
Cytotoxicity of a Lipid-Rich Extract from Native Mexican Avocado Seed (Persea americana var. drymifolia) on Canine Osteosarcoma D-17 Cells and Synergistic Activity with Cytostatic Drugs

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. LEAS Is Cytotoxic for D-17 Cells
2.2. LEAS Does Not Affect the Cell Membrane of D-17 Cells
2.3. LEAS Cytotoxicity on D-17 Cells Is Related to Apoptosis Induction
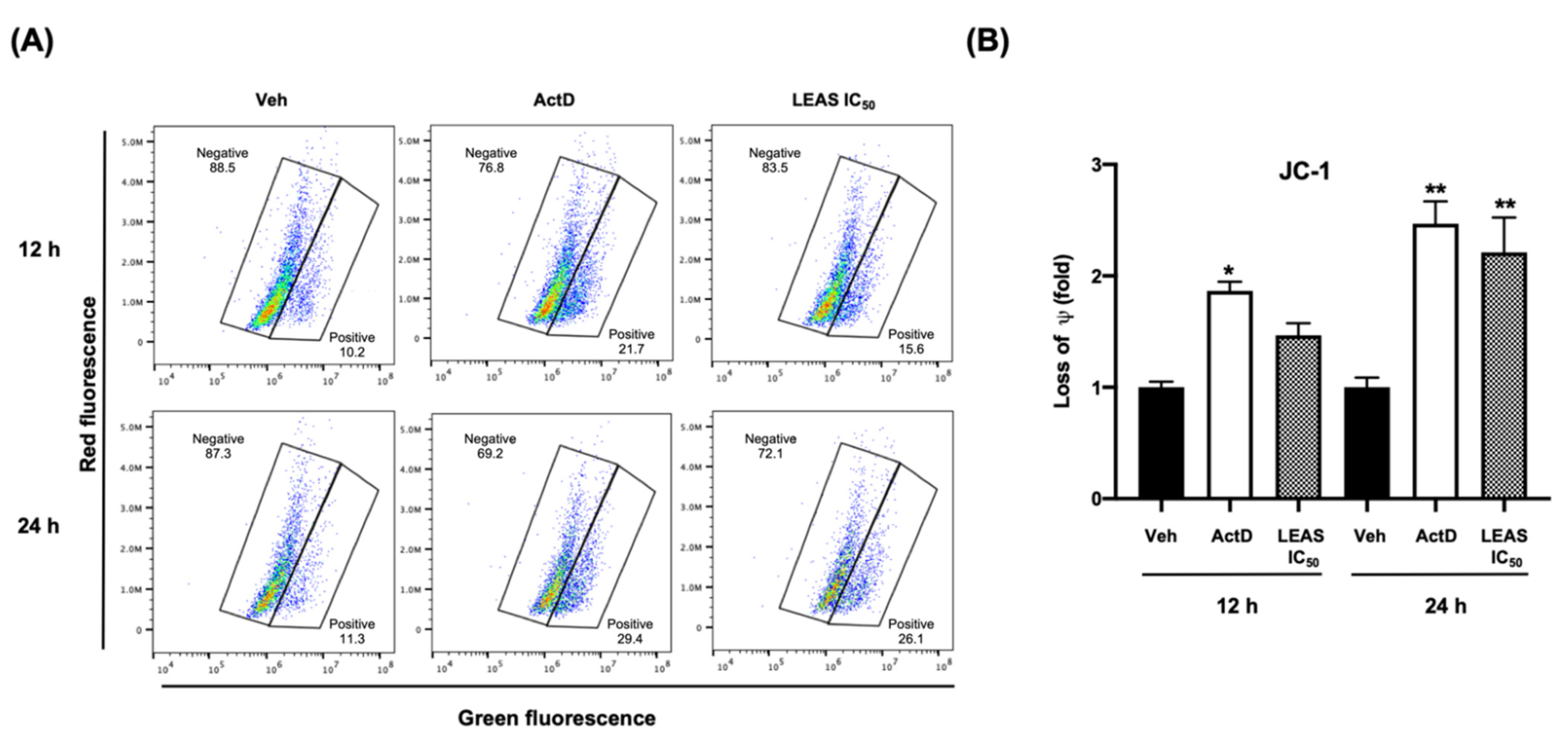
2.4. LEAS Cytotoxicity Is Associated with Loss of Mitochondrial Membrane Potential and ROS Production in D-17 Cells
2.5. LEAS Induces Cell Cycle Arrest in D-17 Cells
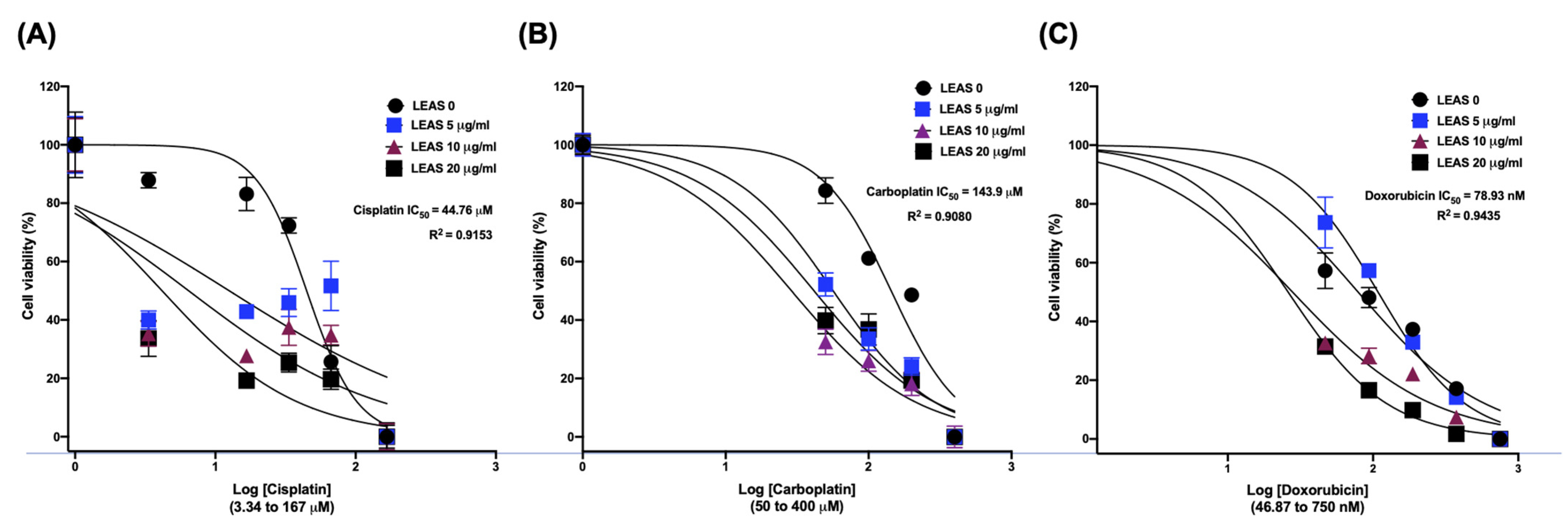
2.6. LEAS Shows a Synergistic Effect with Cytotoxic Drugs in D-17 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents, Cell Lines, and Cell Culture
4.2. Lipid-Rich Extract from Avocado Seed (LEAS)
4.3. MTT Viability Assay
4.4. Calcium Efflux Testing
4.5. Measurement of the Membrane Potential
4.6. Assessment of Mitochondrial Membrane Potential (ΔΨm)
4.7. Determination of Reactive Oxygen Species (ROS)
4.8. Apoptosis Analysis
4.9. Cell Cycle Analysis
4.10. Drug Combination Studies
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Adams, V.J.; Evans, K.M.; Sampson, J.; Wood, J.L. Methods and mortality results of a health survey of purebred dogs in the UK. J. Small Anim. Pract. 2010, 51, 512–524. [Google Scholar] [CrossRef]
- Dobson, J.M. Breed-Predispositions to Cancer in Pedigree Dogs. ISRN Vet. Sci. 2013, 2013, 941275. [Google Scholar] [CrossRef]
- Vail, D.M.; MacEwen, E.G. Spontaneously Occurring Tumors of Companion Animals as Models for Human Cancer. Cancer Investig. 2000, 18, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Mueller, F.; Fuchs, B.; Kaser-Hotz, B. Comparative biology of human and canine osteosarcoma. Anticancer Res. 2007, 27, 155–164. [Google Scholar] [PubMed]
- Schiffman, J.D.; Breen, M. Comparative oncology: What dogs and other species can teach us about humans with cancer. Philos. Trans. R. Soc. B 2015, 370, 20140231. [Google Scholar] [CrossRef] [PubMed]
- Egenvall, A.; Nødtvedt, A.; von Euler, H. Bone tumors in a population of 400,000 insured Swedish dogs up to 10 y of age: Incidence and survival. Can. J. Vet. Res. 2007, 71, 292–299. [Google Scholar]
