
New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene



Abstract
:1. Introduction
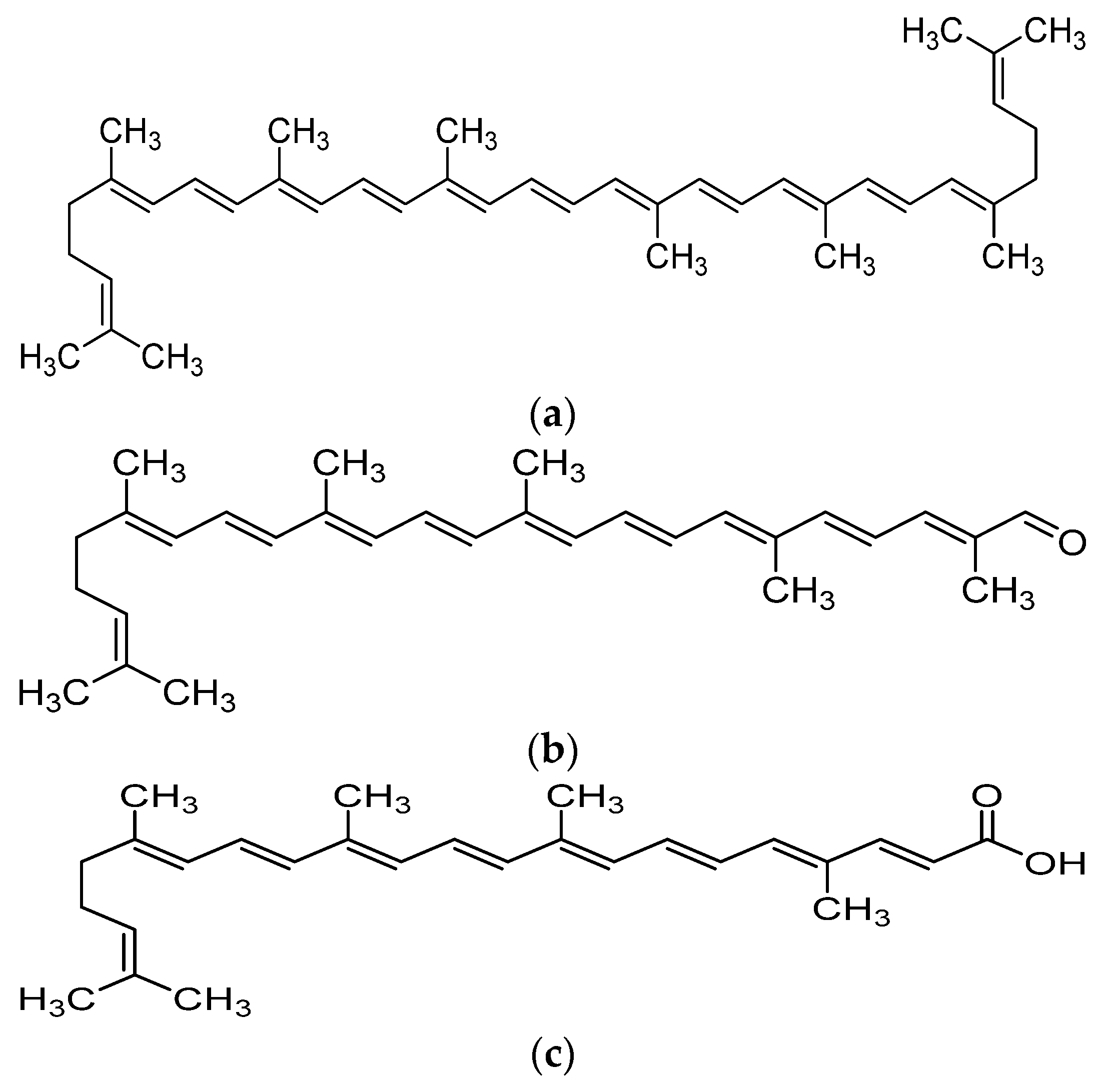
2. Metabolism and Bioavailability of Lycopene
3. Role of Inflammation in Cancer Development and Progression
4. Cancer Suppression through Immunomodulation
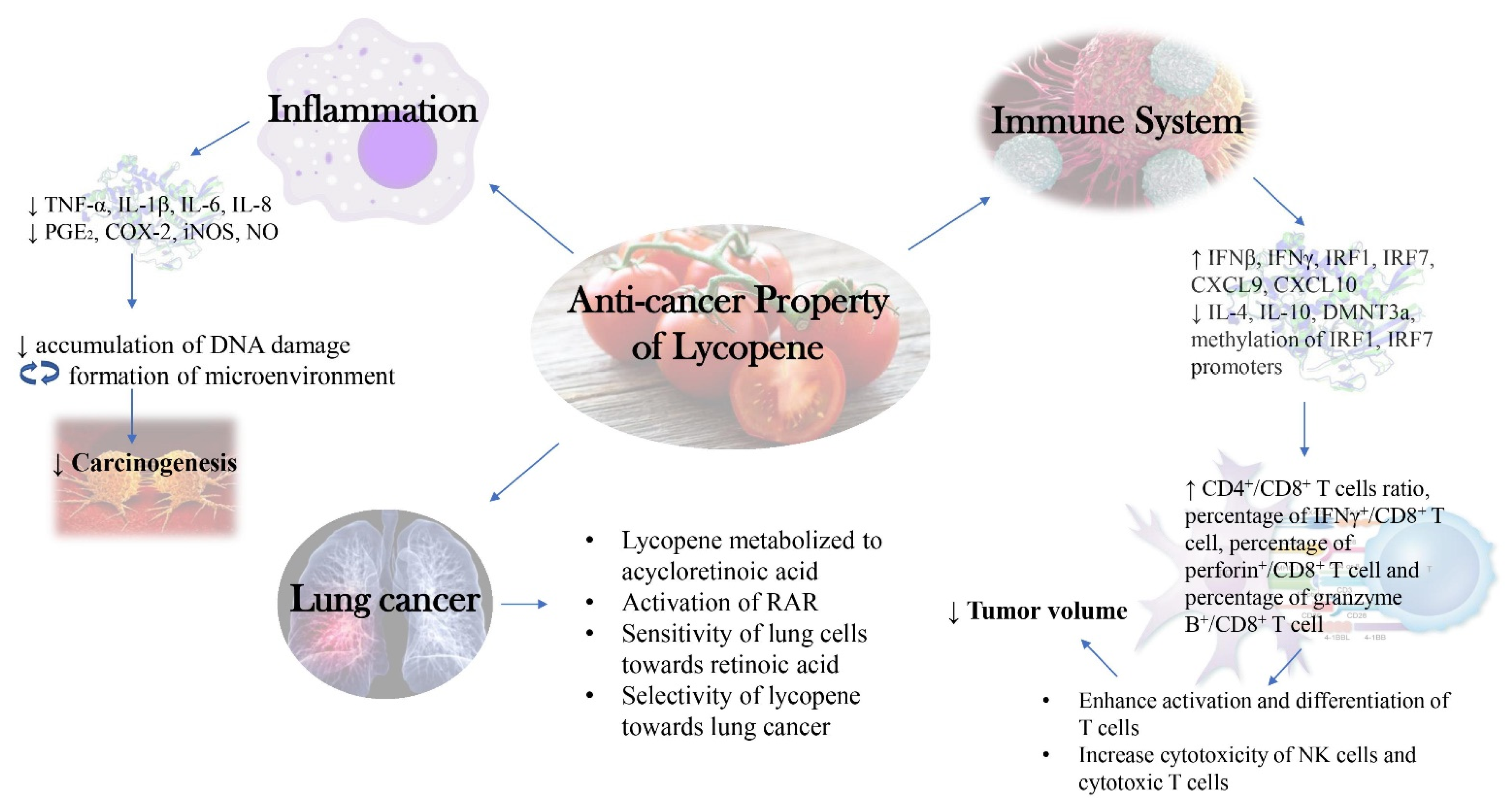
5. Anti-Inflammatory Activities of Lycopene
Utilizing Anti-Inflammatory Activity of Lycopene against Cancers
6. Anti-Cancer Activities of Lycopene
6.1. Cell Culture Studies
6.2. Animal Studies
6.3. Clinical Trials
7. Immunomodulatory Effects of Lycopene
8. Effects of Lycopene on Oxidative Stress and Liver Enzymes
9. Selective Anticancer Activities of Lycopene
9.1. Carotenoids of the Same Kind with Different Fate (Lycopene and Beta-Carotene)
9.2. Lycopene Is Selective against Lung Adenomas and Carcinomas
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SR-B1 | Scavenger receptor class B type 1 |
| CD36 | Cluster of differentiation 36 |
| ISX | Intestine-specific homeobox |
| BCO1 | β-carotene oxygenase 1 |
| IOM | Institute of Medicine |
| GRAS | Generally recognized as safe |
| MHC II | Major histocompatibility complex II |
| IL-2 | Interleukin-2 |
| MAPK | Mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated kinases |
| JNK | c-Jun N-terminal kinases |
| NF-κB | nuclear factor kappa-light-chain-enhance of activated B cells |
| LPS | Lipopolysaccharides |
| ICAM-1 | Intercellular adhesion molecule 1 |
| VCAM-1 | Vascular cell adhesion molecule 1 |
| HUVEC | Human umbilical vein endothelial cell |
| TNF-α | Tumor necrosis factor-alpha |
| ONOO− | Peroxynitrite |
| MDA | Malondialdehyde |
| NO | Nitric oxide |
| ROS | Reactive oxygen species |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| ARE | Antioxidant response element |
| p-IκB | Phosphorylation of IκB |
| NFE2L2 | Nuclear factor erythroid derived 2-like 2 |
| HMOX1 | Heme-oxygenase 1 gene |
| NQO-1 | NAD(P)H dehydrogenase [quinone] 1 |
| c-AMP | Cyclic adenosine monophosphate |
| MMPs | Matrix metalloproteinases |
| iNOS | Inducible nitric oxide synthase |
| COX-2 | Cyclooxygenase-2 |
| PGE2 | Prostaglandin E2 |
| Lyc | Lycopene |
| ET-1 | Endothelin 1 |
| MCP-1 | Monocyte chemoattractant protein-1 |
| RelA | Nuclear factor NF-κB p65 subunit |
| MIP-2 | Macrophage Inflammatory Protein 2 |
| DSS | Dextran sulfate sodium |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
| EGFP | Enhanced green fluorescent protein |
| AST | Aspartate Aminotransferase |
| ALT | Alanine aminotransferase |
| LDH | Lactate dehydrogenase; |
| GGT | Gamma-glutamyl transferase |
| RAGE | Receptor for advanced glycation endproducts |
| eWAT | epididymal white adipose tissue |
| ATM | Adipose tissue macrophage |
| STAT6 | Signal transducer and activator of transcription 6 |
| Akt | Protein kinase B |
| LDL | Low-density lipoprotein |
| CRP | C-reactive protein |
| CAT | Catalase |
| SOD | Superoxide dismutase |
| GPx | Glutathione peroxidase 1 |
| TGF-β1 | Transforming growth factor beta 1 |
| Cyt c | Cytochrome c |
| GSH | Glutathione |
| HO-1 | Heme-oxygenase 1 |
| GCLC | Glutamate-cysteine ligase catalytic subunit |
| BSA | Bovine serum albumin |
| IgG | Immunoglobulin G |
| CFA | Complete Freund’s adjuvant |
| SAPK | Stress-activated kinases |
| BW | Body weight |
| CYP2E1 | Cytochrome P450 2E1 |
| IFNγ | Interferon γ |
| MPO | Myeloperoxidase |
| HC | Hyperhomocysteinemic control group |
| ICV | Intracerebroventricular |
| L-NAME | L-NG-Nitro arginine methyl ester |
| SAH | Subarachnoid hemorrhage |
| RAI | Radioactive iodine |
| DM | Diabetes mellitus |
| CP/CPPS | Chronic prostatitis/chronic pelvic pain syndrome |
| TLE | Tomato lycopene extract |
| Th1 | Th1 helper cells |
| TLR | Toll-like receptor |
| NAD(P)H | Reduced form of nicotinamide adenine dinucleotide phosphate |
| 4HNE | 4-hydroxynonenal |
| RAR | retinoic acid receptor |
| ATRA | all-trans retinoic acid |
| RARE | retinoic acid response elements |
References
- Hassanpour, S.H.; Dehghani, M. Review of cancer from perspective of molecular. J. Cancer Res. Pract. 2017, 4, 127–129. [Google Scholar] [CrossRef]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment (Review). Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef]
- Mohan, G.; TP, A.H.; AJ, J.; KM, S.D.; Narayanasamy, A.; Vellingiri, B. Recent advances in radiotherapy and its associated side effects in cancer—A review. J. Basic Appl. Zool. 2019, 80. [Google Scholar] [CrossRef]
- Subramaniam, S.; Selvaduray, K.R.; Radhakrishnan, A.K. Bioactive compounds: Natural defense against cancer? Biomolecules 2019, 9, 758. [Google Scholar] [CrossRef] [Green Version]
- Siti, H.N.; Jalil, J.; Asmadi, A.Y.; Kamisah, Y. Roles of rutin in cardiac remodeling. J. Funct. Foods 2020, 64, 103606. [Google Scholar] [CrossRef]
- Gui, J.S.; Jalil, J.; Jubri, Z.; Kamisah, Y. Parkia speciosa empty pod extract exerts anti-inflammatory properties by modulating NFκB and MAPK pathways in cardiomyocytes exposed to tumor necrosis factor-α. Cytotechnology 2019, 71, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Jalil, J.; Attiq, A.; Hui, C.C.; Yao, L.J.; Zakaria, N.A. Modulation of inflammatory pathways, medicinal uses and toxicities of uvaria species: Potential role in the prevention and treatment of inflammation. Inflammopharmacology 2020, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Mohd Aluwi, M.F.F.; Rullah, K.; Haque, M.A.; Yamin, B.M.; Ahmad, W.; Amjad, M.W.; Leong, S.W.; Fahmizar, N.A.; Jalil, J.; Abas, F.; et al. Suppression of PGE2 production via disruption of MAPK Phosphorylation by unsymmetrical dicarbonyl curcumin derivatives. Med. Chem. Res. 2017, 26, 3323–3335. [Google Scholar] [CrossRef]
- Rowles, J.L., III; John, W.; Erdman, J. Carotenoids and their role in cancer prevention. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158613. [Google Scholar] [CrossRef]
- Mehta, N.; Patani, P.; Singhvi, I. A review on tomato lycopene. Int. J. Pharm. Sci. Res. 2018, 9, 916–923. [Google Scholar] [CrossRef]
- Zechmeister, L.; LeRosen, A.L.; Went, F.W.; Pauling, L. Prolycopene, a naturally occuring stereoisomer of lycopene. Proc. Natl. Acad. Sci. USA 1941, 27, 468–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.L.; Schwartz, S.J. Lycopene: Chemical and biological properties. Food Technol. 1999, 53, 38–45. [Google Scholar]
