
Composition of Algerian Propolis, Plant Origin, and Its Antiangiogenic Activity In Vitro


Abstract
:1. Introduction
2. Results
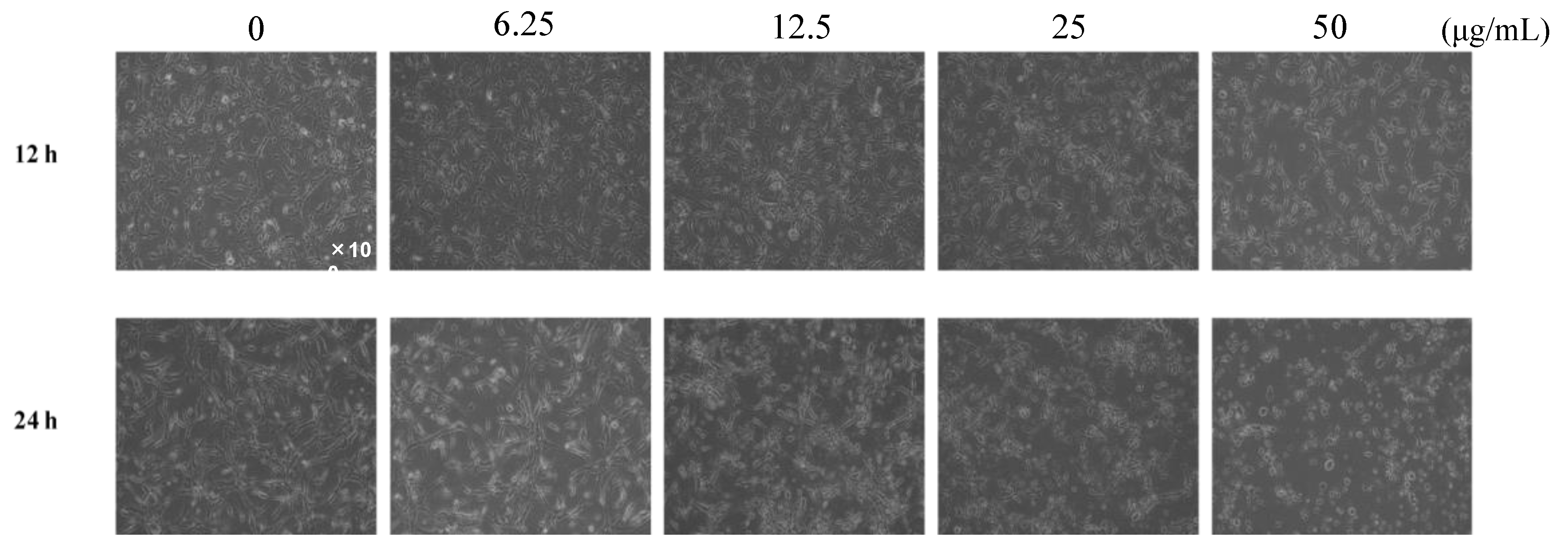