- Simpson, S.; Dunning, M.; de Brot, S.; Alibhai, A.; Bailey, C.; Woodcock, C.L.; Mestas, M.; Akhtar, S.; Jeyapalan, J.N.; Lothion-Roy, J.; et al. Molecular Characterisation of Canine Osteosarcoma in High Risk Breeds. Cancers 2020, 12, 2405. [Google Scholar] [CrossRef]
- Simpson, S.; Dunning, M.D.; de Brot, S.; Grau-Roma, L.; Mongan, N.P.; Rutland, C.S. Comparative review of human and canine osteosarcoma: Morphology, epidemiology, prognosis, treatment and genetics. Acta Vet. Scand. 2017, 59, 1–11. [Google Scholar] [CrossRef]
- Selvarajah, G.T.; Kirpensteijn, J. Prognostic and predictive biomarkers of canine osteosarcoma. Vet. J. 2010, 185, 28–35. [Google Scholar] [CrossRef]
- Szewczyk, M.; Lechowski, R.; Zabielska, K. What do we know about canine osteosarcoma treatment?—Review. Vet. Res. Commun. 2014, 39, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Ostrander, E.A.; Dreger, D.L.; Evans, J.M. Canine Cancer Genomics: Lessons for Canine and Human Health. Annu. Rev. Anim. Biosci. 2019, 7, 449–472. [Google Scholar] [CrossRef]
- Pasic, I.; Lipton, J.H. Current approach to the treatment of chronic myeloid leukaemia. Leuk. Res. 2017, 55, 65–78. [Google Scholar] [CrossRef]
- Wang, H.; Khor, T.O.; Shu, L.; Su, Z.-Y.; Fuentes, F.; Lee, J.H.; Kong, A.-N.T. Plants vs. Cancer: A Review on Natural Phytochemicals in Preventing and Treating Cancers and Their Druggability. Anti-Cancer Agents Med. Chem. 2012, 12, 1281–1305. [Google Scholar] [CrossRef]
- Isani, G.; Bertocchi, M.; Andreani, G.; Farruggia, G.; Cappadone, C.; Salaroli, R.; Forni, M.; Bernardini, C. Cytotoxic Effects of Artemisia annua L. and Pure Artemisinin on the D-17 Canine Osteosarcoma Cell Line. Oxid. Med. Cell. Longev. 2019, 2019, 1615758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, C.B.; Bayle, J.; Biourge, V.; Wakshlag, J.J. Cellular effects of a turmeric root and rosemary leaf extract on canine neoplastic cell lines. BMC Vet. Res. 2017, 13, 388. [Google Scholar] [CrossRef] [Green Version]
- Dabas, D.; Shegog, R.; Ziegler, G.; Lambert, J. Avocado (Persea americana) Seed as a Source of Bioactive Phytochemicals. Curr. Pharm. Des. 2013, 19, 6133–6140. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, S.M.; Han, C.; Pan, L.; Kinghorn, A.D.; Ding, H. Aliphatic acetogenin constituents of avocado fruits inhibit human oral cancer cell proliferation by targeting the EGFR/RAS/RAF/MEK/ERK1/2 pathway. Biochem. Biophys. Res. Commun. 2011, 409, 465–469. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.A.; Angka, L.; Rota, S.-G.; Hanlon, T.; Mitchell, A.; Hurren, R.; Wang, X.M.; Gronda, M.; Boyaci, E.; Bojko, B.; et al. Targeting Mitochondria with Avocatin B Induces Selective Leukemia Cell Death. Cancer Res. 2015, 75, 2478–2488. [Google Scholar] [CrossRef] [Green Version]
