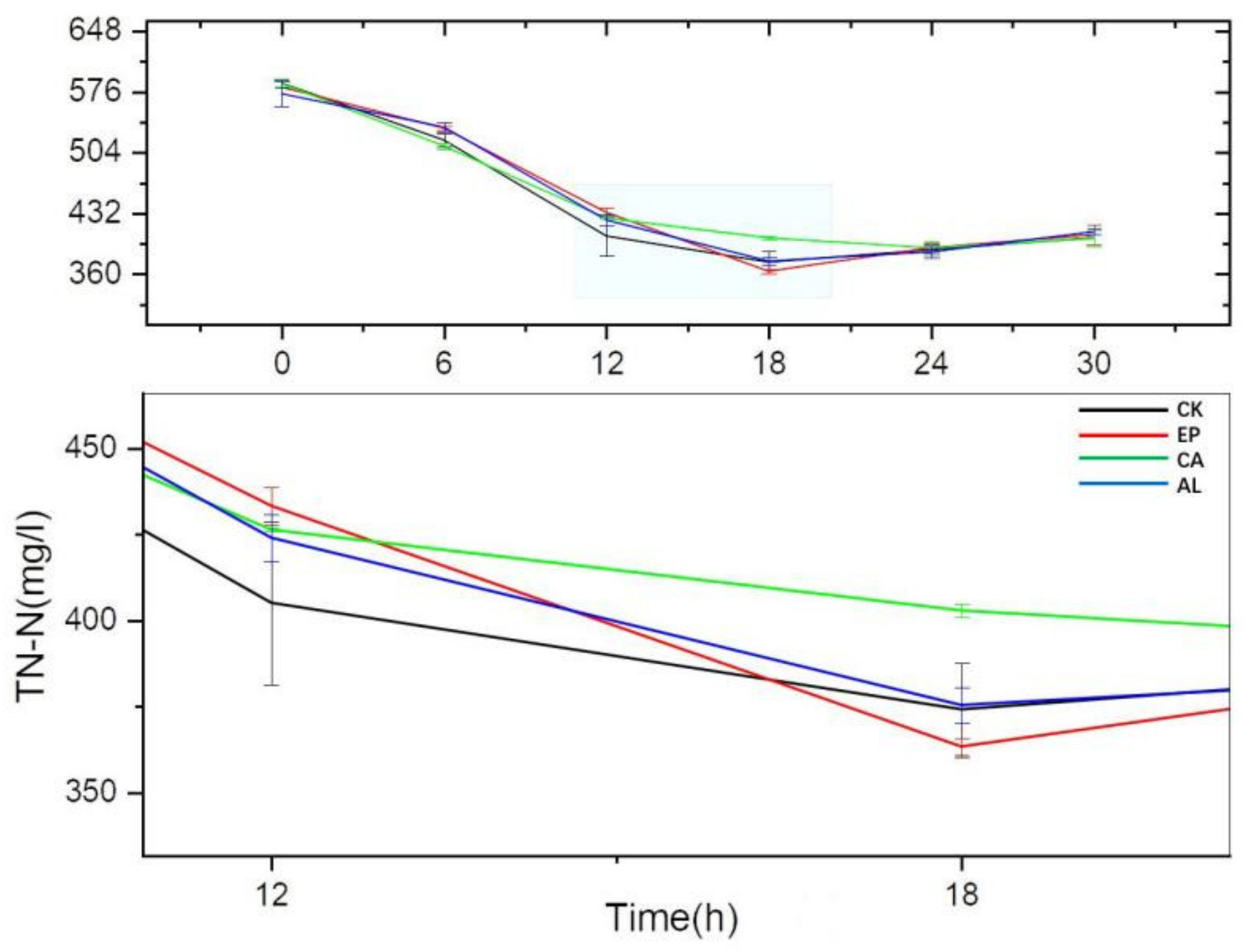
2.3.2. The Changes of Inorganic Nitrogen Concentration in Fermentation Liquid of Enteromorpha linza Polysaccharides (EP), Carrageenan (CA), Sodium Alginate (AL) and Control (CK) Groups
3. Discussion
4. Materials and Methods
4.1. Materials and Bacteria
4.2. Preparation of Seaweed Polysaccharides
4.3. Chemical Analysis of Seaweed Polysaccharides
4.4. Comparison of the Growth and Denitrification Capacity of Three Denitrifying Bacteria
4.5. Effects of Seaweed Polysaccharides on the Growth and Denitrification Capacity of Bacillus subtilis (BS)
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, W.; Lin, X.; Shi, W.; Min, J.; Gao, N.; Zhang, H.; Yin, R.; He, X. Higher rates of nitrogen fertilization decrease soil enzyme activities, microbial functional diversity and nitrification capacity in a Chinese polytunnel greenhouse vegetable land. Plant Soil 2010, 337, 137–150. [Google Scholar] [CrossRef]
- Herold, M.B.; Baggs, L.; Daniell, T. Fungal and bacterial denitrification are differently affected by long-term pH amendment and cultivation of arable soil. Soil Biol. Biochem. 2012, 54, 25–35. [Google Scholar] [CrossRef]
- Ji, B.; Yang, K.; Zhu, L.; Jiang, Y.; Wang, H.; Zhou, J.; Zhang, H. Aerobic denitrification: A review of important advances of the last 30 years. Biotechnol. Bioprocess Eng. 2015, 20, 643–651. [Google Scholar] [CrossRef]
- Wang, J.; Chu, L. Biological nitrate removal from water and wastewater by solid-phase denitrification process. Biotechnol. Adv. 2016, 34, 1103–1112. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.C.; Cheng, Z.; Li, Y.; Tang, J. Effect of fermentation liquid from food waste as a carbon source for enhancing denitrification in wastewater treatment. Chemosphere 2016, 144, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-T.; Choi, T.-G.; Wang, H.-S.; Kim, Y.-K. Nitrate removal mediated by soil microorganism, Enterobacter sp. GG0461. J. Gen. Appl. Microbiol. 2009, 55, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, R.; Hashem, A.; Allah, E.F.A. Bacillus: A Biological Tool for Crop Improvement through Bio-Molecular Changes in Adverse Environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Orozco-Mosqueda, M.D.C.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Huang, X.; Weisener, C.G.; Ni, J.; He, B.; Xie, D.; Li, Z. Nitrate assimilation, dissimilatory nitrate reduction to ammonium, and denitrification coexist in Pseudomonas putida Y-9 under aerobic conditions. Bioresour. Technol. 2020, 312, 123597. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Yang, Q.; Shi, Y.; Xin, Y.; Zhang, L.; Gu, Z.; Shi, G. Insight into the denitrification mechanism of Bacillus subtilis JD-014 and its application potential in bioremediation of nitrogen wastewater. Process. Biochem. 