
Protective Effect of Potentilla glabra in UVB-Induced Photoaging Process



Abstract
:1. Introduction
2. Results
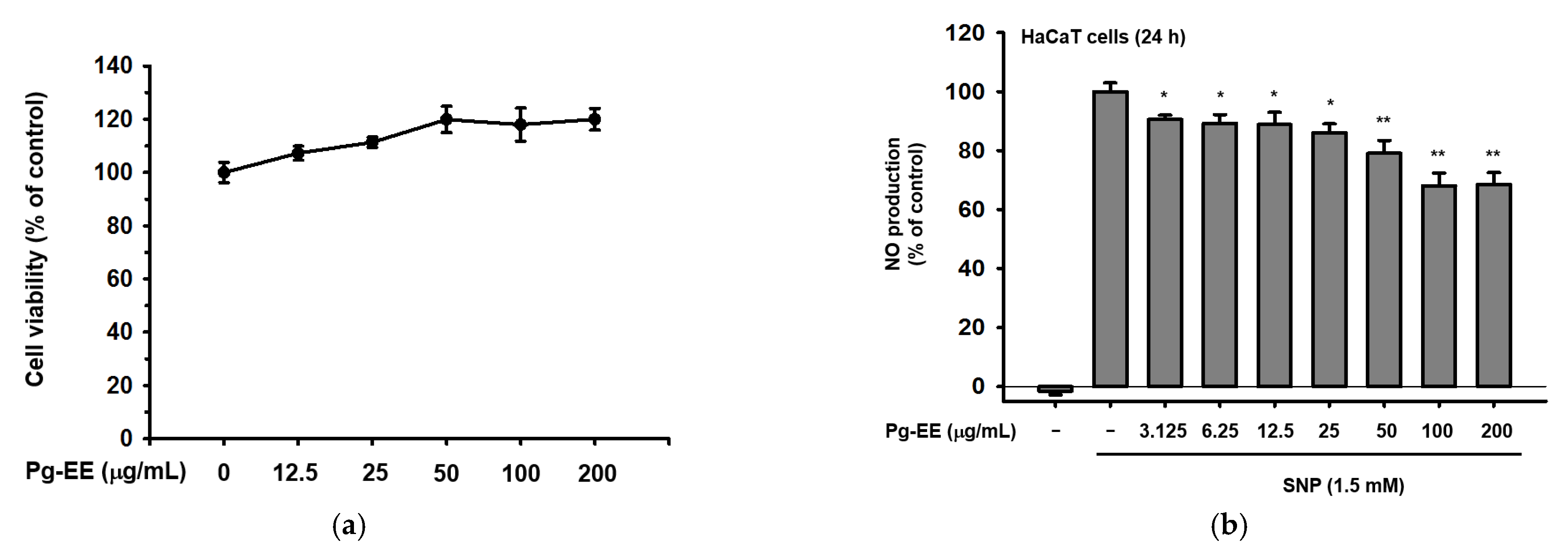
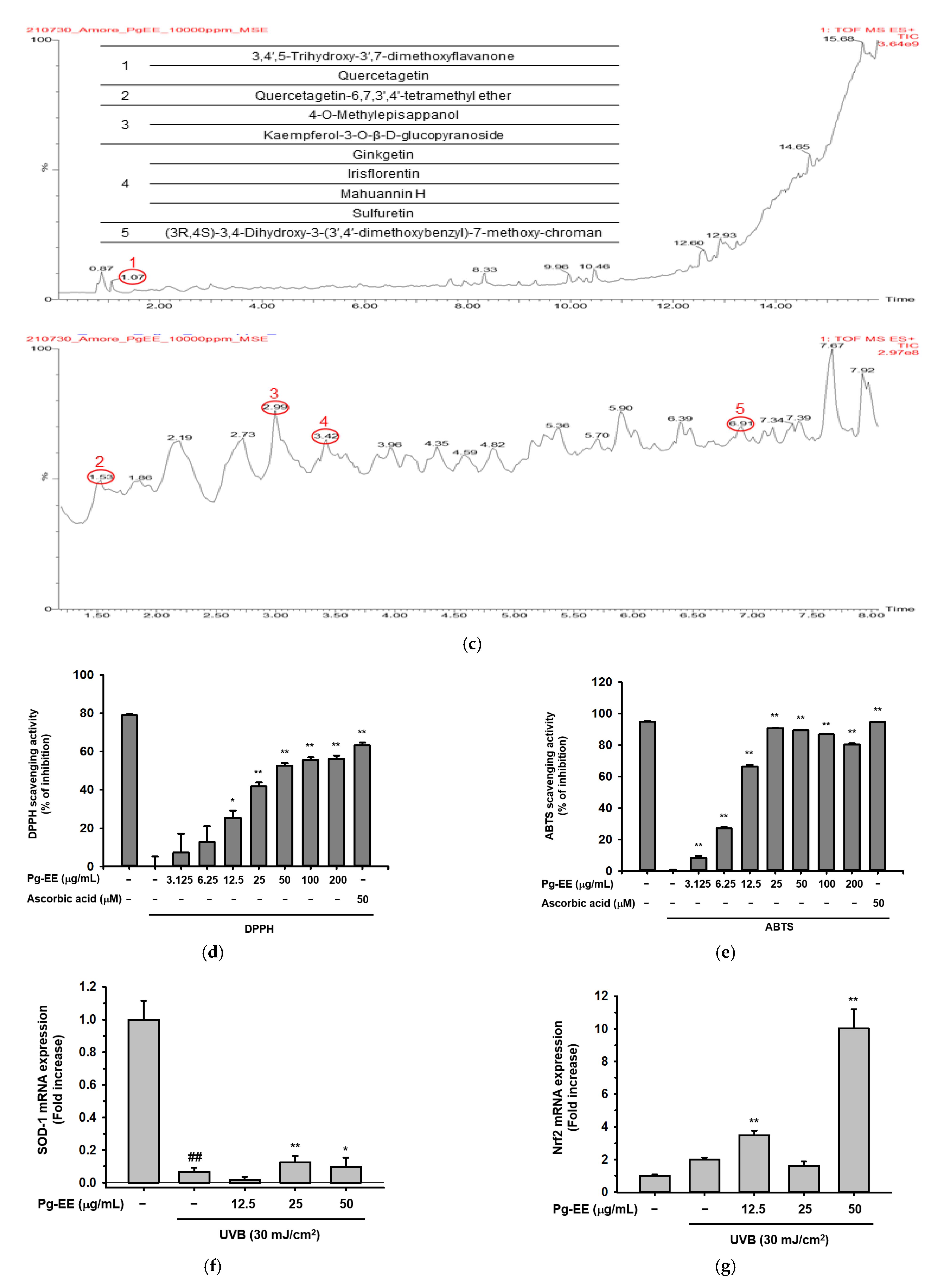
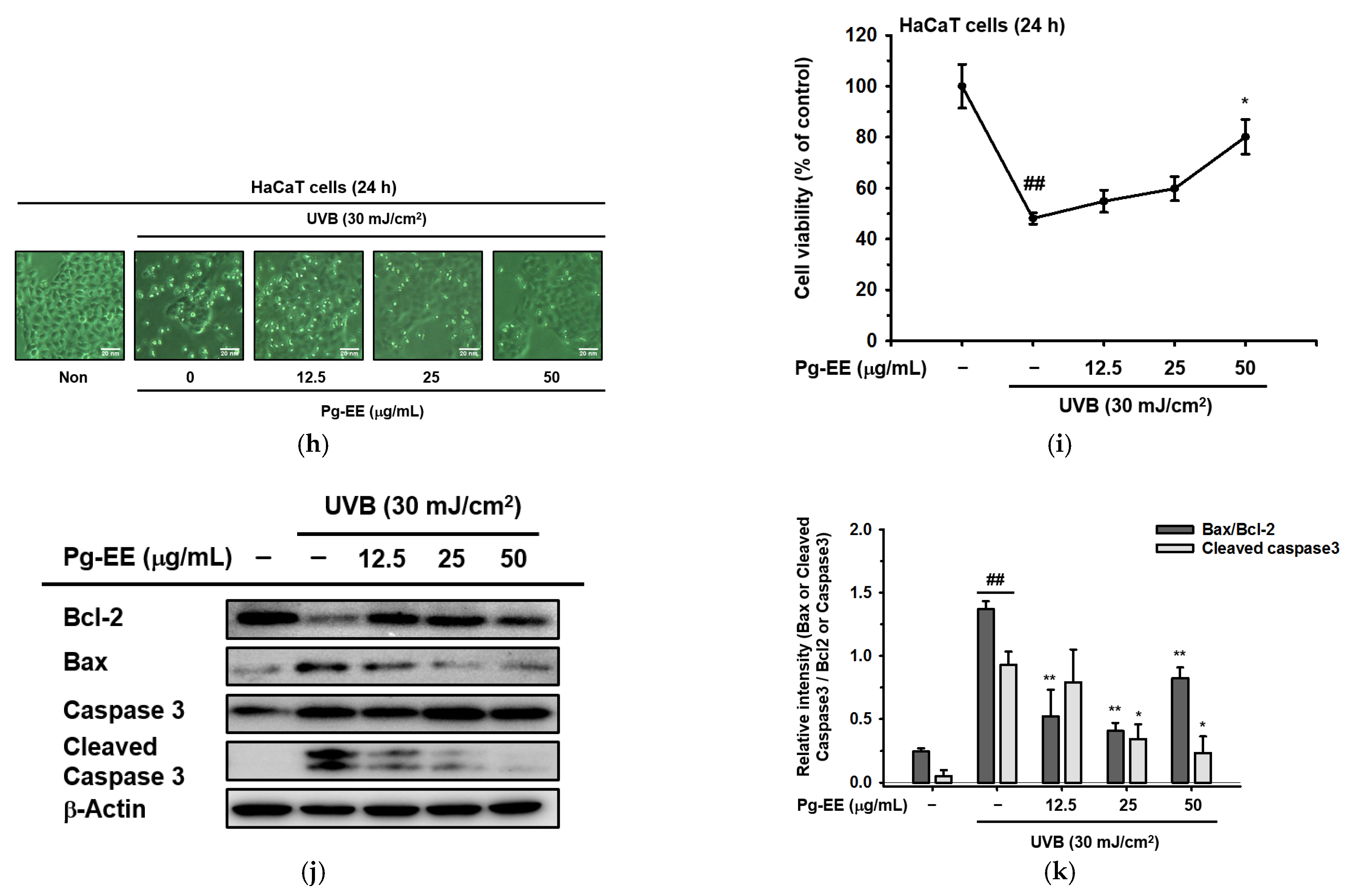
2.1. Effects of Pg-EE on Cell Viability and Skin-Protective Activities
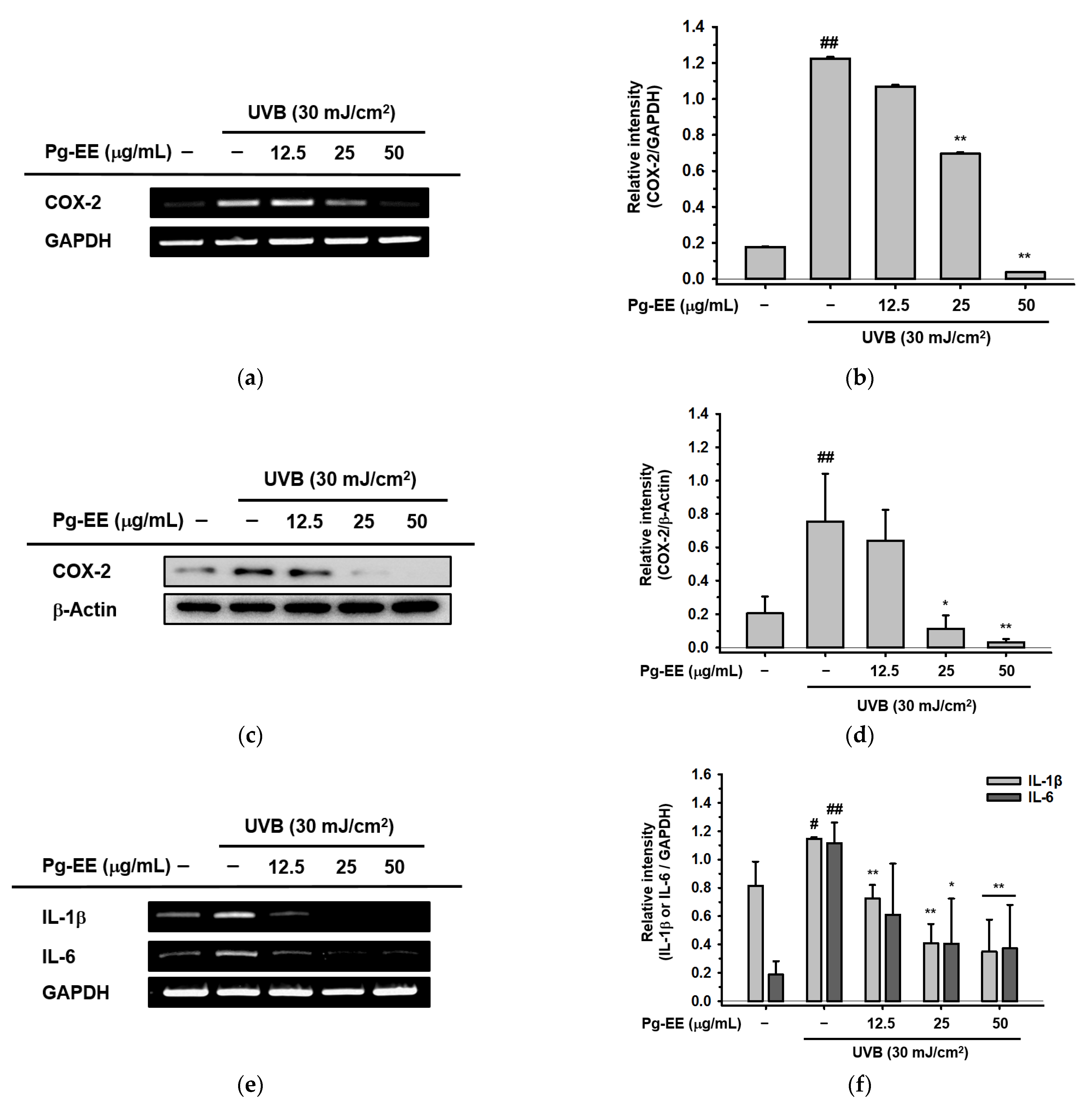
2.2. Anti-Inflammatory Abilities of Pg-EE in Keratinocytes under UVB Irradiation
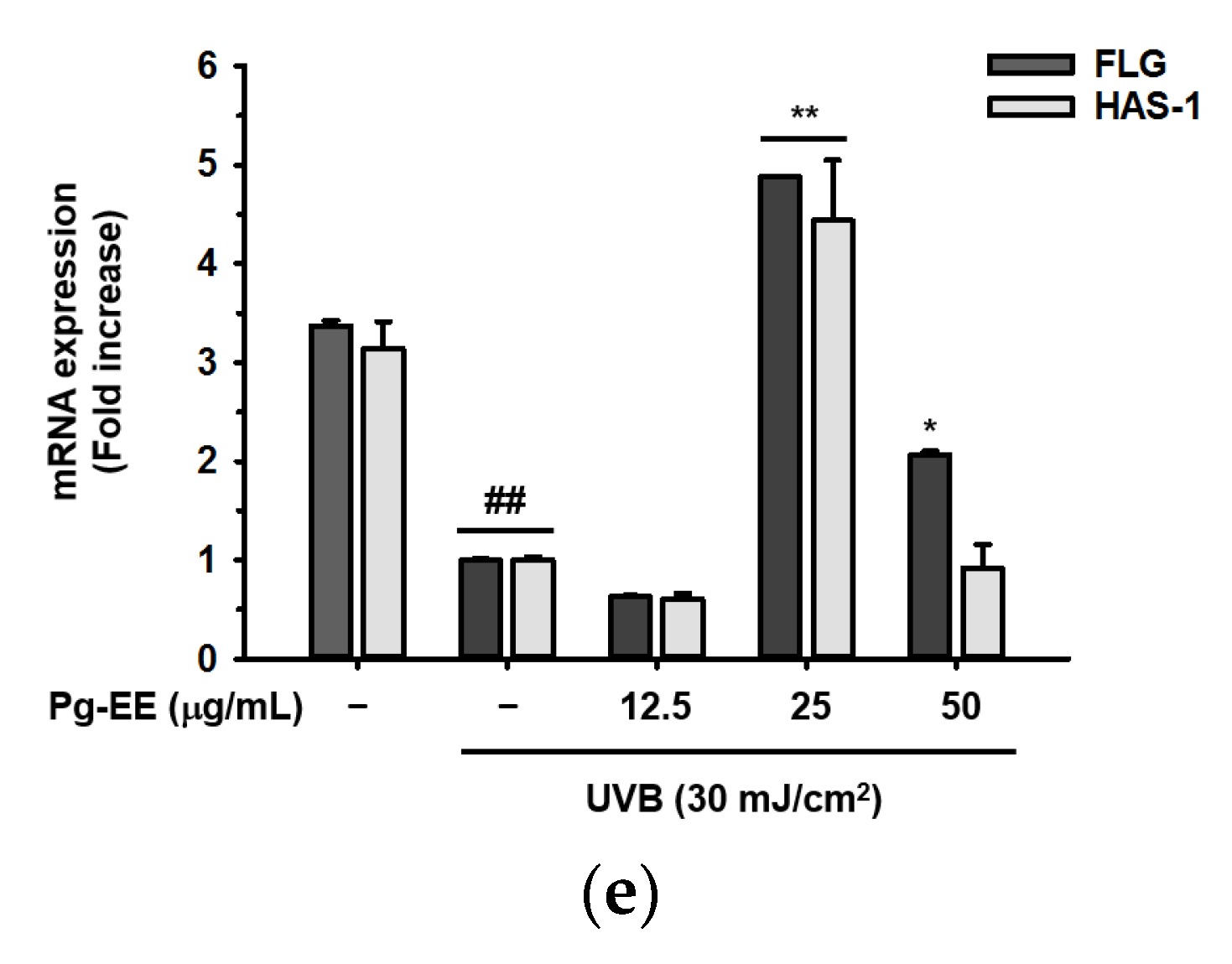
2.3. Effect of Pg-EE on Skin Moisture Protection Activity
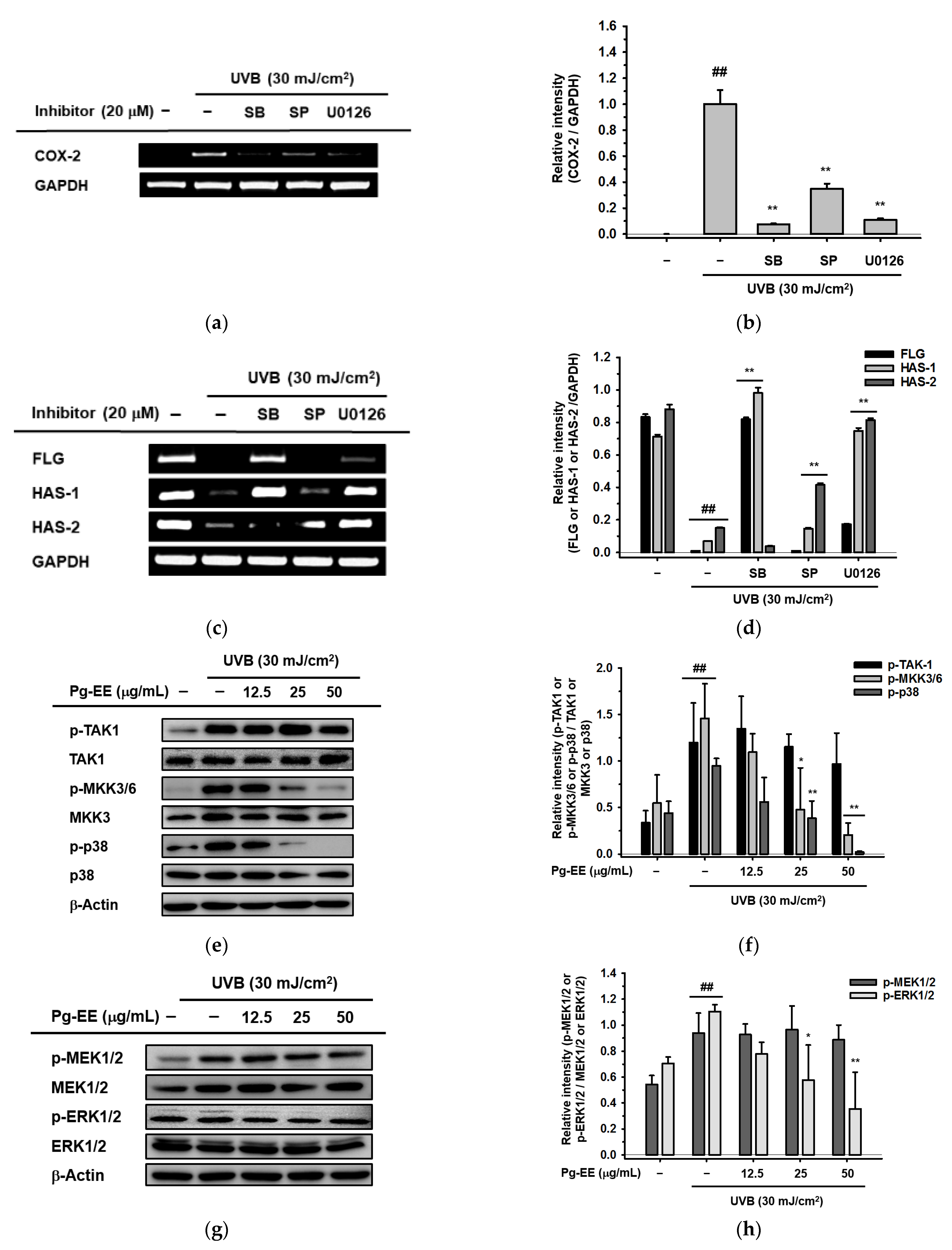
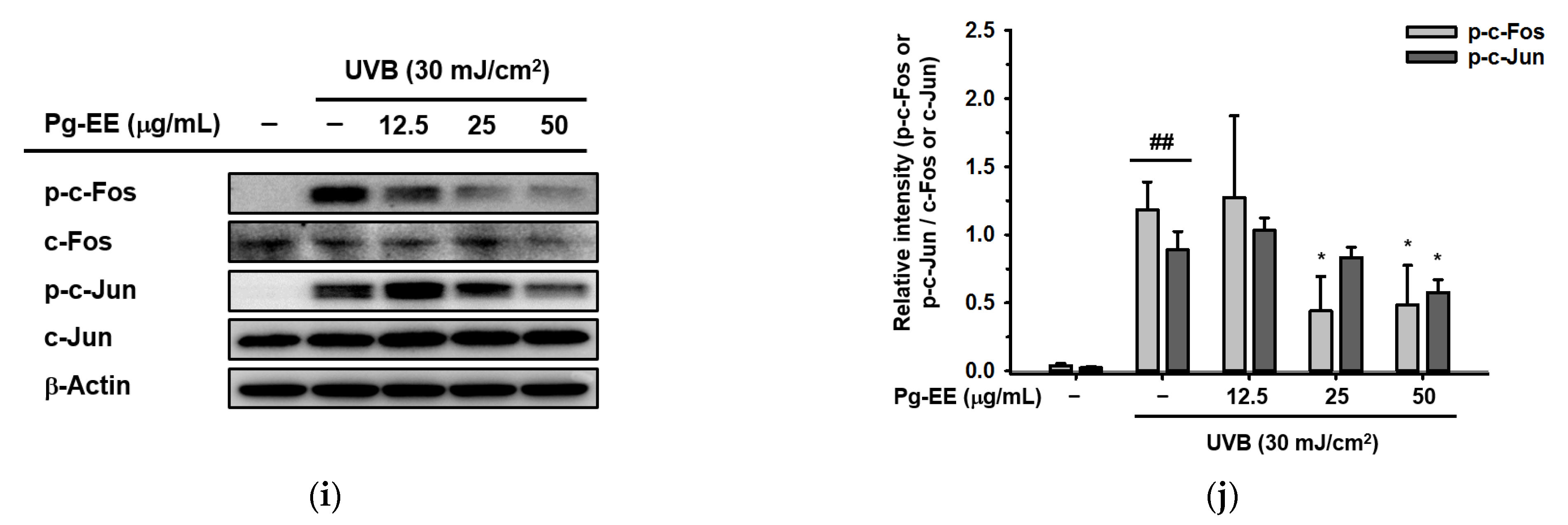
2.4. Pg-EE Shows Anti-Inflammatory and Moisture Protective Abilities via AP-1 Pathway under UVB
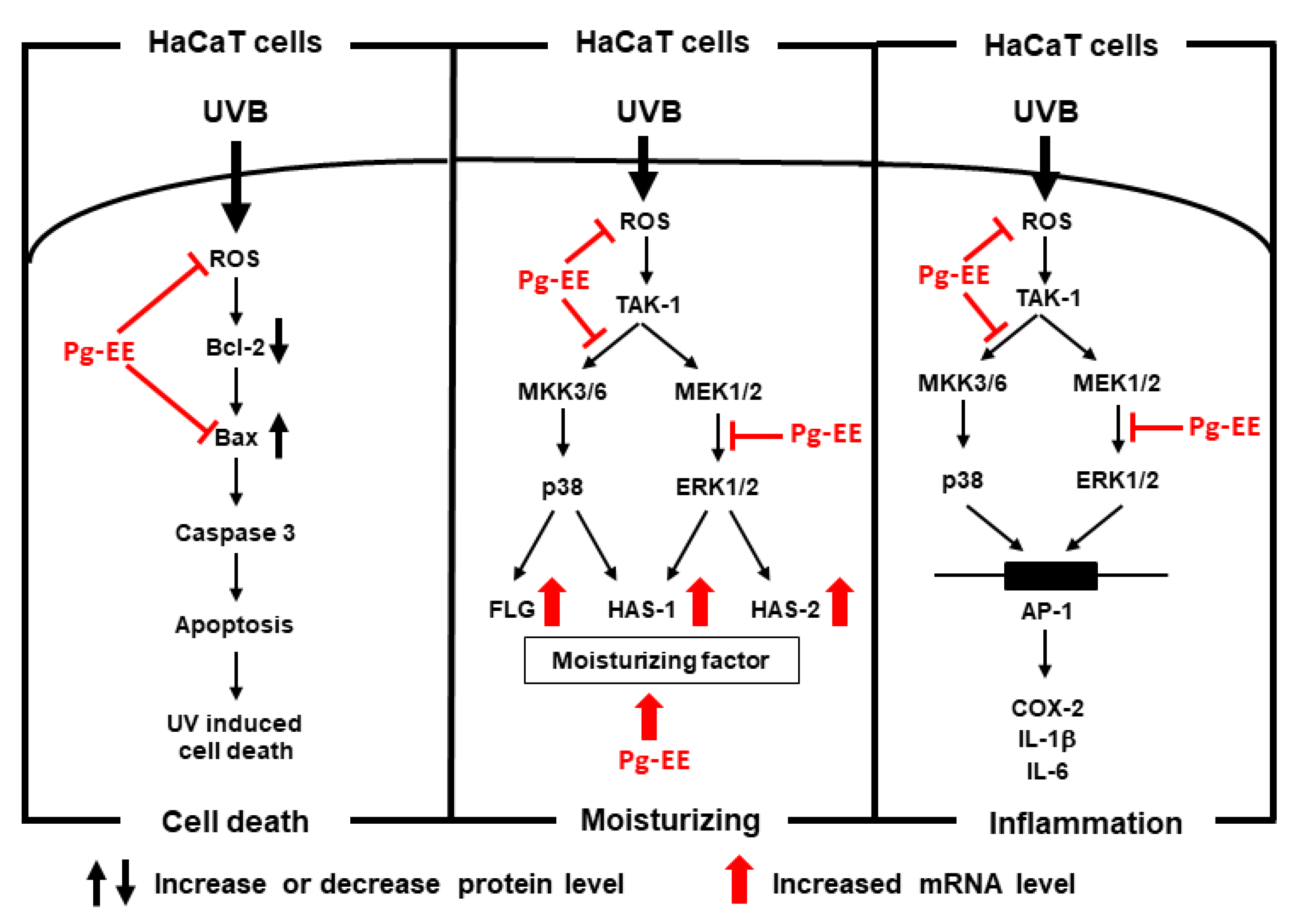
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Pg-EE and Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS)
4.3. Cell Culture
4.4. Compound Treatment
4.5. Cell Viability Assay
4.6. Nitric Oxide (NO) Assay
4.7. DPPH Assay
4.8. ABTS Radical Scavenging Assay
4.9. UVB Irradiation
4.10. Cell Morphology Photography
4.11. Immunoblotting Analysis
4.12. Analysis of mRNA Levels by Semi-Quantitative Reverse Transcription–Polymerase Chain Reaction (RT-PCR) and Quantitative Real-Time Polymerase Chain Reaction (Real-Time PCR)
4.13. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| FLG | Filaggrin |
| TGM-1 | Transglutaminase-1 |
| HAS | Hyaluronic acid synthase |
| MAPKs | Mitogen-activated protein kinases |
| ERK | Extracellular signal-regulated kinase |
| SC | Stratum corneum |
| LPS | Lipopolysaccharides |
| COX-2 | Cyclooxygenase-2 |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin 6 |
| TNF-α | Tumor necrosis factor alpha |
| MKK | Mitogen-activated protein kinase |
| MEK | MAPK or ERK kinases |
References
- Gould, J. Superpowered skin. Nature 2018, 563, S84–S85. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Di Meglio, P.; Qin, J.-Z.; Nickoloff, B.J. Skin immune sentinels in health and disease. Nat. Rev. Immunol. 2009, 9, 679–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoerter, J.D.; Bradley, P.; Casillas, A.; Chambers, D.; Denholm, C.; Johnson, K.; Weiswasser, B. Extrafollicular dermal melanocyte stem cells and melanoma. Stem Cells Int. 2012, 2012, 407079. [Google Scholar] [CrossRef] [Green Version]