- Butt, A.J.; Roberts, C.G.; Seawright, A.A.; Oelrichs, P.B.; MacLeod, J.K.; Liaw, T.Y.; Kavallaris, M.; Somers-Edgar, T.J.; Lehrbach, G.M.; Watts, C.K.; et al. A novel plant toxin, persin, with in vivo activity in the mammary gland, induces Bim-dependent apoptosis in human breast cancer cells. Mol. Cancer Ther. 2006, 5, 2300–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Han, C.; Guo, D.; Chin, Y.-W.; Ding, Y.; Kinghorn, A.D.; D’Ambrosio, S.M. Selective Induction of Apoptosis of Human Oral Cancer Cell Lines by Avocado Extracts via a ROS-Mediated Mechanism. Nutr. Cancer 2009, 61, 348–356. [Google Scholar] [CrossRef]
- Rosenblat, G.; Meretski, S.; Segal, J.; Tarshis, M.; Schroeder, A.; Zanin-Zhorov, A.; Lion, G.; Ingber, A.; Hochberg, M. Polyhydroxylated fatty alcohols derived from avocado suppress inflammatory response and provide non-sunscreen protection against UV-induced damage in skin cells. Arch. Dermatol. Res. 2010, 303, 239–246. [Google Scholar] [CrossRef]
- Lara-Márquez, M.; Báez-Magaña, M.; Raymundo-Ramos, C.; Spagnuolo, P.A.; Macías-Rodríguez, L.; Salgado-Garciglia, R.; Ochoa-Zarzosa, A.; López-Meza, J.E. Lipid-rich extract from Mexican avocado (Persea americana var. drymifolia) induces apoptosis and modulates the inflammatory response in Caco-2 human colon cancer cells. J. Funct. Foods 2020, 64, 103658. [Google Scholar] [CrossRef]
- Ochoa-Zarzosa, A.; Báez-Magaña, M.; Guzmán-Rodríguez, J.J.; Flores-Alvarez, L.J.; Lara-Márquez, M.; Zavala-Guerrero, B.; Salgado-Garciglia, R.; López-Gómez, R.; López-Meza, J.E. Bioactive Molecules from Native Mexican Avocado Fruit (Persea americana var. drymifolia): A Review. Plant Foods Hum. Nutr. 2021, 76, 133–142. [Google Scholar] [CrossRef]