2021, 103, 78–86. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, P.; Hao, B.; Yu, Z. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001. Bioresour. Technol. 2011, 102, 9866–9869. [Google Scholar] [CrossRef] [PubMed]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Herrera, R.M.H.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). Environ. Boil. Fishes 2013, 26, 619–628. [Google Scholar] [CrossRef]
- Mzibra, A.; Aasfar, A.; El Arroussi, H.; Khouloud, M.; Dhiba, D.; Kadmiri, I.M.; Bamouh, A. Polysaccharides extracted from Moroccan seaweed: A promising source of tomato plant growth promoters. Environ. Boil. Fishes 2018, 30, 2953–2962. [Google Scholar] [CrossRef]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Carrageenans from Red Seaweeds as Promoters of Growth and Elicitors of Defense Response in Plants. Front. Mar. Sci. 2016, 3, 1–9. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, F.; Wang, X.; Liu, X.; Hou, Y.; Zhang, Q. Extraction of the polysaccharides from five algae and their potential antioxidant activity in vitro. Carbohydr. Polym. 2010, 82, 118–121. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Zhao, M.; Yu, S.; Qi, H. The immunological and antioxidant activities of polysaccharides extracted from Enteromorpha linza. Int. J. Biol. Macromol. 2013, 57, 45–49. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Liu, D.; Lin, G.-P.; Wu, Y.-J.; Gao, L.-Y.; Ai, C.; Huang, Y.-F.; Wang, M.-F.; El-Seedi, H.R.; Chen, X.-H.; et al. Anti-ageing and antioxidant effects of sulfate oligosaccharides from green algae Ulva lactuca and Enteromorpha prolifera in SAMP8 mice. Int. J. Biol. Macromol. 2019, 139, 342–351. [Google Scholar] [CrossRef]
- Zhong, R.; Wan, X.; Wang, D.; Zhao, C.; Liu, D.; Gao, L.; Wang, M.; Wu, C.; Nabavid, S.M.; Daglia, M.; et al. Polysaccharides from Marine Enteromorpha: Structure and function. Trends Food Sci. Technol. 2020, 99, 11–20. [Google Scholar] [CrossRef]
- Detsch, C.; Stülke, J. Ammonium utilization in Bacillus subtilis: Transport and regulatory functions of NrgA and NrgB. Microbiology 2003, 149, 3289–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, Y.; Yano, T.; Honna, T.; Yamamoto, S.; Inosako, K. Causes of farmland salinization and remedial measures in the Aral Sea basin—Research on water management to prevent secondary salinization in rice-based cropping system in arid land. Agric. Water Manag. 2006, 85, 1–14. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, D.; Chu, S.; Zhi, Y.; Liu, X.; Zhou, P. Genomic Analysis of Bacillus megaterium NCT-2 Reveals Its Genetic Basis for the Bioremediation of Secondary Salinization Soil. Int. J. Genom. 2020, 2020, 4109186. [Google Scholar] [CrossRef] [PubMed]