- Bos, J.D.; Kapsenberg, M.L. The skin immune system Its cellular constituents and their interactions. Immunol. Today 1986, 7, 235–240. [Google Scholar] [CrossRef]
- Chaudhuri, R.K.; Bojanowski, K. Improvement of hydration and epidermal barrier function in human skin by a novel compound isosorbide dicaprylate. Int. J. Cosmet. Sci. 2017, 39, 518–526. [Google Scholar] [CrossRef]
- Eckhart, L.; Zeeuwen, P.L.J.M. The skin barrier: Epidermis vs environment. Exp. Dermatol. 2018, 27, 805–806. [Google Scholar] [CrossRef]
- BLANK, I.H. Factors which influence the water content of the stratum corneum. J. Investig. Dermatol. 1952, 18, 433–440. [Google Scholar] [CrossRef] [Green Version]
- Wildnauer, R.H.; Bothwell, J.W.; Douglass, A.B. Stratum corneum biomechanical properties. I. Influence of relative humidity on normal and extracted human stratum corneum. J. Investig. Dermatol. 1971, 56, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Van Duzee, B.F. The influence of water content, chemical treatment and temperature on the rheological properties of stratum corneum. J. Investig. Dermatol. 1978, 71, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Meirelles, N.C.; Yushmanov, V.E.; Tabak, M. Water increases the fluidity of intercellular membranes of stratum corneum: Correlation with water permeability, elastic, and electrical resistance properties. J. Investig. Dermatol. 1996, 106, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Bouwstra, J.A.; De Graaff, A.; Gooris, G.S.; Nijsse, J.; Wiechers, J.W.; Van Aelst, A.C. Water distribution and related morphology in human stratum corneum at different hydration levels. J. Investig. Dermatol. 2003, 120, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Sparr, E.; Millecamps, D.; Isoir, M.; Burnier, V.; Larsson, Å.; Cabane, B. Controlling the hydration of the skin though the application of occluding barrier creams. J. R. Soc. Interface 2013, 10, 20120788. [Google Scholar] [CrossRef]
- Kim, M.K.; Kim, E.J.; Cheng, Y.; Shin, M.H.; Oh, J.H.; Lee, D.H.; Chung, J.H. Inhibition of DNA methylation in the COL1A2 promoter by anacardic acid prevents UV-induced decrease of type I procollagen expression. J. Investig. Dermatol. 2017, 137, 1343–1352. [Google Scholar] [CrossRef] [Green Version]