- Báez-Magaña, M.; Ochoa-Zarzosa, A.; Alva-Murillo, N.; Salgado-Garciglia, R.; López-Meza, J.E. Lipid-rich extract from Mexican avocado seed (Persea americana var. drymifolia) reduces Staphylococcus aureus internalization and regulates innate immune response in bovine mammary epithelial cells. J. Immunol. Res. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Albarrán, F.; Salgado-Garciglia, R.; Molina-Torres, J.; López-Gómez, R. Oleosome Oil Storage in the Mesocarp of Two Avocado Varieties. J. Oleo Sci. 2019, 68, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.-Y.; Arteaga, J.R.; Zhang, Q.; Huerta, S.; Go, V.L.W.; Heber, D. Inhibition of prostate cancer cell growth by an avocado extract: Role of lipid-soluble bioactive substances. J. Nutr. Biochem. 2005, 16, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Helmerick, E.C.; Loftus, J.; Wakshlag, J.J. The effects of baicalein on canine osteosarcoma cell proliferation and death. Vet. Comp. Oncol. 2012, 12, 299–309. [Google Scholar] [CrossRef]
- Alkhalaf, M.I.; Alansari, W.S.; Ibrahim, E.A.; Elhalwagy, M.E. Anti-oxidant, anti-inflammatory and anti-cancer activities of avocado (Persea americana) fruit and seed extract. J. King Saud Univ. 2019, 31, 1358–1362. [Google Scholar] [CrossRef]
- Widiyastuti, Y.; Pratiwi, R.; Riyanto, S.; Wahyuono, S. Cytotoxic activity and apoptosis induction of avocado Persea americana Mill. seed extract on MCF-7 cancer cell line. Indones. J. Biotechnol. 2018, 23, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Poradowski, D.; Obmińska-Mrukowicz, B. Effect of selected nonsteroidal anti-inflammatory drugs on the viability of canine osteosarcoma cells of the D-17 line: In vitro studies. J. Vet. Res. 2019, 63, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Park, S.; Bazer, F.W.; Lim, W.; Song, G. Myricetin treatment induces apoptosis in canine osteosarcoma cells by inducing DNA fragmentation, disrupting redox homeostasis, and mediating loss of mitochondrial membrane potential. J. Cell. Physiol. 2018, 233, 7457–7466. [Google Scholar] [CrossRef]
- Hosoya, K.; Murahari, S.; Laio, A.; London, C.A.; Couto, C.G.; Kisseberth, W.C. Biological activity of dihydroartemisinin in canine osteosarcoma cell lines. Am. J. Vet. Res. 2008, 69, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Chin, Y.-W.; Kinghorn, A.D.; D’Ambrosio, S.M. Chemopreventive characteristics of avocado fruit. Semin. Cancer Biol. 2007, 17, 386–394. [Google Scholar] [CrossRef]