2.1. Investigation of the Anti-Tube-Forming Activity of Algerian Propolis
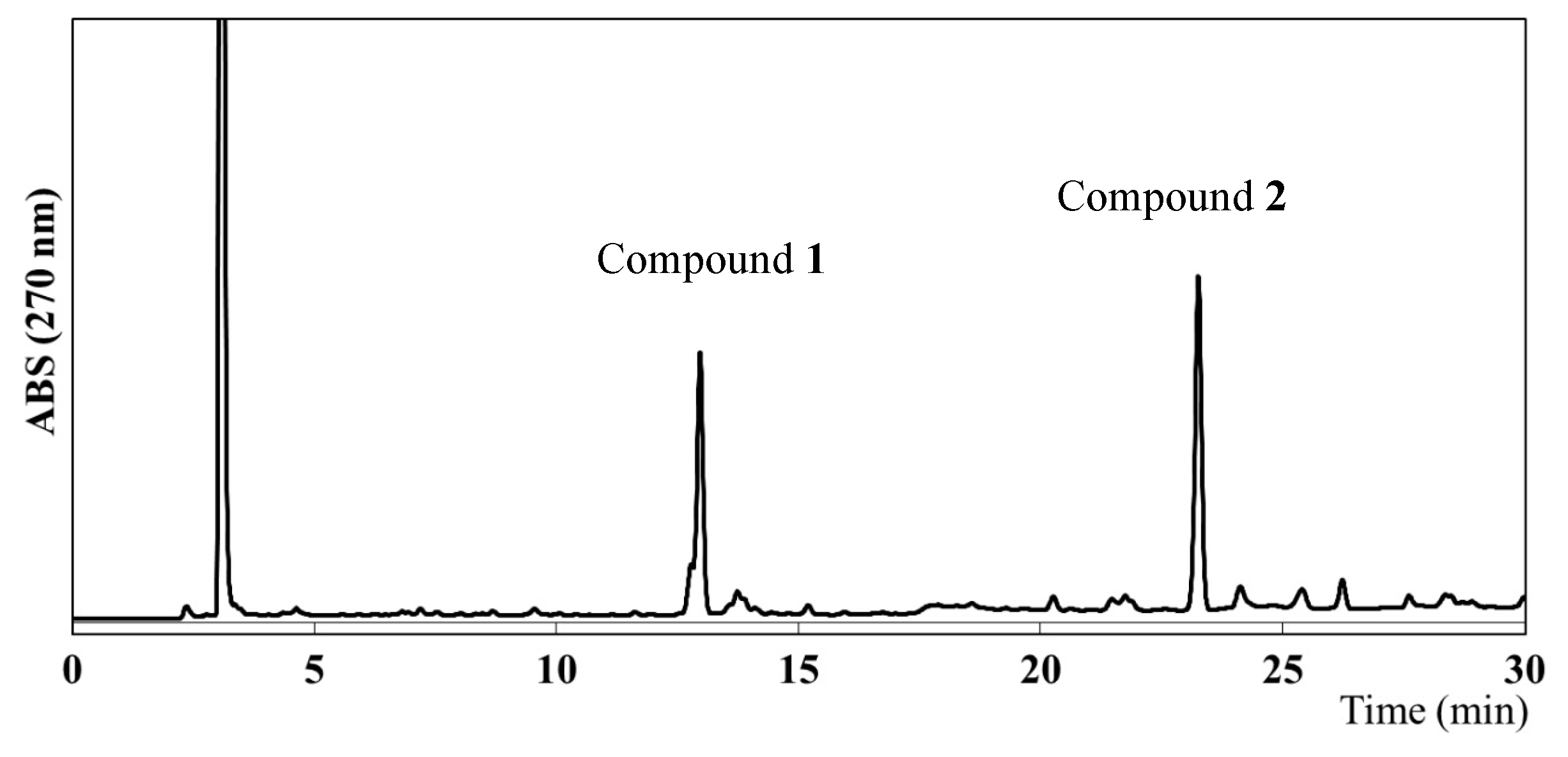
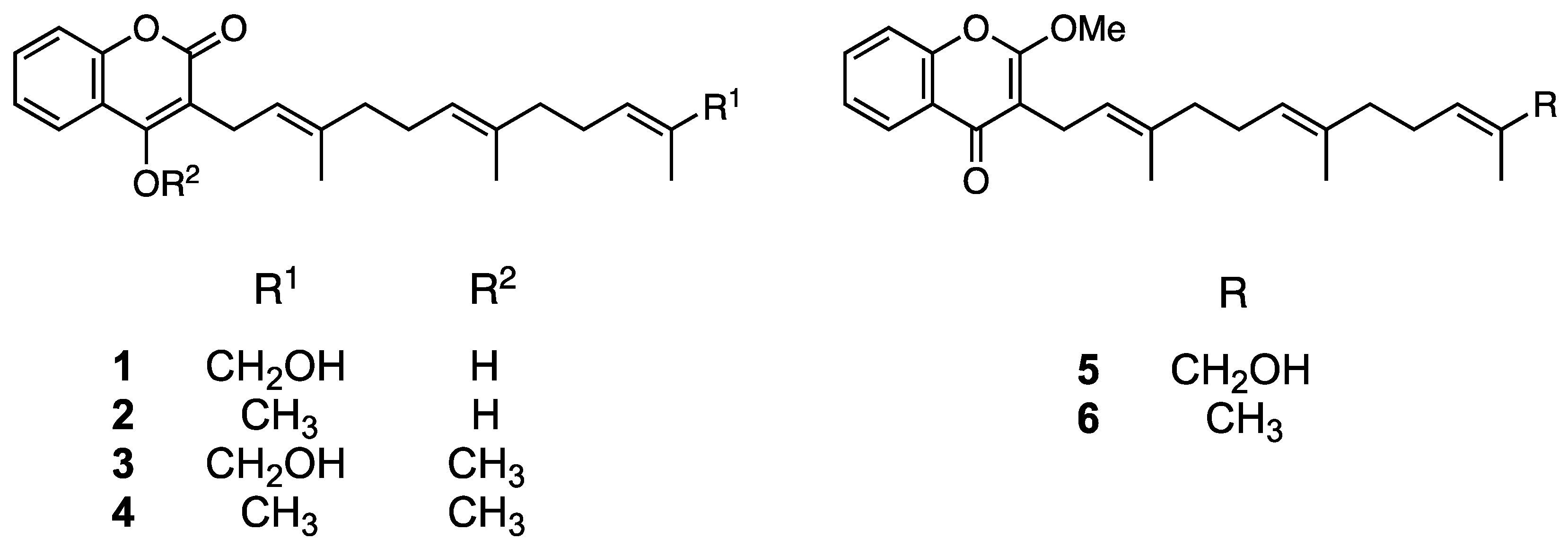
