
Antioxidant Metabolites in Primitive, Wild, and Cultivated Citrus and Their Role in Stress Tolerance




Abstract
:1. Introduction
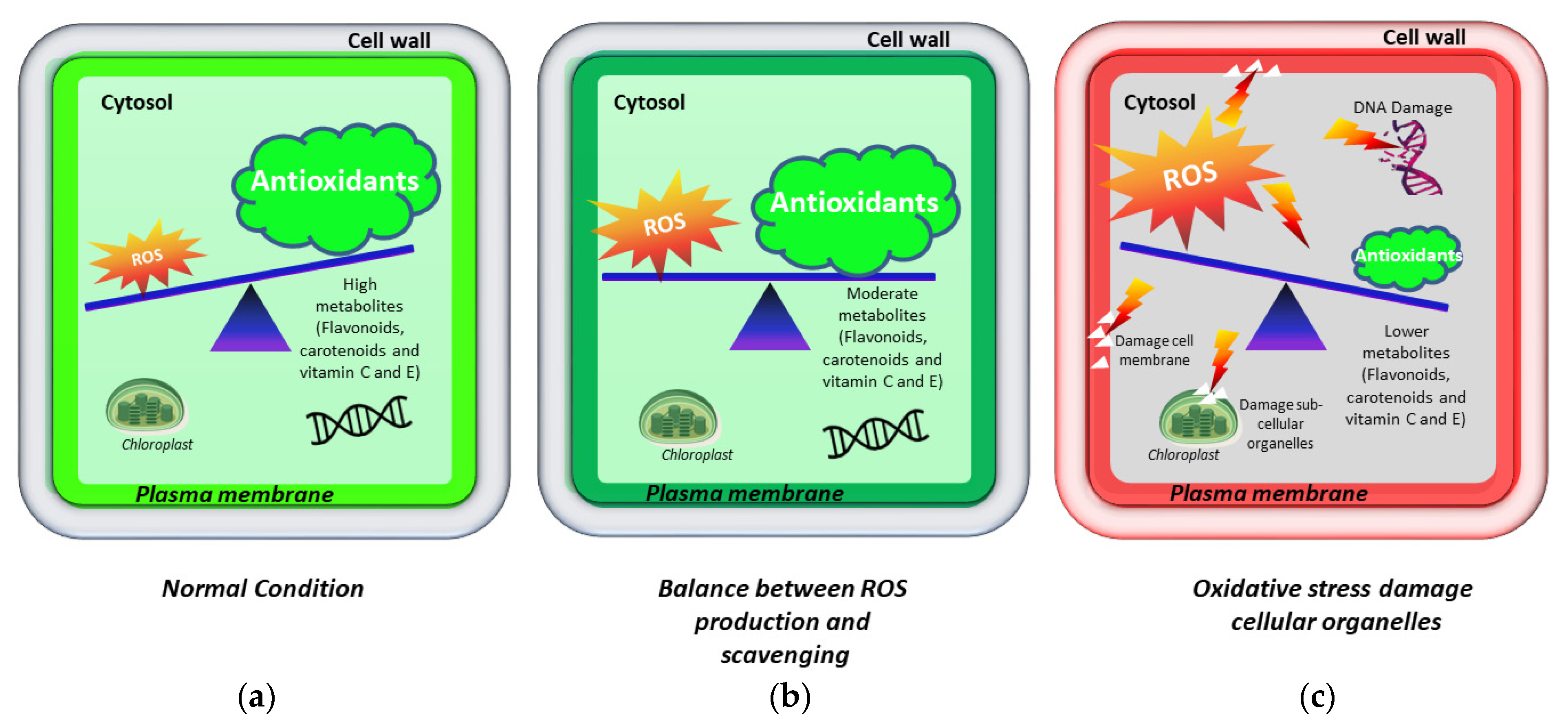
2. Importance of Antioxidants
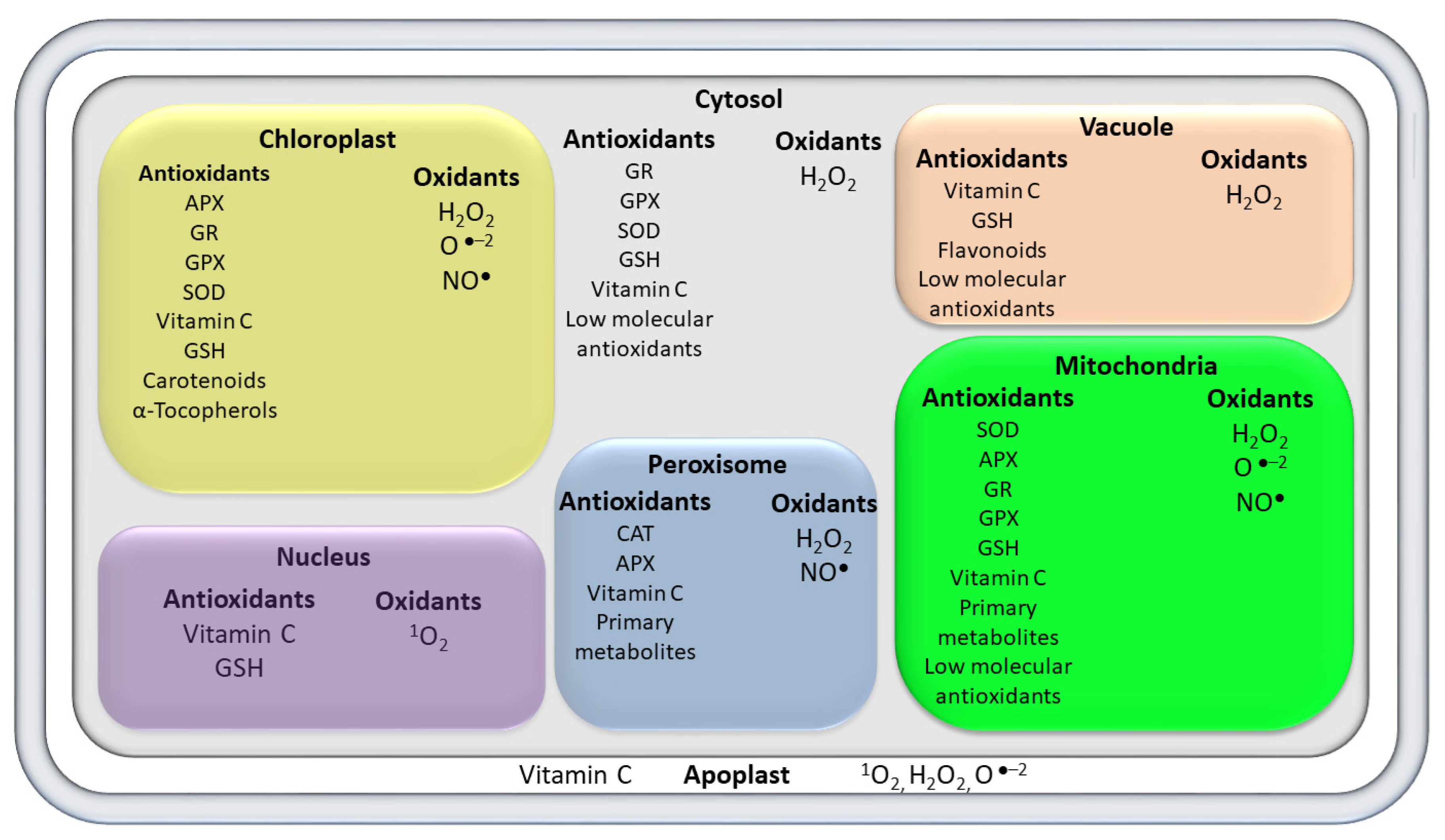
3. The Producing Sites and Scavenging of ROS
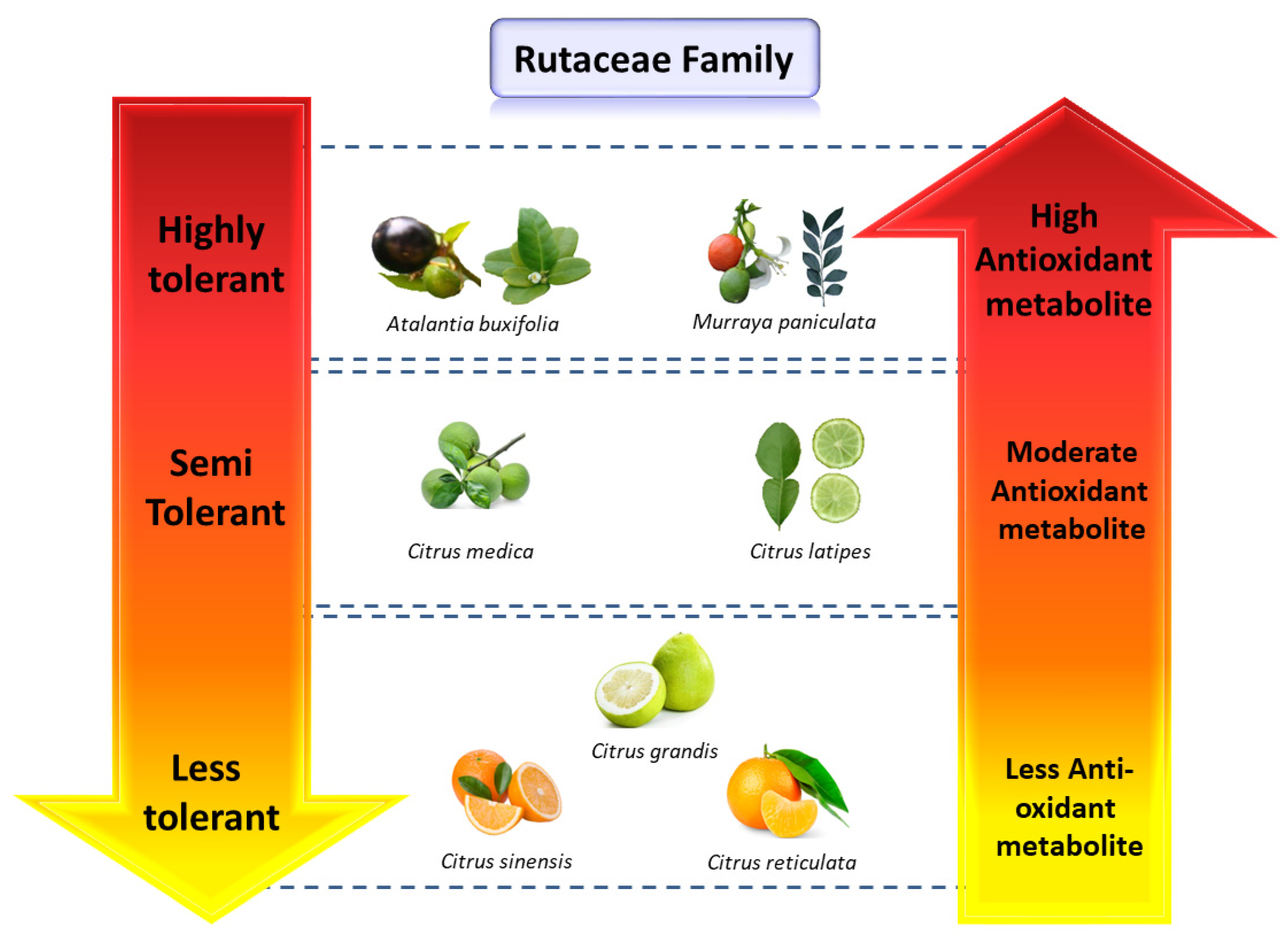
4. Diversity of Antioxidant Metabolites in Citrus
4.1. Antioxidant Volatiles and Fatty Acids
4.2. Antioxidant Alkaloids, Coumarins, and Limonoids
4.3. Antioxidant Carotenoids, Ascorbic Acid, and Tocopherols in Citrus
4.4. Antioxidant Amino Acids
4.5. Hydroxycinnamic Acids and Their Derivatives
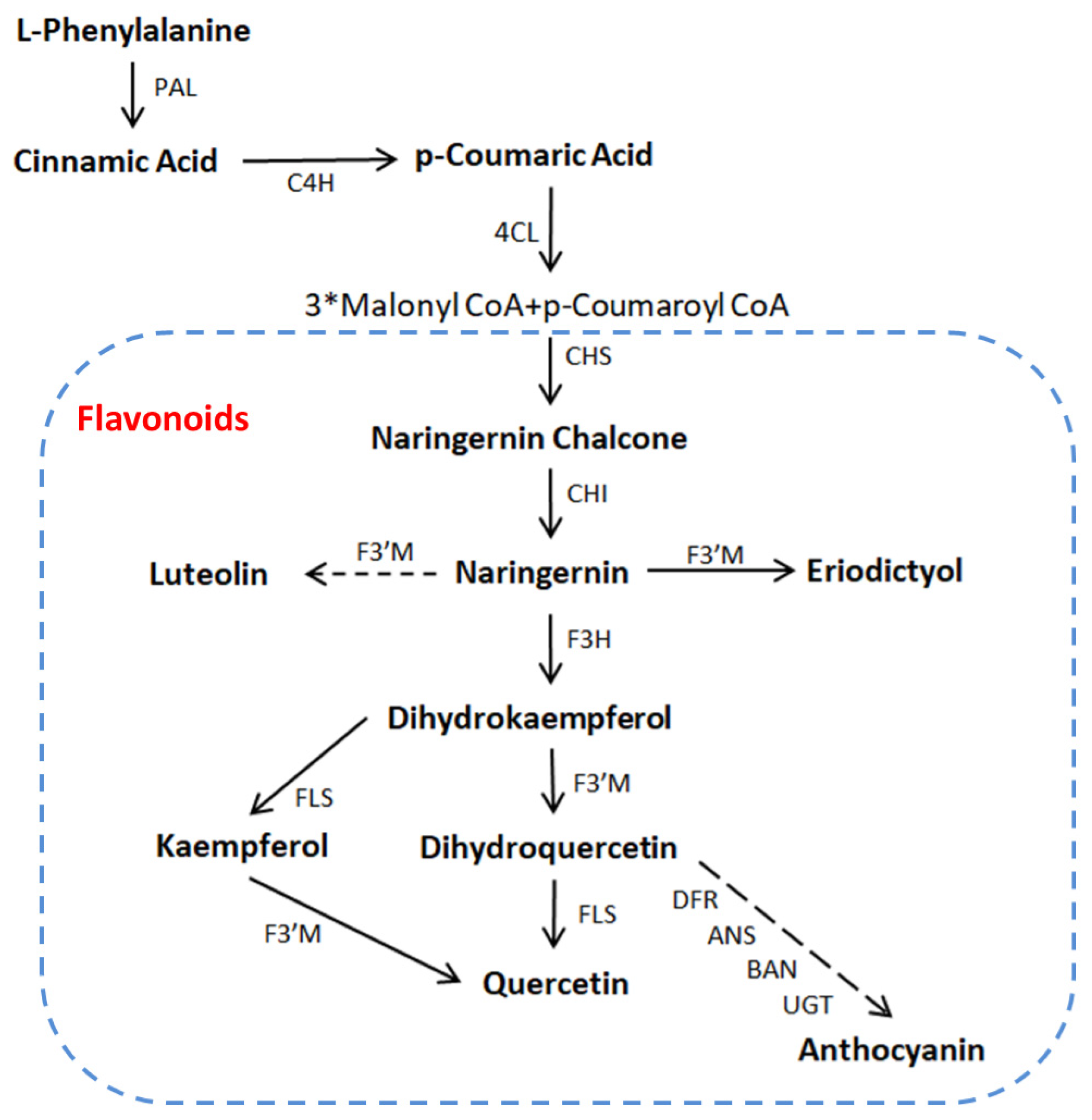
4.6. Antioxidant Flavonoids
5. Therapeutic Applications of Citrus Fruits and Potential Usage of Citrus Wastes
6. Genomic Features of Citrus Species
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Heying, E.; Tanumihardjo, S.A. History, global distribution, and nutritional importance of citrus fruits. Compr. Rev. Food Sci. Food Saf. 2012, 11, 530–545. [Google Scholar] [CrossRef]
- Hussain, S.Z.; Naseer, B.; Qadri, T.; Fatima, T.; Bhat, T.A. Citrus Fruits—Morphology, Taxonomy, Composition and Health Benefits. In Fruits Grown in Highland Regions of the Himalayas; Springer: Cham, Switzerland, 2021; pp. 229–244. [Google Scholar] [CrossRef]
- Al-snafi, A.E. Nutritional value and pharmacological importance of citrus species grown in Iraq. IOSR J. Pharm. 2016, 6, 76–108. [Google Scholar] [CrossRef]
- Dala Paula, B.M.; Raithore, S.; Manthey, J.A.; Baldwin, E.A.; Bai, J.; Zhao, W.; Glória, M.B.A.; Plotto, A. Active taste compounds in juice from oranges symptomatic for Huanglongbing (HLB) citrus greening disease. LWT—Food Sci. Technol. 2018, 91, 518–525. [Google Scholar] [CrossRef]
- Marhuenda, J.; Cerdá, B.; Villaño, D.; Galindo, A.; Zafrilla, P. Citrus and health. In Citrus–Health Benefits and Production Technology; Intechopen: London, UK, 2018; pp. 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Zhang, L.; Sugiura, M.; Kato, M. The Genus Citrus; Elsevier: Amsterdam, The Netherlands, 2020; pp. 495–511. [Google Scholar]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Vun, O.; Fn, M.; Cn, I.; Ku, U. Stematic characterization of six citrus species using petiole anatomy. Med. Aromat. Plants 2015, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Dave, R. In vitro models for antioxidant activity evaluation and some medicinal plants possessing antioxidant properties: An overview. Afr. J. Microbiol. Res. 2009, 3, 981–996. [Google Scholar]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Khalid, M.F.; Saqib, M.; Ahmad, S.; Zafar, W.; Rao, M.J.; Morillon, R.; Anjum, M.A. Drought tolerance in citrus rootstocks is associated with better antioxidant defense mechanism. Acta Physiol. Plant. 2018, 40, 135. [Google Scholar] [CrossRef]
- Racchi, M.L. Antioxidant defenses in plants with attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Sales, C.; Beltrán, J.; Gómez-Cadenas, A.; Arbona, V. Activation of secondary metabolism in citrus plants is associated to sensitivity to combined drought and high temperatures. Front. Plant Sci. 2017, 7, 1954. [Google Scholar] [CrossRef] [Green Version]
- Khalid, M.F.; Morillon, R.; Anjum, M.A.; Ejaz, S.; Rao, M.J.; Hussain, S. Volkamer lemon tetraploid rootstock transmits the salt tolerance when grafted with diploid kinnow mandarin by strong antioxidant defense mechanism and efficient osmotic adjustment. J Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Tang, X.; Wang, X.; Huang, Y.; Ma, L.; Jiang, X.; Rao, M.J.; Xu, Y.; Yin, P.; Yuan, M.; Deng, X.; et al. Natural variations of TFIIAγ gene and LOB1 promoter contribute to citrus canker disease resistance in Atalantia buxifolia. PLoS Genet. 2021, 17, e1009316. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.J.; Ding, F.; Wang, N.; Deng, X.; Xu, Q. Metabolic mechanisms of host species against citrus Huanglongbing (Greening Disease). CRC Crit. Rev. Plant Sci. 2018, 37, 496–511. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Rouseff, R.; Reyes-De-Corcuera, J.I. Untargeted metabolite analysis of healthy and Huanglongbing-infected orange leaves by CE-DAD. Electrophoresis 2009, 30, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Nettore, I.C.; Rocca, C.; Mancino, G.; Albano, L.; Amelio, D.; Grande, F.; Puoci, F.; Pasqua, T.; Desiderio, S.; Rosa, M.; et al. Quercetin and its derivative Q2 modulate chromatin dynamics in adipogenesis and Q2 prevents obesity and metabolic disorders in rats. J. Nutr. Biochem. 2019, 69, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.J.; Xu, Y.; Tang, X.; Huang, Y.; Liu, J.; Deng, X. CsCYT75B1, a citrus CYTOCHROME P450 gene, is involved in accumulation of antioxidant flavonoids and induces drought tolerance in transgenic Arabidopsis. Antioxidants 2020, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayor, R. Oxidative stress and antioxidant defense system. J. Inst. Med. Trop. 2010, 5, 23–39. [Google Scholar]
- Hussain, S.; Rao, M.J.; Anjum, M.A.; Ejaz, S.; Zakir, I.; Ali, M.A.; Ahmad, N.; Ahmad, S. Oxidative stress and antioxidant defense in plants under drought conditions. In Plant Abiotic Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2019; pp. 207–219. [Google Scholar]
- Capanoglu, E.; Kamiloglu, S.; Ozkan, G.; Apak, R. Evaluation of antioxidant activity/capacity measurement methods for food products. In Measurement of Antioxidant Activity and Capacity: Recent Trends and Applications; John Wiley & Sons Ltd.: Chicester, UK, 2017; pp. 273–286. [Google Scholar]
- Canan, İ.; Gündoğdu, M.; Seday, U.; Oluk, C.A.; Karaşahin, Z.; Eroğlu, E.Ç.; Yazici, E.; Ünlü, M. Determination of antioxidant, total phenolic, total carotenoid, lycopene, ascorbic acid, and sugar contents of citrus species and mandarin hybrids. Turk. J. Agric. For. 2016, 40, 894–899. [Google Scholar] [CrossRef]
- Iacopetta, D.; Grande, F.; Caruso, A.; Mordocco, R.A.; Plutino, M.R.; Scrivano, L.; Ceramella, J.; Muià, N.; Saturnino, C.; Puoci, F.; et al. New insights for the use of quercetin analogs in cancer treatment. Future Med. Chem. 2017, 9, 2011–2028. [Google Scholar] [CrossRef] [PubMed]
- Alarcón-Flores, M.I.; Romero-González, R.; Vidal, J.L.M.; Frenich, A.G. Multiclass determination of phytochemicals in vegetables and fruits by ultra high performance liquid chromatography coupled to tandem mass spectrometry. Food Chem. 2013, 141, 1120–1129. [Google Scholar] [CrossRef]
- Jambunathan, N. Determination and detection of reactive oxygen species (ROS), lipid peroxidation, and electrolyte leakage in plants. In Plant Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2010; pp. 291–297. [Google Scholar]
- Nath, M.; Bhatt, D.; Prasad, R.; Tuteja, N. Reactive oxygen species (ROS) metabolism and signaling in plant-mycorrhizal association under biotic and abiotic stress conditions. In Mycorrhiza-Eco-Physiology, Secondary Metabolites, Nanomaterials; Springer: Berlin/Heidelberg, Germany, 2017; pp. 223–232. [Google Scholar]
- Luis, A.; Sandalio, L.M.; Corpas, F.J.; Palma, J.M.; Barroso, J.B. Reactive oxygen species and reactive nitrogen species in peroxisomes. Production, scavenging, and role in cell signaling. Plant Physiol. 2006, 141, 330–335. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Carocho, M.; Ferreira, I.C.F.R. A review on antioxidants, prooxidants and related controversy: Natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casacchia, T.; Occhiuzzi, M.A.; Grande, F.; Rizzuti, B.; Granieri, M.C.; Rocca, C.; Gattuso, A.; Garofalo, A.; Angelone, T.; Statti, G. A pilot study on the nutraceutical properties of the Citrus hybrid Tacle® as a dietary source of polyphenols for supplementation in metabolic disorders. J. Funct. Foods 2019, 52, 370–381. [Google Scholar] [CrossRef]
- Ebrahimabadi, A.H.; Movahedpour, M.M.; Batooli, H.; Ebrahimabadi, E.H.; Mazoochi, A.; Qamsari, M.M. Volatile compounds analysis and antioxidant, antimicrobial and cytotoxic activities of Mindium laevigatum. Iran. J. Basic Med. Sci. 2016, 19, 1337. [Google Scholar]