- Cavinato, M.; Koziel, R.; Romani, N.; Weinmüllner, R.; Jenewein, B.; Hermann, M.; Dubrac, S.; Ratzinger, G.; Grillari, J.; Schmuth, M.; et al. UVB-induced senescence of human dermal fibroblasts involves impairment of proteasome and enhanced autophagic activity. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2017, 72, 632–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavinato, M.; Jansen-Dürr, P. Molecular mechanisms of UVB-induced senescence of dermal fibroblasts and its relevance for photoaging of the human skin. Exp. Gerontol. 2017, 94, 78–82. [Google Scholar] [CrossRef]
- Lin, P.; Hwang, E.; Ngo, H.T.T.; Seo, S.A.; Yi, T.H. Sambucus nigra L. ameliorates UVB-induced photoaging and inflammatory response in human skin keratinocytes. Cytotechnology 2019, 71, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Cavinato, M.; Waltenberger, B.; Baraldo, G.; Grade, C.V.C.; Stuppner, H.; Jansen-Dürr, P. Plant extracts and natural compounds used against UVB-induced photoaging. Biogerontology 2017, 18, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Trautinger, F. Mechanisms of photodamage of the skin and its functional consequences for skin ageing. Clin. Exp. Dermatol. 2001, 26, 573–577. [Google Scholar] [CrossRef]
- Lin, T.Y.; Wu, P.Y.; Hou, C.W.; Chien, T.Y.; Chang, Q.X.; Wen, K.C.; Lin, C.Y.; Chiang, H.M. Protective effects of sesamin against UVB-induced skin inflammation and photodamage in vitro and in vivo. Biomolecules 2019, 9, 479. [Google Scholar] [CrossRef] [Green Version]
- Pal, H.C.; Athar, M.; Elmets, C.A.; Afaq, F. Fisetin inhibits UVB-induced cutaneous inflammation and activation of PI3K/AKT/NFκB signaling pathways in SKH-1 hairless mice. Photochem. Photobiol. 2015, 91, 225–234. [Google Scholar] [CrossRef]
- Kuwano, T.; Kawano, S.; Kagawa, D.; Yasuda, Y.; Inoue, Y.; Murase, T. Dietary intake of glucono-δ-lactone attenuates skin inflammation and contributes to maintaining skin condition. Food Funct. 2018, 9, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.H.; Wu, P.Y.; Chen, C.W.; Lin, P.; Wen, K.C.; Lin, C.Y.; Chiang, H.M. N-(4-bromophenethyl) caffeamide protects skin from UVB-induced inflammation through MAPK/IL-6/NF-κB-dependent signaling in human skin fibroblasts and hairless mouse skin. Molecules 2017, 22, 1639. [Google Scholar] [CrossRef] [Green Version]
- Lou, L.; Li, Y.; Li, Y. Chemical constituents of the essential oil from Potentilla glabra var. mandshurica Maxim. J. Lanzhou Univ. Nat. Sci. 2004, 40, 58–60. [Google Scholar]