- Arathi, B.P.; Sowmya, P.R.-R.; Vijay, K.; Baskaran, V.; Lakshminarayana, R. Biofunctionality of carotenoid metabolites: An insight into qualitative and quantitative analysis. In Metabolomics Fundamentals and Applications; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Lian, F.; Smith, D.E.; Ernst, H.; Russell, R.M.; Wang, X.D. Apo-10′-lycopenoic acid inhibits lung cancer cell growth in vitro, and suppresses lung tumorigenesis in the A/J mouse model in vivo. Carcinogenesis 2007, 28, 1567–1574. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.M.; Huang, S.M.; Liu, C.L.; Hu, M.L. Apo-8′-lycopenal induces expression of HO-1 and NQO-1 via the ERK/P38-Nrf2-ARE pathway in human HepG2 Cells. J. Agric. Food Chem. 2012, 60, 1576–1585. [Google Scholar] [CrossRef]
- Huang, C.-S.; Shih, M.-K.; Chuang, C.-H.; Hu, M.-L. Lycopene inhibits cell migration and invasion and upregulates Nm23-H1 in a highly invasive hepatocarcinoma, SK-Hep-1 Cells. J. Nutr. 2005, 135, 2119–2123. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.S.; Fan, Y.E.; Lin, C.Y.; Hu, M.L. Lycopene inhibits matrix metalloproteinase-9 expression and down-regulates the binding activity of nuclear factor-kappa B and stimulatory protein-1. J. Nutr. Biochem. 2007, 18, 449–456. [Google Scholar] [CrossRef]
- Yang, C.M.; Hu, T.Y.; Hu, M.L. Antimetastatic effects and mechanisms of apo-8ʹ-lycopenal, an enzymatic metabolite of lycopene, against human hepatocarcinoma SK-Hep-1 Cells. Nutr. Cancer 2012, 64, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Jhou, B.Y.; Song, T.Y.; Lee, I.; Hu, M.L.; Yang, N.C. Lycopene inhibits metastasis of human liver adenocarcinoma SK-Hep-1 cells by downregulation of NADPH oxidase 4 protein expression. J. Agric. Food Chem. 2017, 65, 6893–6903. [Google Scholar] [CrossRef]
- Koh, M.S.; Hwang, J.S.; Moon, A. Lycopene inhibits proliferation, invasion and migration of human breast cancer cells. Biomol. Ther. 2010, 18, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.C.; Wang, F.Y.; Kuo, Y.H.; Tang, F.Y. Cancer chemopreventive effects of lycopene: Suppression of MMP-7 expression and cell invasion in human colon cancer cells. J. Agric. Food Chem. 2011, 59, 11304–11318. [Google Scholar] [CrossRef]
- Huang, C.-S.; Liao, J.-W.; Hu, M.-L. Lycopene inhibits experimental metastasis of human hepatoma SK-Hep-1 cells in athymic nude mice. J. Nutr. 2008, 138, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, M.; Ono, M.; Higuchi, T.; Chen, C.; Hara, T.; Nakano, S. Anti-proliferative and apoptosis-inducing activity of lycopene against three subtypes of human breast cancer cell lines. Cancer Sci. 2014, 105, 252–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.; Lim, J.W.; Kim, H. Lycopene Inhibits reactive oxygen species-mediated Nf-Kb signaling and induces apoptosis in pancreatic cancer cells. Nutrients 2019, 11, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ip, B.C.; Hu, K.Q.; Liu, C.; Smith, D.E.; Obin, M.S.; Ausman, L.M.; Wang, X.D. Lycopene metabolite, apo-10-lycopenoic acid, inhibits diethylnitrosamine-initiated, high fat diet-promoted hepatic inflammation and tumorigenesis in mice. Cancer Prev. Res. 2013, 6, 1304–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velmurugan, B.; Nagini, S. Combination chemoprevention of experimental gastric carcinogenesis by S-allylcysteine and lycopene: Modulatory effects on glutathione redox cycle antioxidants. J. Med. Food 2005, 8, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D. Lycopene metabolism and its biological significance. Am. J. Clin. Nutr. 2012, 96, 1214S–1222S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Srivastava, A.K. Lycopene; Chemistry, Biosynthesis, Metabolism and Degradation under Various Abiotic Parameters. J. Food Sci. Technol. 2015, 41–53. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Uptake of lycopene and its geometrical isomers is greater from heat- processed than from unprocessed tomato juice in humans. J. Nutr. 1992, 122, 2161–2166. [Google Scholar] [CrossRef]
- Gärtner, C.; Stahl, W.; Sies, H. Lycopene is more bioavailable from tomato paste than from fresh tomatoes. Am. J. Clin. Nutr. 1997, 66, 116–122. [Google Scholar] [CrossRef]
- Riedl, J.; Linseisen, J.; Hoffmann, J.; Wolfram, G. Some dietary fibers reduce the absorption of carotenoids in women. J. Nutr. 1999, 129, 2170–2176. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Vijayalekshmi, R.V.; Sung, B. Targeting inflammatory pathways for prevention and therapy of cancer: Short-term friend, long-term foe. Clin. Cancer Res. 2009, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Giroux, V.; Rustgi, A.K. Metaplasia: Tissue injury adaptation and a precursor to the dysplasia-cancer sequence. Nat. Rev. Cancer 2017, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Cordon-Cardo, C.; Prives, C. At the crossroads of inflammation and tumorigenesis. J. Exp. Med. 1999, 1367–1370. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Akaike, T. Nitric oxide and oxygen radicals in infection, inflammation, and cancer. Biokhimiya 1998, 63, 1007–1019. [Google Scholar]
- Smyth, M.J.; Cretney, E.; Kershaw, M.H.; Hayakawa, Y. Cytokines in cancer immunity and immunotherapy. Immunol. Rev. 2004, 275–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, Y. Tumor necrosis factor and cancer, buddies or foes? Acta Pharmacol. Sin. 2008, 1275–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumor Biol. 2016, 11553–11572. [Google Scholar] [CrossRef]
- Negus, R.P.M.; Stamp, G.W.H.; Relf, M.G.; Burke, F.; Malik, S.T.A.; Bernasconi, S.; Allavena, P.; Sozzani, S.; Mantovani, A.; Balkwill, F.R. The detection and localization of monocyte chemoattractant protein-1 (MCP-1) in human ovarian cancer. J. Clin. Investig. 1995, 95, 2391–2396. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Baby, D.; Rajguru, J.; Patil, P.; Thakkannavar, S.; Pujari, V. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Oncology meets immunology: The cancer-immunity cycle. Immunity 2013, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dunn, G.P.; Koebel, C.M.; Schreiber, R.D. Interferons, immunity and cancer immunoediting. Nat. Rev. Immunol. 2006, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Pio, R.; Corrales, L.; Lambris, J.D. The role of complement in tumor growth. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 772, pp. 229–262. [Google Scholar] [CrossRef] [Green Version]