- Javed, S.; Ahmad, R.; Shahzad, K.; Nawaz, S.; Saeed, S.; Saleem, Y. Chemical constituents, antimicrobial and antioxidant activity of essential oil of Citrus limetta var. Mitha (sweet lime) peel in Pakistan. Afr. J. Microbiol. Res. 2013, 7, 3071–3077. [Google Scholar]
- González-Mas, M.C.; Rambla, J.L.; Alamar, M.C.; Gutiérrez, A.; Granell, A. Comparative analysis of the volatile fraction of fruit juice from different Citrus species. PLoS ONE 2011, 6, e22016. [Google Scholar] [CrossRef] [Green Version]
- Sarrou, E.; Chatzopoulou, P.; Dimassi-Theriou, K.; Therios, I. Volatile constituents and antioxidant activity of peel, flowers and leaf oils of Citrus aurantium L. growing in Greece. Molecules 2013, 18, 10639–10647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espina, L.; Somolinos, M.; Lorán, S.; Conchello, P.; García, D.; Pagán, R. Chemical composition of commercial citrus fruit essential oils and evaluation of their antimicrobial activity acting alone or in combined processes. Food Control 2011, 22, 896–902. [Google Scholar] [CrossRef]
- Miyazawa, N.; Fujita, A.; Kubota, K. Aroma character impact compounds in Kinokuni mandarin orange (Citrus kinokuni) compared with Satsuma mandarin orange (Citrus unshiu). Biosci. Biotechnol. Biochem. 2010, 74, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Tezcan, F.; Gültekin-Özgüven, M.; Diken, T.; Özçelik, B.; Erim, F.B. Antioxidant activity and total phenolic, organic acid and sugar content in commercial pomegranate juices. Food Chem. 2009, 115, 873–877. [Google Scholar] [CrossRef]
- Kabera, J.N.; Semana, E.; Mussa, A.R.; He, X. Plant secondary metabolites: Biosynthesis, classification, function and pharmacological properties. J. Pharm. Pharmacol. 2014, 2, 377–392. [Google Scholar]
- Servillo, L.; Giovane, A.; Balestrieri, M.L.; Casale, R.; Cautela, D.; Castaldo, D. Citrus genus plants contain N-methylated tryptamine derivatives and their 5-hydroxylated forms. J. Agric. Food Chem. 2013, 61, 5156–5162. [Google Scholar] [CrossRef]
- Percy, D.W.; Adcock, J.L.; Conlan, X.A.; Barnett, N.W.; Gange, M.E.; Noonan, L.K.; Henderson, L.C.; Francis, P.S. Determination of Citrus aurantium protoalkaloids using HPLC with acidic potassium permanganate chemiluminescence detection. Talanta 2010, 80, 2191–2195. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Preuss, H.G.; Shara, M. The safety of Citrus aurantium (bitter orange) and its primary protoalkaloid p-synephrine. Phyther. Res. 2011, 25, 1421–1428. [Google Scholar] [CrossRef]
- Dugrand-Judek, A.; Olry, A.; Hehn, A.; Costantino, G.; Ollitrault, P.; Froelicher, Y.; Bourgaud, F. The distribution of coumarins and furanocoumarins in Citrus species closely matches Citrus phylogeny and reflects the organization of biosynthetic pathways. PLoS ONE 2015, 10, e0142757. [Google Scholar]
- Russo, M.; Torre, G.; Carnovale, C.; Bonaccorsi, I.; Mondello, L.; Dugo, P. A new HPLC method developed for the analysis of oxygen heterocyclic compounds in Citrus essential oils. J. Essent. Oil Res. 2012, 24, 119–129. [Google Scholar] [CrossRef]
- Okuyama, S.; Minami, S.; Shimada, N.; Makihata, N.; Nakajima, M.; Furukawa, Y. Anti-inflammatory and neuroprotective effects of auraptene, a citrus coumarin, following cerebral global ischemia in mice. Eur. J. Pharmacol. 2013, 699, 118–123. [Google Scholar] [CrossRef]
- Masamoto, Y.; Ando, H.; Murata, Y.; Shimoishi, Y.; Tada, M.; Takahata, K. Mushroom tyrosinase inhibitory activity of esculetin isolated from seeds of Euphorbia lathyris L. Biosci. Biotechnol. Biochem. 2003, 67, 631–634. [Google Scholar] [CrossRef]
- Manners, G.D. Citrus limonoids: Analysis, bioactivity, and biomedical prospects. J. Agric. Food Chem. 2007, 55, 8285–8294. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhou, Z. Antioxidant activity of Citrus fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.F.; Hamdan, D.I.; Wink, M.; El-Shazly, A.M. Hepatoprotective effect of limonin, a natural limonoid from the seed of Citrus aurantium var. bigaradia, on D-galactosamine-induced liver injury in rats. Naunyn. Schmiedebergs. Arch. Pharmacol. 2014, 387, 251–261. [Google Scholar] [CrossRef]