- Hu, Y.L.; Wang, W.Y.; Li, Y.W.; Xu, L.J.; Peng, Y. Research progress in Yaowang tea. Drugs Clin. 2013, 28, 236–241. [Google Scholar]
- Kim, H.; Shin, K.K.; Kim, H.G.; Jo, M.; Kim, J.K.; Lee, J.S.; Choung, E.S.; Li, W.Y.; Lee, S.W.; Kim, K.H.; et al. Src/nf-κb-targeted anti-inflammatory effects of Potentilla glabra var. Mandshurica (maxim.) hand.-mazz. ethanol extract. Biomolecules 2020, 10, 648. [Google Scholar] [CrossRef]
- Tobin, D.J. Introduction to skin aging. J. Tissue Viabil. 2017, 26, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plumb, J.A. Cell sensitivity assays: The MTT assay. Methods Mol. Med. 1999, 28, 25–30. [Google Scholar] [CrossRef]
- Katsuki, S.; Arnold, W.; Mittal, C.; Murad, F. Stimulation of guanylate cyclase by sodium nitroprusside, nitroglycerin and nitric oxide in various tissue preparations and comparison to the effects of sodium azide and hydroxylamine. J. Cyclic Nucleotide Res. 1977, 3, 23–35. [Google Scholar]
- Cole, P. The safe use of sodium nitroprusside. Anaesthesia 1978, 33, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Pyrzynska, K.; Pȩkal, A. Application of free radical diphenylpicrylhydrazyl (DPPH) to estimate the antioxidant capacity of food samples. Anal. Methods 2013, 5, 4288–4295. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Factors influencing the antioxidant activity determined by the ABTS+ radical cation assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Pouget, J.P.; Ravanat, J.L.; Sauvaigo, S. Effects of UV and visible radiations on cellular DNA. Curr. Probl. Dermatol. 2001, 29, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Aufiero, B.M.; Talwar, H.; Young, C.; Krishnan, M.; Hatfield, J.S.; Lee, H.K.; Wong, H.K.; Hamzavi, I.; Murakawa, G.J. Narrow-band UVB induces apoptosis in human keratinocytes. J. Photochem. Photobiol. B Biol. 2006, 82, 132–139. [Google Scholar] [CrossRef]
- Korsmeyer, S.J.; Shutter, J.R.; Veis, D.J.; Merry, D.E.; Oltvai, Z.N. Bcl-2/Bax: A rheostat that regulates an anti-oxidant pathway and cell death. Semin. Cancer Biol. 1993, 4, 327–332. [Google Scholar] [PubMed]
- Yin, X.M.; Oltvai, Z.N.; Korsmeyer, S.J. BH1 and BH2 domains of Bcl-2 are required for inhibition of apoptosis and heterodimerization with Bax. Nature 1994, 369, 321–323. [Google Scholar] [CrossRef]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 ratio considered as a prognostic marker with age and tumor location in colorectal cancer? Iran. Biomed. J. 2015, 19, 69–75. [Google Scholar] [CrossRef]