- Waldhauer, I.; Steinle, A. NK cells and cancer immunosurveillance. Oncogene 2008, 27, 5932–5943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrington, R.; Watson, W.; Kim, H.L.; Antonetti, F.R. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2011, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, T.J.; Drake, C.G. Primer on tumor immunology and cancer immunotherapy. J. Immunother. Cancer 2013. [Google Scholar] [CrossRef] [Green Version]
- Minami, Y.; Kono, T.; Miyazaki, T.; Taniguchi, T. The IL-2 receptor complex: Its structure, function, and target genes. Annu. Rev. Immunol. 1993, 11, 245–268. [Google Scholar] [CrossRef]
- NORMAN, P. Immunobiology: The immune system in health and disease. J. Allergy Clin. Immunol. 1995, 96, 274. [Google Scholar] [CrossRef]
- Schroeder, H.W.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.C.; Lanier, L.L. Natural killer cells remember: An evolutionary bridge between innate and adaptive immunity? Eur. J. Immunol. 2009, 39, 2059–2064. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.N.; Liu, Y.B.; Li, B.H. Lycopene exerts anti-inflammatory effect to inhibit prostate cancer progression. Asian J. Androl. 2019, 21, 80–85. [Google Scholar] [CrossRef]
- Li, C.C.; Liu, C.; Fu, M.; Hu, K.Q.; Aizawa, K.; Takahashi, S.; Hiroyuki, S.; Cheng, J.; von Lintig, J.; Wang, X.D. Tomato powder inhibits hepatic steatosis and inflammation potentially through restoring SIRT1 activity and adiponectin function independent of carotenoid cleavage enzymes in mice. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef]
- Sun, X.; Jia, H.; Xu, Q.; Zhao, C.; Xu, C. Lycopene alleviates H2O2-induced oxidative stress, inflammation and apoptosis in bovine mammary epithelial cells: Via the NFE2L2 signaling pathway. Food Funct. 2019, 10, 6276–6285. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Yang, F.; Meng, L.; Chen, D.; Wang, M.; Lu, X.; Chen, D.; Jiang, Y.; Xing, N. Lycopene attenuates chronic prostatitis/chronic pelvic pain syndrome by inhibiting oxidative stress and inflammation via the interaction of NF-ΚB, MAPKs, and Nrf2 signaling pathways in rats. Andrology 2020, 8, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.H.; Kim, W.K.; Ha, A.W.; Kim, M.H.; Chang, M.J. Anti-inflammatory effect of lycopene in SW480 human colorectal cancer cells. Nutr. Res. Pract. 2017, 11, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Quagliariello, V.; Vecchione, R.; Coppola, C.; Di Cicco, C.; De Capua, A.; Piscopo, G.; Paciello, R.; Narciso, V.; Formisano, C.; Taglialatela-Scafati, O.; et al. Cardioprotective effects of nanoemulsions loaded with anti-inflammatory nutraceuticals against doxorubicin-induced cardiotoxicity. Nutrients 2018, 10, 1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricciotti, E.; Fitzgerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the war against inflammation with natural products. Front. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 252–259. [Google Scholar] [CrossRef]
- Sengupta, A.; Ghosh, S.; Das, R.K.; Bhattacharjee, S.; Bhattacharya, S. Chemopreventive potential of diallylsulfide, lycopene and theaflavin during chemically induced colon carcinogenesis in rat colon through modulation of cyclooxygenase-2 and inducible nitric oxide synthase pathways. Eur. J. Cancer Prev. 2006, 15, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Wu, X.G. Lycopene enhances antioxidant enzyme activities and immunity function in N-Methyl-N′-Nitro-N-nitrosoguanidine-induced gastric cancer rats. Int. J. Mol. Sci. 2011, 12, 3340–3351. [Google Scholar] [CrossRef] [Green Version]
- Ip, B.C.; Liu, C.; Ausman, L.M.; Von Lintig, J.; Wang, X.D. Lycopene attenuated hepatic tumorigenesis via differential mechanisms depending on carotenoid cleavage enzyme in mice. Chest 2014, 146, 1219–1227. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.K.; Zhang, R.L.; Bi, T.N.; Lu, Y.; Jiang, L.X. Inhibitory effect of lycopene against the growth of human gastric cancer cells. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Gajowik, A.; Dobrzyńska, M.M. The evaluation of protective effect of lycopene against genotoxic influence of X-irradiation in human blood lymphocytes. Radiat. Environ. Biophys. 2017, 56, 413–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wu, Y.Y.; Wang, X.; Shen, P.; Jia, Q.; Yu, S.; Wang, Y.; Li, X.; Chen, W.; Wang, A.; et al. Lycopene prevents carcinogen-induced cutaneous tumor by enhancing activation of the Nrf2 pathway through P62-triggered autophagic keap1 degradation. Aging 2020, 12, 8167–8190. [Google Scholar] [CrossRef] [PubMed]
- Lian, F.; Wang, X.D. Enzymatic metabolites of lycopene induce Nrf2-mediated expression of phase II detoxifying/antioxidant enzymes in human bronchial epithelial cells. Int. J. Cancer 2008, 123, 1262–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasawa, H.; Mitamura, T.; Sakamoto, S.; Yamamoto, K. Effects of lycopene on spontaneous mammary tumour development in SHN virgin mice. Anticancer Res. 1995, 15, 1173–1178. [Google Scholar] [PubMed]
- Sharoni, Y.; Giron, E.; Rise, M.; Levy, J. Effects of lycopene-enriched tomato oleoresin on 7,12-dimethyl-benz[a]anthracene-induced rat mammary tumors. Cancer Detect. Prev. 1997, 21, 118–123. [Google Scholar]
- Jiang, X.; Wu, H.; Zhao, W.; Ding, X.; You, Q.; Zhu, F.; Qian, M.; Yu, P. Lycopene improves the efficiency of anti-PD-1 therapy via activating IFN signaling of lung cancer cells. Cancer Cell Int. 2019, 19. [Google Scholar] [CrossRef] [Green Version]
- Astorg, P.; Gradelet, S.; Bergès, R.; Suschetet, M. Dietary lycopene decreases the initiation of liver preneoplastic foci by diethylnitrosamine in the rat. Nutr. Cancer 1997, 29, 60–68. [Google Scholar] [CrossRef]
- Passos Toledo, L.; Prates Ong, T.; Galvão Pinho, A.L.; Jordão, A.; Vanucchi, H.; Salvador Moreno, F. Inhibitory effects of lutein and lycopene on placental glutathione s-transferase-positive preneoplastic lesions and DNA strand breakage induced in wistar rats by the resistant hepatocyte model of hepatocarcinogenesis. Nutr. Cancer 2003, 47, 62–69. [Google Scholar] [CrossRef]