- Poulose, S.M.; Harris, E.D.; Patil, B.S. Citrus limonoids induce apoptosis in human neuroblastoma cells and have radical scavenging activity. J. Nutr. 2005, 135, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- Poiroux-Gonord, F.; Fanciullino, A.; Bert, L.; Urban, L. Effect of fruit load on maturity and carotenoid content of clementine (Citrus clementina Hort. ex Tan.) fruits. J. Sci. Food Agric. 2012, 92, 2076–2083. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.L. Carotenoids—Antioxidant Properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N. Vitamin C: The metabolism and functions of ascorbic acid in plants. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2011; Volume 59, pp. 107–177. ISBN 0065-2296. [Google Scholar]
- Wang, Z.; Xiao, Y.; Chen, W.; Tang, K.; Zhang, L. Increased vitamin C content accompanied by an enhanced recycling pathway confers oxidative stress tolerance in Arabidopsis. J. Integr. Plant Biol. 2010, 52, 400–409. [Google Scholar] [CrossRef]
- Zechmann, B.; Stumpe, M.; Mauch, F. Immunocytochemical determination of the subcellular distribution of ascorbate in plants. Planta 2011, 233, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Luo, T.; Liu, C.; Wang, Y.; Yang, H.; Yang, W.; Zheng, L.; Xiao, X.; Zhang, M.; Xu, R.; et al. An R2R3-MYB transcription factor represses the transformation of a- and b-branch carotenoids by negatively regulating expression of CrBCH2 and CrNCED5 in flavedo of Citrus reticulata. Funct. Integr. Genom. 2017, 14, 177–189. [Google Scholar]
- Ito, T.; Fujimoto, S.; Suito, F.; Shimosaka, M.; Taguchi, G. C-Glycosyltransferases catalyzing the formation of di-C-glucosyl flavonoids in citrus plants. Plant J. 2017, 91, 187–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Liu, X.; Zhang, H.; He, Z.; Yang, H.; Chen, J.; Feng, J.; Yang, W.; Jiang, Y.; Yao, J.-L. The IAA-and ABA-responsive transcription factor CgMYB58 upregulates lignin biosynthesis and triggers juice sac granulation in pummelo. Hortic. Res. 2020, 7, 139. [Google Scholar] [CrossRef] [PubMed]
- Catalano, C.; Ciacciulli, A.; Salonia, F.; Russo, M.P.; Caruso, P.; Caruso, M.; Russo, G.; Distefano, G.; Licciardello, C. Target-genes reveal species and genotypic specificity of anthocyanin pigmentation in citrus and related genera. Genes 2020, 11, 807. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Tang, Z.; Fu, J.; Yuan, Y.; Deng, X.; Xu, Q. CsMYB3 and CsRuby1 form an ‘activator-and-repressor’loop for regulation of anthocyanin biosynthesis in citrus. Plant Cell Physiol. 2019, 61, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhu, K.; Sun, Q.; Zhang, W.; Wang, X.; Cao, H.; Tan, M.; Xie, Z.; Zeng, Y.; Ye, J. Natural variation in CCD4 promoter underpins species-specific evolution of red coloration in citrus peel. Mol. Plant 2019, 12, 1294–1307. [Google Scholar] [CrossRef]
- Lu, S.; Zhang, Y.; Zhu, K.; Yang, W.; Ye, J.; Chai, L.; Xu, Q.; Deng, X. The citrus transcription factor CsMADS6 modulates carotenoid metabolism by directly regulating carotenogenic genes. Plant Physiol. 2018, 176, 2657–2676. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.J.; Xu, Y.; Huang, Y.; Tang, X.; Deng, X.; Xu, Q. Ectopic expression of citrus UDP-GLUCOSYL TRANSFERASE gene enhances anthocyanin and proanthocyanidins contents and confers high light tolerance in Arabidopsis. BMC Plant Biol. 2019, 19, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, S.B.; Guo, L.-X.; Shi, C.-Y.; Khan, M.A.; Bai, Y.-X.; Du, W.; Liu, Y.-Z. Assessment of sugar and sugar accumulation-related gene expression profiles reveal new insight into the formation of low sugar accumulation trait in a sweet orange (Citrus sinensis) bud mutant. Mol. Biol. Rep. 2020, 47, 2781–2791. [Google Scholar] [CrossRef]