- Hector, S.; Prehn, J.H.M. Apoptosis signaling proteins as prognostic biomarkers in colorectal cancer: A review. Biochim. Biophys. Acta-Rev. Cancer 2009, 1795, 117–129. [Google Scholar] [CrossRef]
- Mohan, S.; Abdelwahab, S.I.; Kamalidehghan, B.; Syam, S.; May, K.S.; Harmal, N.S.M.; Shafifiyaz, N.; Hadi, A.H.A.; Hashim, N.M.; Rahmani, M.; et al. Involvement of NF-κB and Bcl2/Bax signaling pathways in the apoptosis of MCF7 cells induced by a xanthone compound pyranocycloartobiloxanthone A. Phytomedicine 2012, 19, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2015, 7, a008656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.; Yi, Y.S.; Lee, J.; Park, S.H.; Kim, S.; Hossain, M.A.; Jang, S.G.; Choi, Y.I.; Park, K.J.; Kim, D.S.; et al. Anti-apoptotic and anti-inflammatory activities of edible fresh water algae Prasiola japonica in UVB-irradiated skin keratinocytes. Am. J. Chin. Med. 2019, 47, 1853–1868. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.W.; Cheng, Y.C.; Hung, Y.C.; Lee, C.H.; Fang, J.Y.; Li, W.T.; Wu, Y.R.; Pan, T.L. Red raspberry extract protects the skin against UVB-induced damage with antioxidative and anti-inflammatory properties. Oxid. Med. Cell. Longev. 2019, 2019, 9529676. [Google Scholar] [CrossRef] [Green Version]
- Franco, L.; Marchena, A.; Rodríguez, A. Skin health properties of lycopene and melatonin. J. Dermatol. Ski. Sci. 2021, 3, 26–29. [Google Scholar] [CrossRef]
- Rumzhum, N.N.; Ammit, A.J. Cyclooxygenase 2: Its regulation, role and impact in airway inflammation. Clin. Exp. Allergy 2016, 46, 397–410. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Kondylis, V.; Kumari, S.; Vlantis, K.; Pasparakis, M. The interplay of IKK, NF-κB and RIPK 1 signaling in the regulation of cell death, tissue homeostasis and inflammation. Immunol. Rev. 2017, 277, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Novak, N. Atopic dermatitis and filaggrin. Curr. Opin. Immunol. 2016, 42, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Chung, S.I.; Yoneda, K.; Steinert, P.M. Expression of transglutaminase 1 in human epidermis. J. Investig. Dermatol. 