- Wang, Y.; Ausman, L.M.; Greenberg, A.S.; Russell, R.M.; Wang, X.D. Dietary lycopene and tomato extract supplementations inhibit nonalcoholic steatohepatitis-promoted hepatocarcinogenesis in rats. Int. J. Cancer 2010, 126, 1788–1796. [Google Scholar] [CrossRef] [Green Version]
- Velmurugan, B.; Bhuvaneswari, V.; Burra, U.K.; Nagini, S. Prevention of N-Methyl-N′-Nitro-N-nitrosoguanidine and saturated sodium chloride-induced gastric carcinogenesis in wistar rats by lycopene. Eur. J. Cancer Prev. 2002, 11, 19–26. [Google Scholar] [CrossRef]
- Ucci, M.; Di Tomo, P.; Tritschler, F.; Cordone, V.G.P.; Lanuti, P.; Bologna, G.; Di Silvestre, S.; Di Pietro, N.; Pipino, C.; Mandatori, D.; et al. Anti-inflammatory role of carotenoids in endothelial cells derived from umbilical cord of women affected by gestational diabetes mellitus. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Holzapfel, N.P.; Shokoohmand, A.; Wagner, F.; Landgraf, M.; Champ, S.; Holzapfel, B.M.; Clements, J.A.; Hutmacher, D.W.; Loessner, D. Lycopene reduces ovarian tumor growth and intraperitoneal metastatic load. Am. J. Cancer Res. 2017, 7, 1322–1336. [Google Scholar]
- Sahin, K.; Yenice, E.; Tuzcu, M.; Orhan, C.; Mizrak, C.; Ozercan, I.H.; Sahin, N.; Yilmaz, B.; Bilir, B.; Ozpolat, B.; et al. Lycopene protects against spontaneous ovarian cancer formation in laying hens. J. Cancer Prev. 2018, 23, 25–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narisawa, T.; Fukaura, Y.; Hasebe, M.; Ito, M.; Aizawa, R.; Murakoshi, M.; Uemura, S.; Khachik, F.; Nishino, H. Inhibitory effects of natural carotenoids, α-carotene, β-carotene, lycopene and lutein, on colonic aberrant crypt foci formation in rats. Cancer Lett. 1996, 107, 137–142. [Google Scholar] [CrossRef]
- Narisawa, T.; Fukaura, Y.; Hasebe, M.; Nomura, S.; Oshima, S.; Sakamoto, H.; Inakuma, T.; Ishiguro, Y.; Takayasu, J.; Nishino, H. Prevention of N-methylnitrosourea-induced colon carcinogenesis in F344 rats by lycopene and tomato juice rich in lycopene. Jpn. J. Cancer Res. 1998, 89, 1003–1008. [Google Scholar] [CrossRef]
- Valadez-Bustos, N.; Escamilla-Silva, E.M.; García-Vázquez, F.J.; Gallegos-Corona, M.A.; Amaya-Llano, S.L.; Ramos-Gómez, M. Oral Administration of Microencapsulated, B. Longum BAA-999 and lycopene modulates IGF-1/IGF-1R/IGFBP3 protein expressions in a colorectal murine model. Int. J. Mol. Sci. 2019, 20, 4275. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.J.; Takasuka, N.; Kim, J.M.; Sekine, K.; Ota, T.; Asamoto, M.; Murakoshi, M.; Nishino, H.; Nir, Z.; Tsuda, H. Chemoprevention by lycopene of mouse lung neoplasia after combined initiation treatment with DEN, MNU and DMH. Cancer Lett. 1997, 120, 15–22. [Google Scholar] [CrossRef]
- Watanabe, S.; Kitade, Y.; Masaki, T.; Nishioka, M.; Satoh, K.; Nishino, H. Effects of lycopene and sho-saiko-to on hepatocarcinogenesis in a rat model of spontaneous liver cancer. Nutr. Cancer 2001, 39, 96–101. [Google Scholar] [CrossRef]
- Cohen, L.A.; Zhao, Z.; Pittman, B.; Khachik, F. Effect of dietary lycopene on N-methylnitrosourea-induced mammary tumorigenesis. Nutr. Cancer 1999, 34, 153–159. [Google Scholar] [CrossRef]
- Giovannucci, E.; Ascherio, A.; Rimm, E.B.; Stampfer, M.J.; Colditz, G.A.; Willett, W.C. Intake of carotenoids and retino in relation to risk of prostate cancer. J. Natl. Cancer Inst. 1995, 87, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E. A prospective study of tomato products, lycopene, and prostate cancer risk. CancerSpectrum Knowl. Environ. 2002, 94, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Erdman, J.W.; Schwartz, S.J.; Platz, E.A.; Leitzmann, M.; Clinton, S.K.; DeGroff, V.; Willett, W.C.; Giovannucci, E. Plasma and dietary carotenoids, and the risk of prostate cancer: A nested case-control study. Cancer Epidemiol. Biomark. Prev. 2004, 13, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucuk, O.; Sarkar, F.H.; Sakr, W.; Djuric, Z.; Pollak, M.N.; Khachik, F.; Li, Y.W.; Banerjee, M.; Grignon, D.; Bertram, J.S.; et al. Phase II randomized clinical trial of lycopene supplementation before radical prostatectomy. Cancer Epidemiol. Biomark. Prev. 2001, 10, 861–868. [Google Scholar]
- Bowen, P.; Chen, L.; Stacewicz-Sapuntzakis, M.; Duncan, C.; Sharifi, R.; Ghosh, L.; Kim, H.S.; Christov-Tzelkov, K.; Van Breemen, R. Tomato sauce supplementation and prostate cancer: Lycopene accumulation and modulation of biomarkers of carcinogenesis. Exp. Biol. Med. 2002, 227, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Paur, I.; Lilleby, W.; Bøhn, S.K.; Hulander, E.; Klein, W.; Vlatkovic, L.; Axcrona, K.; Bolstad, N.; Bjøro, T.; Laake, P.; et al. Tomato-based randomized controlled trial in prostate cancer patients: Effect on PSA. Clin. Nutr. 2017, 36, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Song, Y.; Zhang, L. Lycopene/tomato consumption and the risk of prostate cancer: A systematic review and meta-analysis of prospective studie. J. Nutr. Sci. Vitaminol. 2013, 59, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Zhang, W.; Wang, X.; Zhao, K.; Negi, D.S.; Zhuo, L.; Qi, M.; Wang, X.; Zhang, X. Lycopene and risk of prostate cancer. Medicine 2015, 94, e1260. [Google Scholar] [CrossRef]
- Bae, J.M. Reinterpretation of the results of a pooled analysis of dietary carotenoid intake and breast cancer risk by using the interval collapsing method. Epidemiol. Health 2016, 38, e2016024. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Lu, M.S.; Wang, L.; Mo, X.F.; Luo, W.P.; Du, Y.F.; Zhang, C.X. Specific serum carotenoids are inversely associated with breast cancer risk among chinese women: A case-control study. Br. J. Nutr. 2016, 115, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Abar, L.; Vieira, A.R.; Aune, D.; Stevens, C.; Vingeliene, S.; Navarro Rosenblatt, D.A.; Chan, D.; Greenwood, D.C.; Norat, T. Blood concentrations of carotenoids and retinol and lung cancer risk: An update of the WCRF–AICR Systematic review of published prospective studies. Cancer Med. 2016, 5, 2069–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsing, A.W.; Comstock, G.W.; Abbey, H.; Polk, B.F. Serologic precursors of cancer. Retinol, carotenoids, and tocopherol and risk of prostate cancer. J. Natl. Cancer Inst. 1990, 82, 941–946. [Google Scholar] [CrossRef]