- Kita, M.; Hirata, Y.; Moriguchi, T.; Endo-inagaki, T.; Matsumoto, R.; Hasegawa, S.; Suhayda, C.G.; Omura, M. Molecular cloning and characterization of a novel gene encoding limonoid UDP-glucosyltransferase in Citrus. Febs Lett. 2000, 469, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Daniel, J.J.; Owens, D.K.; McIntosh, C.A. Secondary product glucosyltransferase and putative glucosyltransferase expression during Citrus paradisi (cv Duncan) growth and development. Phytochemistry 2011, 72, 1732–1738. [Google Scholar] [CrossRef]
- Munné-Bosch, S. The role of α-tocopherol in plant stress tolerance. J. Plant Physiol. 2005, 162, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Weiler, E.W.; Alegre, L.; Müller, M.; Düchting, P.; Falk, J. α-Tocopherol may influence cellular signaling by modulating jasmonic acid levels in plants. Planta 2007, 225, 681–691. [Google Scholar] [CrossRef]
- Hincha, D.K. Effects of α-tocopherol (vitamin E) on the stability and lipid dynamics of model membranes mimicking the lipid composition of plant chloroplast membranes. FEBS Lett. 2008, 582, 3687–3692. [Google Scholar] [CrossRef] [Green Version]
- Killiny, N. Metabolite signature of the phloem sap of fourteen citrus varieties with different degrees of tolerance to Candidatus Liberibacter asiaticus. Physiol. Mol. Plant Pathol. 2017, 97, 20–29. [Google Scholar] [CrossRef]
- Killiny, N.; Hijaz, F. Amino acids implicated in plant defense are higher in Candidatus Liberibacter asiaticus-tolerant citrus varieties. Plant Signal. Behav. 2016, 11, e1171449. [Google Scholar] [CrossRef] [Green Version]
- Hijaz, F.; Nehela, Y.; Killiny, N. Possible role of plant volatiles in tolerance against huanglongbing in citrus. Plant Signal. Behav. 2016, 11, e1138193. [Google Scholar] [CrossRef] [Green Version]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Rao, M.J.; Anjum, M.A.; Ejaz, S.; Umar, U.U.; Ali, M.A.; Khalid, M.F.; Sohail, M.; Ercisli, S.; Zia-Ul-Haq, M.; et al. Effect of different combinations of antibiotics on fruit quality and antioxidant defense system in Huanglongbing infected Kinnow orchards. AMB Express 2019, 9, 147. [Google Scholar] [CrossRef]
- Abd Ghafar, M.F.; Prasad, K.N.; Weng, K.K.; Ismail, A. Flavonoid, hesperidine, total phenolic contents and antioxidant activities from Citrus species. Afr. J. Biotechnol. 2010, 9, 326–330. [Google Scholar]
- Rekha, C.; Poornima, G.; Manasa, M.; Abhipsa, V.; Devi, J.P.; Kumar, H.T.V.; Kekuda, T.R.P. Ascorbic acid, total phenol content and antioxidant activity of fresh juices of four ripe and unripe citrus fruits. Chem. Sci. Trans. 2012, 1, 303–310. [Google Scholar] [CrossRef]
- Chen, M.; Yang, D.; Liu, S. Effects of drying temperature on the flavonoid, phenolic acid and antioxidative capacities of the methanol extract of citrus fruit (Citrus sinensis (L.) Osbeck) peels. Int. J. Food Sci. Technol. 2011, 46, 1179–1185. [Google Scholar] [CrossRef]
- Tripoli, E.; La Guardia, M.; Giammanco, S.; Di Majo, D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Hijaz, F.M.; Manthey, J.A.; Folimonova, S.Y.; Davis, C.L.; Jones, S.E.; Reyes-De-Corcuera, J.I. An HPLC-MS characterization of the changes in sweet orange leaf metabolite profile following infection by the bacterial pathogen Candidatus Liberibacter asiaticus. PLoS ONE 2013, 8, e79485. [Google Scholar] [CrossRef] [PubMed]
- Bellés, J.M.; López-Gresa, M.P.; Fayos, J.; Pallás, V.; Rodrigo, I.; Conejero, V. Induction of cinnamate 4-hydroxylase and phenylpropanoids in virus-infected cucumber and melon plants. Plant Sci. 2008, 174, 524–533. [Google Scholar] [CrossRef]
- López-Gresa, M.P.; Torres, C.; Campos, L.; Lisón, P.; Rodrigo, I.; Bellés, J.M.; Conejero, V. Identification of defence metabolites in tomato plants infected by the bacterial pathogen Pseudomonas syringae. Environ. Exp. Bot. 2011, 74, 216–228. [Google Scholar] [CrossRef]
- Pietta, P.-G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Rao, M.J.; Ahmed, U.; Ahmed, M.H.; Duan, M.Z.; Wang, J.; Wang, Y.; Wang, L.Q. Comparison and quantification of metabolites and their antioxidant activities in the young and mature leaf of Sugarcane. ACS Food Sci. Technol. 2021, 3, 362–373. [Google Scholar] [CrossRef]
- Yu, J.; Wang, L.; Walzem, R.L.; Miller, E.G.; Pike, L.M.; Patil, B.S. Antioxidant activity of citrus limonoids, flavonoids, and coumarins. J. Agric. Food Chem. 2005, 53, 2009–2014. [Google Scholar] [CrossRef]