1995, 104, 211–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, M.L.; Farasat, S.; Steinbach, P.J.; Wei, M.H.; Toure, O.; Fleckman, P.; Blake, P.; Bale, S.J.; Toro, J.R. Transglutaminase-1 gene mutations in autosomal recessive congenital ichthyosis: Summary of mutations(including 23 novel) and modeling of TGase-1. Hum. Mutat. 2009, 30, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Farasat, S.; Wei, M.H.; Herman, M.; Liewehr, D.J.; Steinberg, S.M.; Bale, S.J.; Fleckman, P.; Toro, J.R. Novel transglutaminase-1 mutations and genotypephenotype investigations of 104 patients with autosomal recessive congenital ichthyosis in the USA. J. Med. Genet. 2009, 46, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Siiskonen, H.; Oikari, S.; Pasonen-Seppänen, S.; Rilla, K. Hyaluronan synthase 1: A mysterious enzyme with unexpected functions. Front. Immunol. 2015, 6, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaisse, J.; Bourguignon, V.; De Vuyst, E.; Lambert De Rouvroit, C.; Nikkels, A.F.; Flamion, B.; Poumay, Y. Hyaluronan metabolism in human keratinocytes and atopic dermatitis skin is driven by a balance of hyaluronan synthases 1 and 3. J. Investig. Dermatol. 2014, 134, 2174–2182. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Gu, L.; Liu, S.; Jin, F. Magnesia reactivity on activating efficacy for ground granulated blastfurnace slag for soft clay stabilisation. Appl. Clay Sci. 2016, 126, 57–62. [Google Scholar] [CrossRef]
- Lee, D.E.; Ayoub, N.; Agrawal, D.K. Mesenchymal stem cells and cutaneous wound healing: Novel methods to increase cell delivery and therapeutic efficacy. Stem Cell Res. Ther. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaidbey, K.H.; Agin, P.P.; Sayre, R.M.; Kligman, A.M. Photoprotection by melanin—A comparison of black and Caucasian skin. J. Am. Acad. Dermatol. 1979, 1, 249–260. [Google Scholar] [CrossRef]
- Han, A.; Chien, A.L.; Kang, S. Photoaging. Dermatol. Clin. 2014, 32, 291–299. [Google Scholar] [CrossRef]
- Lam, S.M. A new paradigm for the aging face. Textb. Aging Ski. 2010, 27, 1139–1145. [Google Scholar] [CrossRef]
- Gordon, J.R.S.; Brieva, J.C. Unilateral dermatoheliosis. N. Engl. J. Med. 2012, 366, e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riahi, R.R.; Bush, A.E.; Cohen, P.R. Topical retinoids: Therapeutic mechanisms in the treatment of photodamaged skin. Am. J. Clin. Dermatol. 2016, 17, 265–276. [Google Scholar] [CrossRef]