- Vogt, T.M.; Mayne, S.T.; Graubard, B.I.; Swanson, C.A.; Sowell, A.L.; Schoenberg, J.B.; Swanson, G.M.; Greenberg, R.S.; Hoover, R.N.; Hayes, R.B.; et al. Serum lycopene, other serum carotenoids, and risk of prostate cancer in US blacks and whites. Am. J. Epidemiol. 2002, 155, 1023–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuurman, A.G.; Goldbohm, R.A.; Brants, H.A.M.; Van Den Brandt, P.A. A Prospective cohort study on intake of retinol, vitamins C and E, and carotenoids and prostate cancer risk (Netherlands). Cancer Causes Control. 2002, 13, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.M.; Weinberg, V.; Magbanua, M.J.; Sosa, E.; Simko, J.; Shinohara, K.; Federman, S.; Mattie, M.; Hughes-Fulford, M.; Haqq, C.; et al. Nutritional supplements, COX-2 and IGF-1 expression in men on active surveillance for prostate cancer. Cancer Causes Control 2011, 22, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.Y.; Kim, J.H.; Ahn, S.C.; Lee, H.J.; Moon, D.O.; Lee, C.M.; Park, Y.M. Lycopene suppresses the lipopolysaccharide-induced phenotypic and functional maturation of murine dendritic cells through inhibition of mitogen-activated protein kinases and nuclear factor-ΚB. Immunology 2004, 113, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Manabe, Y.; Hirata, T.; Sugawara, T. Suppressive effects of carotenoids on the antigeninduced degranulation in RBL-2H3 rat basophilic leukemia cells. J. Oleo Sci. 2014, 63, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Fachinello, M.R.; Fernandes, N.L.M.; de Souto, E.R.; dos Santos, T.C.; da Costa, A.E.R.; Pozza, P.C. Lycopene affects the immune responses of finishing pigs. Ital. J. Anim. Sci. 2018, 17, 666–674. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, M.R. The interferon-gamma paradox in cancer. J. Interf. Cytokine Res. 2019, 39, 30–38. [Google Scholar] [CrossRef]
- Ren, W.; Gao, L.; Song, J. Structural basis of DNMT1 and DNMT3A-Mediated DNA methylation. Genes 2018, 9, 620. [Google Scholar] [CrossRef] [Green Version]
- Alsamman, K.; El-Masry, O.S. Interferon regulatory factor 1 inactivation in human cancer. Biosci. Rep. 2018, 38, BSR20171672. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Chen, W.; Zhu, W.; Meng, H.; Chen, J.; Zhang, J. Overexpression of interferon regulatory factor 7 (IRF7) reduces bone metastasis of prostate cancer cells in mice. Oncol. Res. 2017, 25, 511–522. [Google Scholar] [CrossRef]
- Makon-Sébastien, N.; Francis, F.; Eric, S.; Henri, V.P.; François, L.J.; Laurent, P.; Yves, B.; Serge, C. Lycopene modulates THP1 and Caco2 cells inflammatory state through transcriptional and nontranscriptional processes. Mediat. Inflamm. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. P62 links the autophagy pathway and the ubiqutin-proteasome system upon ubiquitinated protein degradation. Cell. Mol. Biol. Lett. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Yin, Y.; Lu, R.; Jiang, Z. Lycopene ameliorated oxidative stress and inflammation in Type 2 diabetic rats. J. Food Sci. 2019, 84, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Bignotto, L.; Rocha, J.; Sepodes, B.; Eduardo-Figueira, M.; Pinto, R.; Chaud, M.; De Carvalho, J.; Moreno, H.; Mota-Filipe, H. Anti-inflammatory effect of lycopene on carrageenan-induced paw oedema and hepatic ischaemia-reperfusion in the rat. Br. J. Nutr. 2009, 102, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Denniss, S.G.; Haffner, T.D.; Kroetsch, J.T.; Davidson, S.R.; Rush, J.W.E.; Hughson, R.L. Effect of short-term lycopene supplementation and postprandial dyslipidemia on plasma antioxidants and biomarkers of endothelial health in young, healthy individuals. Vasc. Health Risk Manag. 2008, 4, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Markovits, N.; Amotz, A.B.; Levy, Y. The effect of tomato-derived lycopene on low carotenoids and enhanced systemic inflammation and oxidation in severe obesity. Isr. Med. Assoc. J. 2009, 11, 598–601. [Google Scholar]
- Petyaev, I.M.; Dovgalevsky, P.Y.; Klochkov, V.A.; Chalyk, N.E.; Pristensky, D.V.; Chernyshova, M.P.; Udumyan, R.; Kocharyan, T.; Kyle, N.H.; Lozbiakova, M.V.; et al. Effect of lycopene supplementation on cardiovascular parameters and markers of inflammation and oxidation in patients with coronary vascular disease. Food Sci. Nutr. 2018, 6, 1770–1777. [Google Scholar] [CrossRef] [Green Version]
- Li, W.W.; Wang, T.Y.; Cao, B.; Liu, B.; Rong, Y.M.; Wang, J.J.; Wei, F.; Wei, L.Q.; Chen, H.; Liu, Y.X. Synergistic protection of matrine and lycopene against lipopolysaccharide-induced acute lung injury in mice. Mol. Med. Rep. 2019, 20, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veeramachaneni, S.; Ausman, L.M.; Choi, S.W.; Russell, R.M.; Wang, X.-D. High dose lycopene supplementation increases hepatic cytochrome P4502E1 protein and inflammation in alcohol-fed rats. J. Nutr. 2008, 138, 1329–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.J. The role of carotenoids in human health. Nutr. Clin. Care Off. Publ. Tufts Univ. 2002, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Mayne, S.T. Beta-carotene, carotenoids, and disease prevention in humans. FASEB J. 1996, 10, 690–701. [Google Scholar] [CrossRef] [Green Version]
- Arab, L.; Steck-Scott, S.; Bowen, P. Participation of lycopene and beta-carotene in carcinogenesis: Defenders, aggressors, or passive bystanders? Epidemiol. Rev. 2001, 23, 211–230. [Google Scholar] [CrossRef] [Green Version]
- The Alpha-Tocopherol Beta Carotene Cancer Prevention Study Group. The effect of vitamin e and beta carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Risk factors for lung cancer and for intervention effects in CARET, the beta-carotene and retinol efficacy trial. J. Natl. Cancer Inst. 1996, 88, 1550–1559. [Google Scholar] [CrossRef]
- Männistö, S.; Smith-Warner, S.A.; Spiegelman, D.; Albanes, D.; Anderson, K.; Van Den Brandt, P.A.; Cerhan, J.R.; Colditz, G.; Feskanich, D.; Freudenheim, J.L.; et al. Dietary carotenoids and risk of lung cancer in a pooled analysis of seven cohort studies. Cancer Epidemiol. Biomark. Prev. 2004, 13, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Touvier, M.; Kesse, E.; Clavel-Chapelon, F.; Boutron-Ruault, M.C. Dual association of β-carotene with risk of tobacco-related cancers in a cohort of french women. J. Natl. Cancer Inst. 2005, 97, 1338–1344. [Google Scholar] [CrossRef] [Green Version]