- Turbé, A.; De Toni, A.; Benito, P.; Lavelle, P.; Lavelle, P.; Ruiz, N.; Van der Putten, W.H.; Labouze, E.; Mudgal, S. Soil Biodiversity: Functions, Threats and Tools for Policy Makers; Report for European Commission (DG Environment); Bio Intelligence Service, IRD and NIOO: Paris, France, 2010; pp. 4–5. [Google Scholar]
- Yang, T.; Xin, Y.; Zhang, L.; Gu, Z.; Li, Y.; Ding, Z.; Shi, G. Characterization on the aerobic denitrification process of Bacillus strains. Biomass Bioenergy 2020, 140, 105677. [Google Scholar] [CrossRef]
- Zhao, B.; Cheng, D.Y.; Tan, P.; An, Q.; Guo, J.S. Characterization of an aerobic denitrifier Pseudomonas stutzeri strain XL-2 to achieve efficient nitrate removal. Bioresour. Technol. 2018, 250, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, X.-H.; Cui, S.; Ren, Y.-X.; Yu, J.; Chen, N.; Xiao, Q.; Guo, L.-K.; Wang, R.-H. Simultaneous removal of nitrogen and phosphorous by heterotrophic nitrification-aerobic denitrification of a metal resistant bacterium Pseudomonas putida strain NP5. Bioresour. Technol. 2019, 285, 121360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Chen, H.; Lyu, Y.; Wang, Y. Nitrogen removal by a metal-resistant bacterium, Pseudomonas putida ZN1, capable of heterotrophic nitrification–aerobic denitrification. J. Chem. Technol. Biotechnol. 2019, 94, 1165–1175. [Google Scholar] [CrossRef]
- Wang, X.; Xia, K.; Yang, X.; Tang, C. Growth strategy of microbes on mixed carbon sources. Nat. Commun. 2019, 10, 1279. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sun, Y.; Hu, L.; Liu, S.; Yu, H.; Xing, R.; Li, R.; Wang, X.; Li, P. In vitro prebiotic effects of seaweed polysaccharides. J. Oceanol. Limnol. 2018, 36, 926–932. [Google Scholar] [CrossRef]
- Kong, Q.; Dong, S.; Gao, J.; Jiang, C. In vitro fermentation of sulfated polysaccharides from E. prolifera and L. japonica by human fecal microbiota. Int. J. Biol. Macromol. 2016, 91, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, T.; Ueda, N.; Kitagawa, M.; Hanke, G.; Suzuki, A.; Hase, T.; Sakakibara, H. Arabidopsis Root-Type Ferredoxin:NADP(H) Oxidoreductase 2 is Involved in Detoxification of Nitrite in Roots. Plant Cell Physiol. 2016, 57, 2440–2450. [Google Scholar] [CrossRef] [Green Version]
- Oke, O.L. Nitrite Toxicity to Plants. Nat. Cell Biol. 1966, 212, 528. [Google Scholar] [CrossRef]
- Buresh, R.J.; Patrick, W.H. Nitrate Reduction to Ammonium in Anaerobic Soil. Soil Sci. Soc. Am. J. 1978, 42, 913–918. [Google Scholar] [CrossRef]