- Griffiths, C.E.M. The role of retinoids in the prevention and repair of aged and photoaged skin. Clin. Exp. Dermatol. 2001, 26, 613–618. [Google Scholar] [CrossRef]
- Darlenski, R.; Surber, C.; Fluhr, J.W. Topical retinoids in the management of photodamaged skin: From theory to evidence-based practical approach. Br. J. Dermatol. 2010, 163, 1157–1165. [Google Scholar] [CrossRef]
- Mukherjee, S.; Date, A.; Patravale, V.; Korting, H.C.; Roeder, A.; Weindl, G. Retinoids in the treatment of skin aging: An overview of clinical efficacy and safety. Clin. Interv. Aging 2006, 1, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Poon, F.; Kang, S.; Chien, A.L. Mechanisms and treatments of photoaging. Photodermatol. Photoimmunol. Photomed. 2015, 31, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Oh, S.-K. Studies on the anti-aging action of Korean ginseng. Korean J. Food Sci. Technol. 1985, 17, 506–515. [Google Scholar]
- Cui, X.; Ma, Y.; Wang, H.; Huang, J.; Li, L.; Tang, J.; Cheng, B. The anti-photoaging effects of pre- and post-treatment of platelet-rich plasma on UVB-damaged HaCaT keratinocytes. Photochem. Photobiol. 2021, 97, 589–599. [Google Scholar] [CrossRef]
- Chiu, L.Y.; Wu, N.L.; Hung, C.F.; Bai, P.; Dai, Y.S.; Lin, W.W. PARP-1 involves in UVB-induced inflammatory response in keratinocytes and skin injury via regulation of ROS-dependent EGFR transactivation and p38 signaling. FASEB J. 2021, 35, e21393. [Google Scholar] [CrossRef]
- Ko, H.J.; Kim, J.; Ahn, M.; Kim, J.H.; Lee, G.S.; Shin, T. Ergothioneine alleviates senescence of fibroblasts induced by UVB damage of keratinocytes via activation of the Nrf2/HO-1 pathway and HSP70 in keratinocytes. Exp. Cell Res. 2021, 400, 112516. [Google Scholar] [CrossRef]
- Kim, J.K.; Choi, E.; Hong, Y.H.; Kim, H.; Jang, Y.J.; Lee, J.S.; Choung, E.S.; Woo, B.Y.; Hong, Y.D.; Lee, S.; et al. Syk/NF-κB-targeted anti-inflammatory activity of Melicope accedens (Blume) T.G. Hartley methanol extract. J. Ethnopharmacol. 2021, 271, 113887. [Google Scholar] [CrossRef]
- Baik, J.S.; Seo, Y.N.; Yi, J.M.; Rhee, M.H.; Park, M.T.; Kim, S.D. Ginsenoside-Rp1 inhibits radiation-induced effects in lipopolysaccharide-stimulated J774A.1 macrophages and suppresses phenotypic variation in CT26 colon cancer cells. J. Ginseng Res. 2020, 44, 843–848. [Google Scholar] [CrossRef]