- Goralczyk, R. Beta-carotene and lung cancer in smokers: Review of hypotheses and status of research. Nutr. Cancer 2009, 61, 767–774. [Google Scholar] [CrossRef]
- Michaud, D.S.; Feskanich, D.; Rimm, E.B.; Colditz, G.A.; Speizer, F.E.; Willett, W.C.; Giovannucci, E. Intake of specific carotenoids and risk of lung cancer in 2 prospective US cohorts. Am. J. Clin. Nutr. 2000, 72, 990–997. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.V.; Agarwal, S. Effect of diet and smoking on serum lycopene and lipid peroxidation. Nutr. Res. 1998, 18, 713–721. [Google Scholar] [CrossRef]
- Heber, D. Colorful cancer prevention: α-carotene, lycopene, and lung cancer. Am. J. Clin. Nutr. 2000, 72, 901–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satia, J.A.; Littman, A.; Slatore, C.G.; Galanko, J.A.; White, E. Long-term use of β-carotene, retinol, lycopene, and lutein supplements and lung cancer risk: Results from the vitamins and lifestyle (vital) study. Am. J. Epidemiol. 2009, 169, 815–828. [Google Scholar] [CrossRef]
- Ford, N.A.; Smith, J.W.; Clinton, S.K.; Erdman, J.W. Tomato powder or lycopene reduces serum and testicular testosterone and enzymes controlling androgen and estrogen metabolism in mice lacking carotene-15,15’-monooxygenase. Exp. Biol. 2011. [Google Scholar] [CrossRef]
- Luca, L.M.D.; Darwiche, N.; Celli, G.; Kosa, K.; Jones, C.; Ross, S.; Chen, L.-C. Vitamin A in epithelial differentiation and skin carcinogenesis. Nutr. Rev. 1994, 52, S45–S52. [Google Scholar] [CrossRef]
- Fernandes-Silva, H.; Araújo-Silva, H.; Correia-Pinto, J.; Moura, R.S. Retinoic acid: A key regulator of lung development. Biomolecules 2020, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- National Research Council (US) Panel on Dosimetric Assumptions Affecting the Application of Radon Risk Estimates. Comparative Dosimetry of Radon in Mines and Homes; National Academy of Sciences: Washington, DC, USA, 1991. [Google Scholar] [CrossRef]
- Ben-Dor, A.; Nahum, A.; Danilenko, M.; Giat, Y.; Stahl, W.; Martin, H.D.; Emmerich, T.; Noy, N.; Levy, J.; Sharoni, Y. Effects of acyclo-retinoic acid and lycopene on activation of the retinoic acid receptor and proliferation of mammary cancer cells. Arch. Biochem. Biophys. 2001, 391, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Von Laar, J.; Martin, H.D.; Emmerich, T.; Sies, H. Stimulation of gap junctional communication: Comparison of acyclo- retinoic acid and lycopene. Arch. Biochem. Biophys. 2000, 373, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Li, M.T.; Richter, F.; Chang, C.; Irwin, R.J.; Huang, H.F.S. Androgen and retinoic acid interaction in LNCaP cells, effects on cell proliferation and expression of retinoic acid receptors and epidermal growth factor receptor. BMC Cancer 2002, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.C.; Hsu, S.L.; Lin, H.; Yang, T.Y. Retinoic acid and cancer treatment. BioMedicine 2014, 4, 1–6. [Google Scholar] [CrossRef]
- Alsafadi, S.; Even, C.; Falet, C.; Goubar, A.; Commo, F.; Scott, V.; Quidville, V.; Albiges, L.; Dieci, M.V.; Guegan, J.; et al. Retinoic acid receptor alpha amplifications and retinoic acid sensitivity in breast cancers. Clin. Breast Cancer 2013, 13, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, S.; Jiang, C.; Wang, Y.; Zhu, H.; Wang, X.D. High expression of RARβ is a favorable factor in colorectal cancer. Dis. Markers 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Compound | Subject | Experiment Design | Outcome | Reference |
|---|---|---|---|---|
| Lycopene (Lyc) | SK-Hep-1 cells (highly invasive hepatoma cell line) | Treatment with 1, 2.5, 5, 10, 20 µmol/L Lyc | ↓cell migration, invasion (bell-shaped manner) ↑nm23-H1 (bell-shaped manner) nm23-H1 and cell migration and invasion (-ve r) | [16] |
| Treatment with 1–10 µM Lyc | ↓cell invasion, MMP-9, NF-κB, Sp1, IGF-1R, ROS | [17] | ||
| Treatment with 1, 2.5, 5, 10 µM Apo-8′-lycopenal (Lyc metabolite), 10 µM Lyc | (Lyc, Apo-8′-lyc)↓cell invasion, migration (Apo-8′-lyc) ↓MMP-2, -9, Rho GTPase, ERK/p38, PI3K-Akt ↑nm23-H1, TIMP-1,-2 | [18] | ||
| Treatment with Lyc (0.1–5 µM), induced with TGF-β | ↓NOX4 mRNA, NOX, ROS, cell migration, invasion, adhesion activity, MMP-9, MMP-2 | [19] | ||
| H-Ras MCF10A, MDA-MB231 (highly aggressive breast cancer cell) | Treatment with Lyc | ↓cell invasion, migration, proliferation ↓ERK, Akt | [20] | |
| HT-29 cells (human colon cancer cells) | Treatment with Lyc | ↓cell invasion, MMP-7, phosphorylation of Akt, GSK-3β, ERK ½, AP-1, β-catenin ↑E-cadherin stabilization | [21] | |
| ER/PR+ MCF-7, HER2+ SK-BR-3, MDA-MB-468 cell lines | Treatment with Lyc (168 h) | inhibition of cell cycle progression G0/G1 ↑PARP cleavage, ERK1/2, p21, Bax ↓cyclin D1, Akt, mTOR ↔Bcl-xL | [23] | |
| HGC-27 cell lines | Incubated with various conc of Lyc for 24, 48 or 72 h | ↑LC3-I, p-ERK | [63] | |
| Balb/c nude mice model | Injected with HGC-27 cells, fed with 20, 30, 60 mg/kg Lyc per d, oral | ↓tumour weight | ||
| Lymphocytes from human blood | Incubation with 10, 20, 40 µM/mL Lyc, before and after X-irradiation at doses of 0.5, 1 and 2 Gy | ↓DNA damage Note: Lyc administration after irradiation, no effect Note: Low doses are useful | [64] | |
| (Pancreatic cancer) PANC-1 cells | Treatment with 0.25, 0.5 µM for 24 h | ↓ROS, NF-κB, cIAP1, cIAP2, survivin ↑caspase-3, Bax:Bcl-2 | [24] | |
| Mouse epidermal cell line, JBG P+ (JB6 Cl 41-5a) | Pretreatment with Lyc for 5 days, incubation with TPA, with or without Lyc for 14 days | ↓colony formation, (mRNA) KEAP1 ↑(mRNA) SOD1, GSR, GPX1, CAT, GCLC, GCLM, NQO-1, HMOX1, nuclear NRF2 localization, LC3, p62 | [65] | |
| Mice | Subjected to DMBA (60 µg) dissolved in 0.2 mL topically on back, after 1 week, TPA (4 µg) twice a week for 32 weeks; Control group (1), 8 µmol Lyc/d since first week (2), 8 µmol Lyc/d from first wk to 4th week only (3), 8 µmol Lyc/d since fourth week (4), Acetone/d since fourth week (5): 32 weeks experiment | ↓incidence rate, multiplicity of cutaneous papillomas, increased in epidermal thickness, invasion of benign papillomas, 8-OHdG, 4HNE ↑survival rate, GSH/GSSG ratio, SOD, GR, GPx, CAT, (mRNA) SOD1, GSR, GPX1, CAT, GCLC, GCLM, NQO-1, HMOX1 Note: lycopene was more effective as a pretreatment and during promotion phase of induced tumors.NRF2 was required for the effect of lycopene-induced prevention against tumor | ||