- Silver, W.L.; Herman, D.J.; Firestone, M.K. Dissimilatory nitrate reduction to ammonium in upland tropical forest soils. Ecology 2001, 82, 2410–2416. [Google Scholar] [CrossRef]
- Li, S.-X.; Wang, Z.-H.; Stewart, B. Responses of Crop Plants to Ammonium and Nitrate N. Adv. Agron. 2013, 205–397. [Google Scholar] [CrossRef]
- Song, L.; Chen, X.; Liu, X.; Zhang, F.; Hu, L.; Yue, Y.; Li, K.; Li, P. Characterization and Comparison of the Structural Features, Immune-Modulatory and Anti-Avian Influenza Virus Activities Conferred by Three Algal Sulfated Polysaccharides. Mar. Drugs 2015, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.Y.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F.G. A Colorimetric Method for the Determination of Sugars. Nat. Cell Biol. 1951, 168, 167. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Q.; Wang, J.; Shi, X.; Zhang, Z. Analysis of the monosaccharide composition of fucoidan by precolumn derivation HPLC. Chin. J. Oceanol. Limnol. 2009, 27, 578–582. [Google Scholar] [CrossRef]
- SEPA. Methods for Monitoring and Analysis of Water and Wastewater, 4th ed.; China Environmental Science Press: Beijing, China, 2002; pp. 250–285. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Sugar (%) | Sulfate (%) | Mw (kDa) | TC (%) | Composition of Monosaccharide (in Molar Ratio) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rha | GlcUA | Xyl | Glc | Gal | Man | Fuc | GulA | ManA | |||||
| EP | 96.50 ± 1.31 | 14.07 | 756 | 31.23% | 1.00 | 0.49 | 0.26 | 0.08 | 0.04 | 0.02 | 0.02 | - | - |
| CA | - | 10.05 | 1588 | 34.06% | - | - | - | 0.61 | 0.61 | 1.00 | - | - | - |
| AL | - | - | 833 | 27.16% | 0.41 | - | - | 0.51 | - | - | - | 0.15 | 1.00 |
| BS | PP | PS | |
|---|---|---|---|
| a | 0.82 | 0.72 | 0.63 |
| tc | 12.88 | 15.78 | 16.89 |
| k | 0.32 | 0.30 | 0.27 |
| R2 | 0.99 | 0.97 | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Song, L.; Chen, X.; Li, P. An Exploration of Seaweed Polysaccharides Stimulating Denitrifying Bacteria for Safer Nitrate Removal. Molecules 2021, 26, 3390. https://doi.org/10.3390/molecules26113390
Zhang H, Song L, Chen X, Li P. An Exploration of Seaweed Polysaccharides Stimulating Denitrifying Bacteria for Safer Nitrate Removal. Molecules. 2021; 26(11):3390. https://doi.org/10.3390/molecules26113390
Chicago/Turabian StyleZhang, Hui, Lin Song, Xiaolin Chen, and Pengcheng Li. 2021. "An Exploration of Seaweed Polysaccharides Stimulating Denitrifying Bacteria for Safer Nitrate Removal" Molecules 26, no. 11: 3390. https://doi.org/10.3390/molecules26113390
APA StyleZhang, H., Song, L., Chen, X., & Li, P. (2021). An Exploration of Seaweed Polysaccharides Stimulating Denitrifying Bacteria for Safer Nitrate Removal. Molecules, 26(11), 3390. https://doi.org/10.3390/molecules26113390