- Yoo, K.M.; Hwang, I.K.; Park, J.H.; Moon, B. Major phytochemical composition of 3 native Korean citrus varieties and bioactive activity on V79-4 cells induced by oxidative stress. J. Food Sci. 2009, 74, 462–468. [Google Scholar] [CrossRef]
- Kim, H.G.; Kim, G.; Park, S.; Lee, J.H.; Seo, O.N.; Lee, S.J.; Kim, J.H.; Shim, J.; Abd El-Aty, A.M.; Jin, J.S. Flavonoid profiling in three citrus varieties native to the Republic of Korea using liquid chromatography coupled with tandem mass spectrometry: Contribution to overall antioxidant activity. Biomed. Chromatogr. 2012, 26, 464–470. [Google Scholar] [CrossRef]
- Hirata, T.; Fujii, M.; Akita, K.; Yanaka, N.; Ogawa, K.; Kuroyanagi, M.; Hongo, D. Identification and physiological evaluation of the components from Citrus fruits as potential drugs for anti-corpulence and anticancer. Bioorg. Med. Chem. 2009, 17, 25–28. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Sood, S.; Muthuraman, A. In-vitro evaluation of bioactive compounds, anti-oxidant, lipid peroxidation and lipoxygenase inhibitory potential of Citrus karna L. peel extract. J. Food Sci. Technol. 2014, 51, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Solovchenko, A. Photoprotection in Plants: Optical Screening-Based Mechanisms; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Volume 14, ISBN 364213887X. [Google Scholar]
- Pourcel, L.; Routaboul, J.-M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, L.-L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.-B.; Hao, B.-H.; Lyon, M.P. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59. [Google Scholar] [CrossRef]
- Sir Elkhatim, K.A.; Elagib, R.A.A.; Hassan, A.B. Content of phenolic compounds and vitamin C and antioxidant activity in wasted parts of Sudanese citrus fruits. Food Sci. Nutr. 2018, 6, 1214–1219. [Google Scholar] [CrossRef]
- Chhikara, N.; Kour, R.; Jaglan, S.; Gupta, P.; Gat, Y.; Panghal, A. Citrus medica: Nutritional, phytochemical composition and health benefits—A review. Food Funct. 2018, 9, 1978–1992. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Azmat, R. Citrus: An ancient fruits of promise for health benefits. In Citrus-Health Benefits and Production Technology; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.; Sehrawat, N.; Singh, M.; Upadhyay, S.K.; Aggarwal, D.; Sharma, A.K. Cardioprotective and hepatoprotective potential of citrus flavonoid naringin: Current status and future perspectives for health benefits. Asian J. Biol. Life Sci. 2020, 9, 1. [Google Scholar] [CrossRef]
- Selvamuthukumaran, M.; Boobalan, M.S.; Shi, J. Bioactive components in citrus fruits and their health benefits. Phytochem. Citrus Appl. Funct. Foods 2017. [Google Scholar] [CrossRef]
- Benavente-Garcia, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef]
- Park, J.Y.; Shin, M.S.; Kim, S.N.; Kim, H.Y.; Kim, K.H.; Shin, K.S.; Kang, K.S. Polysaccharides from Korean Citrus hallabong peels inhibit angiogenesis and breast cancer cell migration. Int. J. Biol. Macromol. 2016, 85, 522–529. [Google Scholar] [CrossRef]
- Meneguzzo, F.; Ciriminna, R.; Zabini, F.; Pagliaro, M. Accelerated production of hesperidin-rich citrus pectin from waste citrus peel for prevention and therapy of COVID-19. 2020. [Google Scholar] [CrossRef]
- Musumeci, L.; Maugeri, A.; Cirmi, S.; Lombardo, G.E.; Russo, C.; Gangemi, S.; Calapai, G.; Navarra, M. Citrus fruits and their flavonoids in inflammatory bowel disease: An overview. Nat. Prod. Res. 2020, 34, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Shinde, D.B.; Renge, B.B.; Solanke, G.M.; Tewari, S. Enlargement immunity system citrus fruits (orange) with assessment of nutritional value and more health benefits a review. Stud. Indian Place Names 2020, 40, 4968–4982. [Google Scholar]