| Apo-10′-lycopenoic acid, A-10-LA (Lyc metabolite) | NHBE cells (human bronchial epithelial cells), BEAS-2B-immortalized normal bronchial epithelial cells, A549 (non-small cell lung cancer cells) | Treatment with apo-10′-lycopenoic acid | ↓cyclin E inhibition of cell cycle progression G(1)→S ↑p21, p27, RAR beta | [14] |
| A/J mouse model | NNK injection (induction) and supplemented (10, 40, 120 mg/kg of A-10-LA | ↓tumor multiplicity | ||
| BEAS-2B cells | Treatment with apo-10′-lycopenoic acid | ↑NRF2, HO-1, NAD(P)H dehydrogenase (quinone 1), GSTs, GCL, GSH | [66] | |
| Human liver THLE-2, HuH7 cells | Treatment with apo-10′-lycopenoic acid | ↑SIRT1, p21, apoptosis ↓cyclin D1 | ||
| C57BI/6J mice | Supplementation with A-10-LA (10 mg/kg) for 24 wks, high fat diet, induced with diethylnitrosamine | ↓tumor multiplicity, volume, incidence, caspase-1, TNF-α, IL-6, NF-κB p65, STAT3, Akt, cyclin D1 ↑SIRT1, PARP cleavage | [25] | |
| Apo-8′-lycopenal (Lyc metabolite) | Human HepG2 cells | Treatment with 1, 5, 10 µM Apo-8′-lycopenal (Lyc metabolite), 10 µM Lyc | ↓cell invasion, migration ↑NRF2, HO-1, NQO-1 ↓KEAP1 | [15] |
| Compound | Subject | Experiment Design | Outcome | Reference |
|---|---|---|---|---|
| Lycopene (Lyc) | High mammary tumor strain of SHN virgin mice | Control (1), 5 × 10−5% Lyc (2), AIN-76TM diet | ↓mammary tumor development, TYMS, serum FFA, prolactin | [67] |
| Sprague Dawley rats | N-methylnitrosourea (intrarectal, 1 wk), followed by administration of Lyc (1), lutein (2), α-carotene (3), β-carotene (4), palm carotene (5), daily gavage (wk 2 and wk 5) | ↓aberrant crypt foci development | [77] | |
| Male weanling rats | Induction of hepatocarcinogenesis by 6 × 100 mg/kg BW diethylnitrosamine (DEN)/100 mg/kg BW 2-nitropropane (2-NP), fed with 300 mg/kg β-carotene (1), canthaxanthin (2), astaxanthin (3), Lyc (4), 15,000 retinol equiv. excess vit A (5), 3-methycholanthrene (6) intraperitoneal, 3–4 wks | ↔No., size of preneoplastic liver foci ↓size of GGT+, GST+ foci, Liver volume fraction occupied by foci Note: modulate P-450 2E1, not antioxidant properties | [70] | |
| Multiorgan carcinogenesis B6C3F1 mice model | Combined treatment with diethylnitrosamine (DEN), N-methyl-N-nitrosourea (MNU) and 1,2-dimethylhydrazine (DMH), Lyc + water: 25/50 ppm (1), Control (2), Lyc only: 25/50 ppm (3), 21 wks | ↓incidences and multiplicities of lung adenomas and carcinomas Note: restricted to male, G1 with 50ppm Lyc ↔aberrant crypt loci, tumors in colon and kidney among groups | [80] | |
| F344/NSlc rats | 2 mg/ 4 mg N-methylnitrourea x 3 per wk (3 wks), plain water (1), 17 ppm Lyc (2), diluted tomato juice with 17 ppm Lyc (3), diluted tomato juice with 3.4 ppm Lyc (4) | (3) ↓colon cancer incidence, but not in (2) | [78] | |
| Hepatocellular carcinoma (HCC)LEC rats | Diet containing 0.005% Lyc (1), 1% TJ-9: crude extracts of 7 herbs (2), control (3) administered from 6 wks age to 76 wks age | ↔number, mean area and % area GST-P-+ focal lesions (liver, HCC); Note: TJ-9 had higher number of GST-P-+ lesion in HCC), AFP, cumulative survival rates ↓iron conc. in liver | [81] | |
| N-methyl-N’-nitrosoguanidine (MNNG) and saturated NaCl (S-NaCl) induced Male Wistar rats | N-methyl-N’-nitrosoguanidine (MNNG) + saturated NaCl (1), MNNG + S-NaCl + Lyc (2), Lyc (3), Control (4) | ↓gastric carcinomas ↑GSH, GPx, GST, GR | [73] | |
| MNNG + S-NaCl (1), MNNG + S-NaCl + Sallylcysteine (SAC) (2), MNNG + S-NaCl + Lyc (3), MNNG + S-NaCl + SAC + Lyc (4), chemoprevention agents (5–7), Control (8) | ↔GSH (stomach, erythrocytes), GPx (liver, erythrocytes), GPx activities (stomach), Bax, Bim ↑GSH (liver), GPx (stomach), GSH activities, GPx activities (liver, erythrocytes), caspase-8 ↓tumor burden, Bcl-2 | [26] | ||
| Resistant hepatocyte (RH) model of hepatocarcinogenesis Wistar rats | 70 mg/kg BW lutein (1), Lyc (2), Control (3) | ↑liver carotenoid conc. ↔incidence, total number, multiplicity of hepatocyte nodules ↓No., size, area of GST+ preneoplastic lesions, hepatic DNA strand breakage | [71] | |
| Colon carcinogenesis Sprague Dawley rat model | Induction by azoxymethane, followed by treatment with diallylsulfide (1), Lyc (2), theaflavin (3) | ↓aberrant crypt foci, preneoplastic lesion, COX-2, iNOS | [60] | |
| Nude mice | Supplementation 2× per wk (12 wks), with 1, 20 mg/kg BW Lyc, 20 mg/kg BW β-carotene; starting wk 2, injection with SK-Hep-1 cells via tail vein | ↓MMP-2, VEGF, tumor metastasis, mean no. of tumors, tumor cross-sectional area, PCNA, MMP-9 ↑nm23-H1 | [22] | |
| Hepatocarcinogenesis in rat model | Injected with diethylnitrosamine (DEN) and fed with control diet or high fat diet (HFD) with or without Lyc or tomato extract | (HFD + Lyc) ↓no. of GST+ hepatic foci, PCNA, cyclin D1, ERKs, NF-κB ↔TNF-α, IL-1β, IL-12, CYP2E1 ↑HO-1, NRF2 | [72] | |
| N-methyl-N′-nitrosoguanidine (MNNG) gastric cancer rat model | Control (1), 200 mg/kg BW MNNG + saturated NaCl (2), 200 mg/kg BW MNNG + saturated NaCl + 50 mg/kg BW Lyc (3) 200 mg/kg BW MNNG + saturated NaCl + 100 mg/kg BW Lyc (4) 200 mg/kg BW MNNG + saturated NaCl + 150 mg/kg BW Lyc (5) | ↑SOD, CAT, GSH-Px, IL-2, IL-4, IL-10, TNF-α, IgG, IgA, IgM ↓MDA, IL-6 | [61] | |
| BCO2-knockout and wild-type male mice | Lyc supplementation (100 mg/kg diet, 24 wks), induced by high fat diet | (BCO2-KO) ↑hepatic Lyc, miR-199a/b, miR214 ↓hepatocellular carcinoma incidence, multiplicity, ER(UPR), Met mRNA, β-catenin, mTORC1 (Wild type) ↓NF-κB p65, STAT3, IL-6, inflammatory foci | [62] | |
| (In vitro) OV-MZ-6 cells | Treatment with 2, 5 µM Lyc | ↓ITGA5, pERK 1/2 ↔ITGB1, tERK, vimentin | [75] | |