- Park, Y.S.; Lee, J.E.; Park, J.I.; Myung, C.H.; Lim, Y.H.; Park, C.K.; Hwang, J.S. Inhibitory mechanism of ginsenoside Rh3 on granulocyte-macrophage colony-stimulating factor expression in UV-B-irradiated murine SP-1 keratinocytes. J. Ginseng Res. 2020, 44, 274–281. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Rho, T.; Jeong, H.W.; Hong, Y.D.; Yoon, K.; Cho, J.Y.; Yoon, K.D. Identification of a novel triterpene saponin from Panax ginseng seeds, pseudoginsenoside RT8, and its antiinflammatory activity. J. Ginseng Res. 2020, 44, 145–153. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ to 3′) | |
|---|---|---|
| Semi-quantitative RT-PCR | ||
| FLG | F | AGGGAAGATCCAAGAGCCCA |
| R | ACTCTGGATCCCCTACGCTT | |
| TGM-1 | F | GAAATGCGGCAGATGACGAC |
| R | AACTCCCCAGCGTCTGATTG | |
| HAS-1 | F | CCACCCAGTACAGCGTCAAC |
| R | CATGGTGCTTCTGTCGCTCT | |
| HAS-2 | F | TTCTTTATGTGACTCATCTGTCTCACCGG |
| R | ATTGTTGGCTACCAGTTTATCCAAACG | |
| COX-2 | F | GGGATTTTGGAACGTTGTGAA |
| R | CGACATTGTAAGTTGGTGGACTGT | |
| IL-1β | F | ATTTGAATTCCCTGGGTGAG |
| R | CCTCATCCTGGAAGGTCCAC | |
| IL-6 | F | TACCCCCAGGAGAAGATTCC |
| R | TTTTCTGCCAGTGCCTCTTT | |
| GAPDH | F | GCACCGTCAAGGCTGAGAAC |
| R | ATGGTGGTGAAGACGCCAGT | |
| Quantitative real-time PCR | ||
| FLG | F | GGGCACTGAAAGGCAAAAAG |
| R | CACCATAATCATAATCTGCACTACCA | |
| HAS-1 | F | CAGCCTGCGATACTGGGTAG |
| R | GCCGGTCATCCCCAAAAGTA | |
| SOD-1 | F | AAGCGGTGAACCAGTTGTGT |
| R | GCCAATGATGGAATGCTCTC | |
| Nrf-2 | F | ACATCCTTTGGAGGCAAGAC |
| R | TCGGGTCATTGTGAGTCAGT | |
| GAPDH | F | CACTCACGGCAAATTCAACGGC |
| R | GACTCCACGACATACTCAGCA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, L.; Kim, M.-Y.; Cho, J.Y. Protective Effect of Potentilla glabra in UVB-Induced Photoaging Process. Molecules 2021, 26, 5408. https://doi.org/10.3390/molecules26175408
You L, Kim M-Y, Cho JY. Protective Effect of Potentilla glabra in UVB-Induced Photoaging Process. Molecules. 2021; 26(17):5408. https://doi.org/10.3390/molecules26175408
Chicago/Turabian StyleYou, Long, Mi-Yeon Kim, and Jae Youl Cho. 2021. "Protective Effect of Potentilla glabra in UVB-Induced Photoaging Process" Molecules 26, no. 17: 5408. https://doi.org/10.3390/molecules26175408
APA StyleYou, L., Kim, M. -Y., & Cho, J. Y. (2021). Protective Effect of Potentilla glabra in UVB-Induced Photoaging Process. Molecules, 26(17), 5408. https://doi.org/10.3390/molecules26175408