- Pragasam, S.J.; Rasool, M. Dietary component p-coumaric acid suppresses monosodium urate crystal-induced inflammation in rats. Inflamm. Res. 2013, 62, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Mahato, N.; Sharma, K.; Sinha, M.; Cho, M.H. Citrus waste derived nutra-/pharmaceuticals for health benefits: Current trends and future perspectives. J. Funct. Foods 2018, 40, 307–316. [Google Scholar] [CrossRef]
- Sharma, K.; Mahato, N.; Cho, M.H.; Lee, Y.R. Converting citrus wastes into value-added products: Economic and environmently friendly approaches. Nutrition 2017, 34, 29–46. [Google Scholar] [CrossRef]
- Shetty, S.B.; Mahin-Syed-Ismail, P.; Varghese, S.; Thomas-George, B.; Kandathil-Thajuraj, P.; Baby, D.; Haleem, S.; Sreedhar, S.; Devang-Divakar, D. Antimicrobial effects of Citrus sinensis peel extracts against dental caries bacteria: An in vitro study. J. Clin. Exp. Dent. 2016, 8, e71. [Google Scholar] [CrossRef] [Green Version]
- Gigi, M.K. Fungal inhibitory effect of Citrus Limon peel essential oil on Candida albicans. Dent. J. 2015, 84, 84–88. [Google Scholar]
- Fernández-Fernández, A.M.; Dellacassa, E.; Medrano-Fernandez, A.; Del Castillo, M.D. Citrus Waste Recovery for Sustainable Nutrition and Health. In Food Wastes By-Products Nutraceutical Heal. Potential; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 193–222. [Google Scholar]
- Andrade, W.R.; Pires, A.J.V.; de Jesus, M.O. Citrus Co-Products in Ruminants Feeds: A Review. Iran. J. Appl. Anim. Sci. 2020, 10, 191–202. [Google Scholar]
- Curk, F.; Ollitrault, F.; Garcia-Lor, A.; Luro, F.; Navarro, L.; Ollitrault, P. Phylogenetic origin of limes and lemons revealed by cytoplasmic and nuclear markers. Ann. Bot. 2016, 117, 565–583. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xu, Y.; Zhang, S.; Cao, L.; Huang, Y.; Cheng, J.; Wu, G.; Tian, S.; Chen, C.; Liu, Y. Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction. Nat. Genet. 2017, 49, 765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, M.J.; Wang, L.Q. CRISPR/Cas9 technology for improving agronomic traits and future prospective in agriculture. Planta 2021, 254, 68. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xu, Y.; Jiang, X.; Yu, H.; Jia, H.; Tan, C.; Hu, G.; Hu, Y.; Rao, M.J.; Deng, X.X.; et al. Genome of citrus rootstock and global DNA demethylation caused by hetero-grafting. Hortic. Res. 2021, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nat. Biotechnol. 2014, 32, 656. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ROS | Reacts with | Enzymatic Scavenging System | Metabolic Antioxidants | Reaction with ROS to Enhance Stress Tolerance |
|---|---|---|---|---|
| Superoxide (O•−2) | Fe–S proteins dismutate to H2O2 | SOD EC 1.15.1.1 | Proline/Glycine betaine | Helps in enhancing stress tolerance |
| Hydrogen peroxide (H2O2) | Proteins, heme-proteins, and DNA | CAT EC 1.11.1.6 GPX EC 1.11.1.9 GST EC 2.5.1.18 APX EC 1.11.1.11 | Amino acids, carotenoids, α-tocopherol/ascorbic acid, and glutathione | Hunts ROS |
| Singlet oxygen (1O2) | Oxidized lipids, G-residues of DNA, and proteins | - | Carotenoids and α-tocopherol (vitamin E) | Neutralizes free radicals and protects the photosynthetic apparatus from ROS |
| Hydroxyl radical (OH•) | DNA, RNA, lipids, and proteins | - | Flavonoids, sugars, proline Ascorbate. | Helps in maintaining cell homeostasis |
| Other reactive radicals | - | POD EC 1.11.1.x GR EC 1.6.4.2 | Fatty acids/organic acids and polyphenols (flavonoids) | Protect cells from negative effects of ROS by trapping free radicals |
| Serial No. | Genes | Identified in | Common Name | Category | Metabolism | Involved in | References |
|---|---|---|---|---|---|---|---|
| 1 | CrMYB68 | Citrus reticulata cv. Suavissima | Mandarin | R2R3-MYB transcription factor | Carotenoid metabolism | α- and β-branch carotenoids | [58] |
| 2 | UGT708G1 | Fortunella crassifolia | Kumquat | UGT-glucosyltransferase enzyme | Flavonoid accumulation | Anthocyanin biosynthesis | [59] |
| 3 | UGT708G2 | Citrus unshiu | Satsuma mandarin | UGT-glucosyltransferase enzyme | flavonoid accumulation | Anthocyanin pigments | [59] |
| 4 | CgMYB58 | Citrus maxima | Pummelo | MYB transcription factor | Lignin biosynthesis | Lignin accumulation in juice vesicles | [60] |