| (In vivo) Ovarian cancer-bearing mice | Prevention Gp: Placebo (1), Lyc (2) Treatment Gp: Placebo (1), Lyc (2), Lyc + Taxol (3), Taxol + Platin (4), Platin (5), Lyc + Taxol + Platin (6) Concentration of Lyc: 0.75 mg/mL | (Lyc Prevention Gp) ↓metastatic load, Ki67, ITGA5B1, ITGA5, ILK, ITGB1, FAK, MMP-9, serum and ascites CA125, EMT markers in metastatic tissue (Note: MMP-9 restricted to metastatic tissue, not tumor tissue) ↔tumor load, serum and ascites MMP-9 (Lyc Treatment Gp) ↓tumor load, Ki67, ITGA5, ITGA5B1, ascites CA125 ↑MMP-9 ↔ILK, ITGB1, FAK, serum and ascites MMP-9, serum CA125 | ||
| Laying hens | Control (1), 200 mg/kg per kg diet Lyc (2), 400 mg/kg per kg diet Lyc (3) | ↓incidence, no. and size ovarian tumor, rate of adenocarcinoma, MDA, NF-κB, STAT3 ↑NFE2, HO-1 | [76] | |
| (In vitro) Lewis lung carcinoma (LLC) cells | Control (1), Lyc: 10 µM (2), Lyc: 20 µM (3), Lyc: 40 µM (4) | ↑(mRNA) IFNβ, IFNγ, IRF1, IRF7, CXCL9, CXCL10, pJAK, pSTAT3 ↓(mRNA) DMNT3a, methylation levels of promoters (IRF1, IRF7), PD-1 due to IFNγ, pAkt ↔(mRNA) DNMT1, DNMT3b | [69] | |
| (In vivo) C57BL/6 mice | Control (1), Anti PD-1, 6 mg/kg (2), Lyc, 40 mg/kg (3), Anti-PD-1 + Lyc (4); intraperitoneal, 3 days, 4 times | ↓tumor volume, weight, IL-4, IL-10, (mRNA) DMNT3a, methylation levels of promoters (IRF1, IRF7) ↑IL-2, IFNγ, CD4+:CD8+, % IFNγ+/CD8+ T cell, % perforin+/CD8+ T cell, % granzyme B+/CD8+ T cell, (mRNA) IFNβ, IFNγ, IRF1, IRF7, CXCL9, CXCL10 ↔(mRNA) IRF3, IRF8, DNMT1, DNMT3b | ||
| CD-1 mice in AOM-DSS model | Normal (1), AOM+DSS control (2), (Bifidobacterium longum) BF + AOM + DSS (3), BF + Lyc 20 mg/kg + AOM + DSS (4), BF + Lyc 50 mg/kg + AOM + DSS (5), Lyc 20 mg/kg + AOM + DSS (6), Lyc 50 mg/kg + AOM + DSS (7), Metformin + AOM + DSS (8) | ↑positive rates of IGF-1, IGF-2 (high dose), IGF-1R, IGF2BP1, IGFBP2 (low dose), IGFBP3 (high dose), lymphocyte infiltration ↓inflammation incidence, positive rates of IGF-2 (low dose), IGFBP2 (high dose), IGFBP3 (low dose) ↔No. of tumors, adenocarcinomas incidence Note: presence of focal necrosis | [79] | |
| Lycopene-Enriched Tomato Oleoresin (LETO) | Rat mammary tumor model | Induced with 7, 12-dimethyl-benz[a]anthracene (DMBA) 2 wks, followed by injection of 10mg/kg LETO (1), β-carotene (2), control (3) twice per wk, 16 wks | ↑plasma, hepatic Lyc ↓tumors, tumor area | [68] |
| Supplementation with 250 ppm Lyc (1), 500 ppm (2), 250 ppm lycopene-rich tomato carotenoid oleoresin (TCO) (3), 500 ppm TCO (4), control (5) followed by initiation with N-methylnitrosourea (NMU) (7 days) 18 wks experimentation | ↔tumor incidence, latency, multiplicity, volume, total tumors per group Note: supplementation with TCO ↑serum Lyc conc. > supplementation with pure Lyc | [82] |
| Compound | Subject | Experiment Design | Outcome | Ref |
|---|---|---|---|---|
| Lycopene (Lyc) | 47,894 human subjects initially free of diagnosed cancer | Validated semiquantitative food-frequency questionnaire | ↓risk of non-stage A1 prostate cancer | [83] |
| 26 male patients with prostate cancer, 14 stage T1, 12 stage T2 | Control (1), 15 mg Lyc (2) | ↓plasma prostate-specific antigen (PSA) ↑connexin 43 ↔Bcl-2, Bax Note: sample size is relatively small | [86] | |
| 47,365 participants | Dietary questionnaires | ↓risk of prostate cancer Note: moderate association | [84] | |
| 32 patients with localized prostate adenocarcinoma | Randomized placebo-controlled study: 30 mg Lyc/day | ↑serum and prostate Lyc conc., apoptotic index (hyperplastic and neoplastic cells) ↓serum PSA, leukocyte 8OHdG | [87] | |
| 58,279 men aged 55–69 yrs: 642 prostate cancer cases | Cohort study, 6.3 yrs follow-up, semi-quantitative food-frequency questionnaire | ↔risk of prostate cancer | [96] | |
| 69 men with favourable risk prostate cancer | 3-month randomized, double blinded clinical trial: 30 mg/day Lyc (1), 3 g/day fish oil (2), placebo (3) | ↔IGF-1, COX-2 | [97] | |
| 11 cohort studies, 6 nested case–control studies | Meta-analysis | OR < 1 (high tomato intake and incidence of prostate cancer) Modest effect in prevention of prostate cancer | [89] | |
| 26 studies with 17,517 cases of prostate cancer, from 563,299 participants | Meta-analysis | ↓risk of prostate cancer (Lyc: 9–21 mg/day; plasma Lyc: 2.17–85 µg/dL) | [90] | |
| 18 prospective cohort studies in 2012 | Pooled analysis (interval collapsing method) | Protective effect towards ER−/PR+ or ER−/PR− breast cancer | [91] | |
| Plasma Lycopene | 25,802 persons: 103 men with prostate cancer, 103 men as control | Analysis of serum | ↔risk of prostate cancer | [94] |
| 209 prostate cancer cases, 228 control, Black and white men in US (40–79 yrs old) | Analysis of serum carotenoids | ↔risk of prostate cancer, only useful particularly for aggressive disease Note: insignificant inverse association (serum Lyc and prostate cancer) | [95] | |
| 450 incident prostate cancer cases | Case-control study nested within prospective Health Professionals Follow-up Study | ↓risk of prostate cancer Note: restricted to older participants, without family history | [85] | |
| 521 women with breast cancer | Analysis of serum using HPLC | ↓risk of breast cancer among premenopausal women and all ER/PR subtypes | [92] | |
| 17 prospective studies with 3603 cases, 458,434 participants | Meta-analysis | Nonlinear dose-dependent (lung cancer and plasma Lyc) Note: stronger inverse association at low plasma Lyc conc.) | [93] | |
| Lycopene-rich tomato | 79 prostate cancer patients | Nutritional intervention: tomato products with 30 mg Lyc (1), tomato products + selenium, omega-3 fatty acids, soy isoflavones, grape/pomegranate juice and green/black tea (2), Control (3) | ↓PSA level | [88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puah, B.-P.; Jalil, J.; Attiq, A.; Kamisah, Y. New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene. Molecules 2021, 26, 3888. https://doi.org/10.3390/molecules26133888
Puah B-P, Jalil J, Attiq A, Kamisah Y. New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene. Molecules. 2021; 26(13):3888. https://doi.org/10.3390/molecules26133888
Chicago/Turabian StylePuah, Boon-Peng, Juriyati Jalil, Ali Attiq, and Yusof Kamisah. 2021. "New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene" Molecules 26, no. 13: 3888. https://doi.org/10.3390/molecules26133888
APA StylePuah, B. -P., Jalil, J., Attiq, A., & Kamisah, Y. (2021). New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene. Molecules, 26(13), 3888. https://doi.org/10.3390/molecules26133888