- Zhao, J.; Li, R.; Pawlak, A.; Henklewska, M.; Sysak, A.; Wen, L.; Yi, J.-E.; Obmińska-Mrukowicz, B. Antitumor Activity of Betulinic Acid and Betulin in Canine Cancer Cell Lines. In Vivo 2018, 32, 1081–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, I.; Melo-Alvim, C.; Lopes-Brás, R.; Esperança-Martins, M.; Costa, L. Osteosarcoma Pathogenesis Leads the Way to New Target Treatments. Int. J. Mol. Sci. 2021, 22, 813. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Rodríguez, J.J.; López-Gómez, R.; Salgado-Garciglia, R.; Ochoa-Zarzosa, A.; López-Meza, J.E. The defensin from avocado (Persea americana var. drymifolia) PaDef induces apoptosis in the human breast cancer cell line MCF-7. Biomed. Pharmacother. 2016, 82, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Aapro, M.; Alberts, D.; Salmon, S. Interactions of human leukocyte interferon with vinca alkaloids and other chemotherapeutic agents against human tumors in clonogenic assay. Cancer Chemother. Pharmacol. 1983, 10, 161–166. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padilla-Arellanes, S.; Salgado-Garciglia, R.; Báez-Magaña, M.; Ochoa-Zarzosa, A.; López-Meza, J.E. Cytotoxicity of a Lipid-Rich Extract from Native Mexican Avocado Seed (Persea americana var. drymifolia) on Canine Osteosarcoma D-17 Cells and Synergistic Activity with Cytostatic Drugs. Molecules 2021, 26, 4178. https://doi.org/10.3390/molecules26144178
Padilla-Arellanes S, Salgado-Garciglia R, Báez-Magaña M, Ochoa-Zarzosa A, López-Meza JE. Cytotoxicity of a Lipid-Rich Extract from Native Mexican Avocado Seed (Persea americana var. drymifolia) on Canine Osteosarcoma D-17 Cells and Synergistic Activity with Cytostatic Drugs. Molecules. 2021; 26(14):4178. https://doi.org/10.3390/molecules26144178
Chicago/Turabian StylePadilla-Arellanes, Salvador, Rafael Salgado-Garciglia, Marisol Báez-Magaña, Alejandra Ochoa-Zarzosa, and Joel Edmundo López-Meza. 2021. "Cytotoxicity of a Lipid-Rich Extract from Native Mexican Avocado Seed (Persea americana var. drymifolia) on Canine Osteosarcoma D-17 Cells and Synergistic Activity with Cytostatic Drugs" Molecules 26, no. 14: 4178. https://doi.org/10.3390/molecules26144178
APA StylePadilla-Arellanes, S., Salgado-Garciglia, R., Báez-Magaña, M., Ochoa-Zarzosa, A., & López-Meza, J. E. (2021). Cytotoxicity of a Lipid-Rich Extract from Native Mexican Avocado Seed (Persea americana var. drymifolia) on Canine Osteosarcoma D-17 Cells and Synergistic Activity with Cytostatic Drugs. Molecules, 26(14), 4178. https://doi.org/10.3390/molecules26144178