| 6 | Ruby and Noemi (bHLH) | Citrus sinensis, Citrus medica, and their hybrid | Orange, citron, and their hybrid | Transcription factor | Color formation | Flavonoid and anthocyanin biosynthesis | [61] |
| 7 | CsMYB3 and CsRuby1 | Citrus sinensis | Sweet orange | Transcription factor | Anthocyanin biosynthesis | Anthocyanin pigment accumulation | [62] |
| 8 | CCD4 | Citrus reticulata | Mandarin and its hybrids | CAROTENOID CLEAVAGE DIOXYGENASE | Carotenoid metabolism | [63] | |
| 9 | CsMADS6 | Citrus sinensis | Sweet orange | Transcription factor | Carotenoid metabolism | Activating downstream carotenoid genes | [64] |
| 10 | CsUGT78D3 | Citrus sinensis | Sweet orange | UDP-glucosyl transferase enzyme | Enhances proanthocyanidins and anthocyanins | High light stress tolerance by high anthocyanin contents | [65] |
| 11 | CsCYT75B1 | Citrus sinensis | Sweet orange | Cytochrome P450 75B1 enzyme | Flavonoid biosynthesis | Drought tolerance due to high flavonoid content | [19] |
| 12 | CWINVs, VINV, SPS2, SUT2, VPPs | Citrus sinensis (HAL) | Orange (Hong Anliu) | Genes encoding enzymes | Sugar | Sugar accumulation | [66] |
| 13 | CitLGT | Citrus unshiu Marc. | Satsuma mandarin | Limonoids UDP-glucosyl transferase enzyme | Limonoid GTase | Converting limonoid aglycones to glucosides | [67] |
| 14 | CpGTs | Citrus paradisi cv. Duncan | Grapefruit | Glucosyltransferases (GTs) | Color development | Color development | [68] |
| Citrus Species Common and Scientific Name | Antioxidant Capacity (µmol TE/100 g) | Total Phenolics (mg Gallic Acid Equivalent/g) | Total Amino Acids (g/100 g of Sample) | Total Carotenoids (mg/kg) | Total Flavonoids (mg/100 mL Juice) | Total Volatiles (1 Unit Equals to 10 mg/g Fresh Weight) | β-Carotene (mg/kg) | Lycopene (mg/kg) | Ascorbic Acid (mg/kg) | Total Acidity (g/100 mL Juice) |
|---|---|---|---|---|---|---|---|---|---|---|
| Robinson (Citrus reticulata) | 20.45 ± 0.98 | 209.37 ± 1.37 | - | 26.67 ± 0.67 | - | - | 22.67 ± 0.54 | 4.19 ± 0.12 | 651.33 ± 0.93 | 0.563 |
| Clementine (Citrus reticulata) | 33.10 ± 0.68 | 302.38 ± 0.91 | 25.54 ± 0.22 | 27.23 ± 0.12 | 19.23 ± 0.97 | 191.23 ± 1.29 | 22.33 ± 0.13 | 3.27 ± 0.20 | 656.43 ± 1.03 | 0.588 |
| Cocktail (Citrus paradisi) | 45.28 ± 0.76 | 214.88 ± 0.87 | - | 37.40 ± 0.33 | - | 155.4 ± 2.51 | 31.79 ± 0.93 | 3.20 ± 0.07 | 353.17 ± 0.77 | |
| Valencia (Citrus sinensis) | 40.32 ± 1.01 | 270.56 ± 0.67 | 20.56 ± 0.31 | 29.87 ± 0.98 | 18.34 ± 1.22 | 289.43 ± 4.81 | 25.89 ± 0.36 | 2.09 ± 0.24 | 579.99 ± 1.10 | 1.024 |
| Wild lime (Citrus hystrix) | 83.91 ± 0.81 | 490.74 ± 1.75 | - | - | 22.25 ± 0.20 | - | - | - | - | - |
| Common lime (Citrus aurantifolia) | 69.54 ± 0.58 | 211.70 ± 0.0 | - | - | 10.67 ± 0.27 | 512.92 ± 2.19 | - | - | - | 3.328 |
| Citrus maxima | 29.34 ± 1.04 | 501.43 ± 2.98 | 23.9 ± 0.9 | 23.17 ± 1.67 | 19.45 ± 0.65 | - | - | - | - | - |
| Atalantia buxifolia | 74.24 ± 2.19 | 645.89 ± 3.47 | 24.84 ± 0.16 | 27.83 ± 1.45 | 28.53 ± 1.24 | 1567.11 ± 3.82 | - | - | - | - |
| Poncirus trifoliata | - | - | - | - | - | 145. 78 ± 1.35 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, M.J.; Wu, S.; Duan, M.; Wang, L. Antioxidant Metabolites in Primitive, Wild, and Cultivated Citrus and Their Role in Stress Tolerance. Molecules 2021, 26, 5801. https://doi.org/10.3390/molecules26195801
Rao MJ, Wu S, Duan M, Wang L. Antioxidant Metabolites in Primitive, Wild, and Cultivated Citrus and Their Role in Stress Tolerance. Molecules. 2021; 26(19):5801. https://doi.org/10.3390/molecules26195801
Chicago/Turabian StyleRao, Muhammad Junaid, Songguo Wu, Mingzheng Duan, and Lingqiang Wang. 2021. "Antioxidant Metabolites in Primitive, Wild, and Cultivated Citrus and Their Role in Stress Tolerance" Molecules 26, no. 19: 5801. https://doi.org/10.3390/molecules26195801
APA StyleRao, M. J., Wu, S., Duan, M., & Wang, L. (2021). Antioxidant Metabolites in Primitive, Wild, and Cultivated Citrus and Their Role in Stress Tolerance. Molecules, 26(19), 5801. https://doi.org/10.3390/molecules26195801