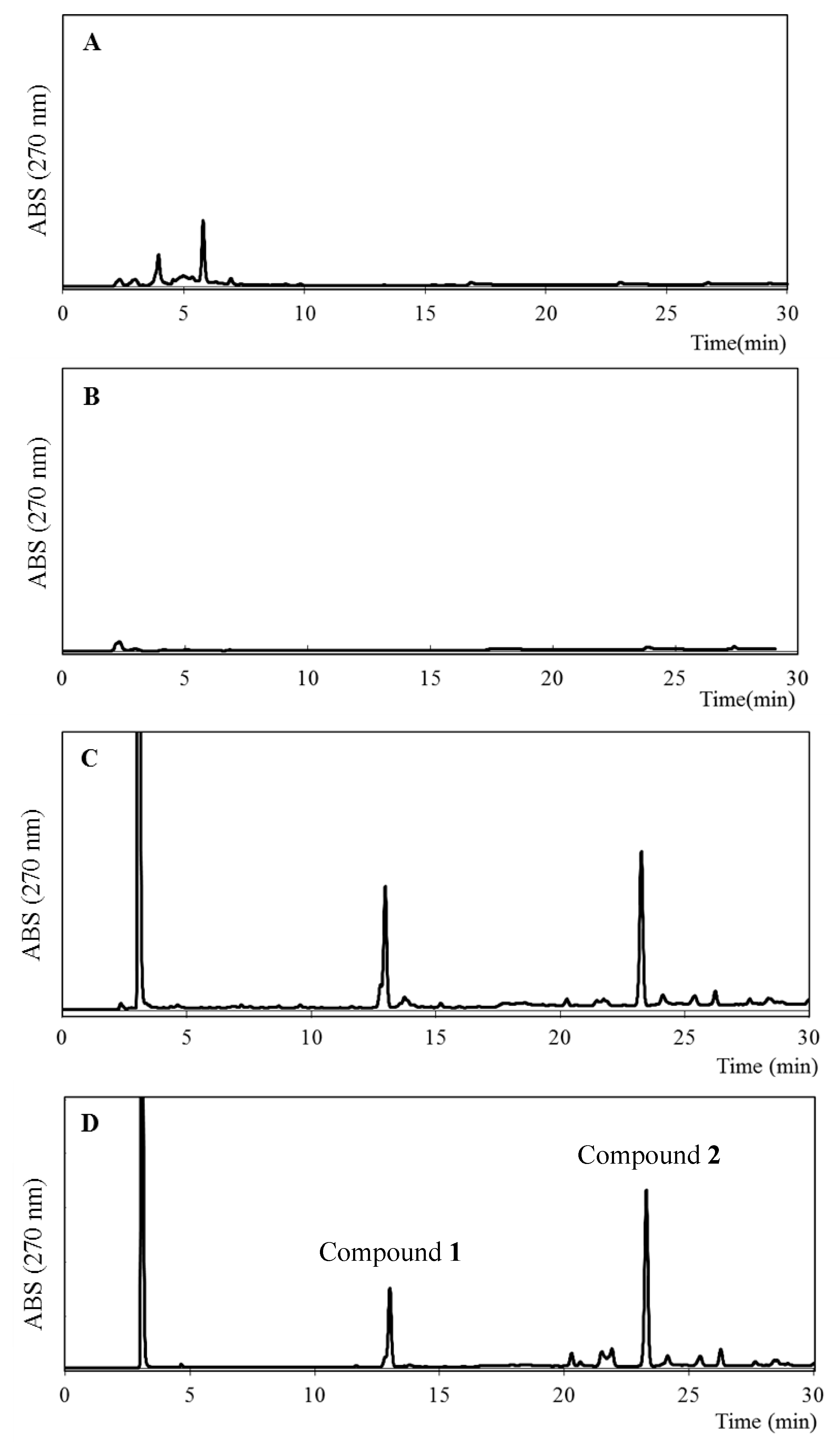
2.2. Identification of the Main Compounds in EEAP1
2.3. Anti-Tube-Forming Activity of ω-Hydroxyferulenol (1) and Ferulenol (2)
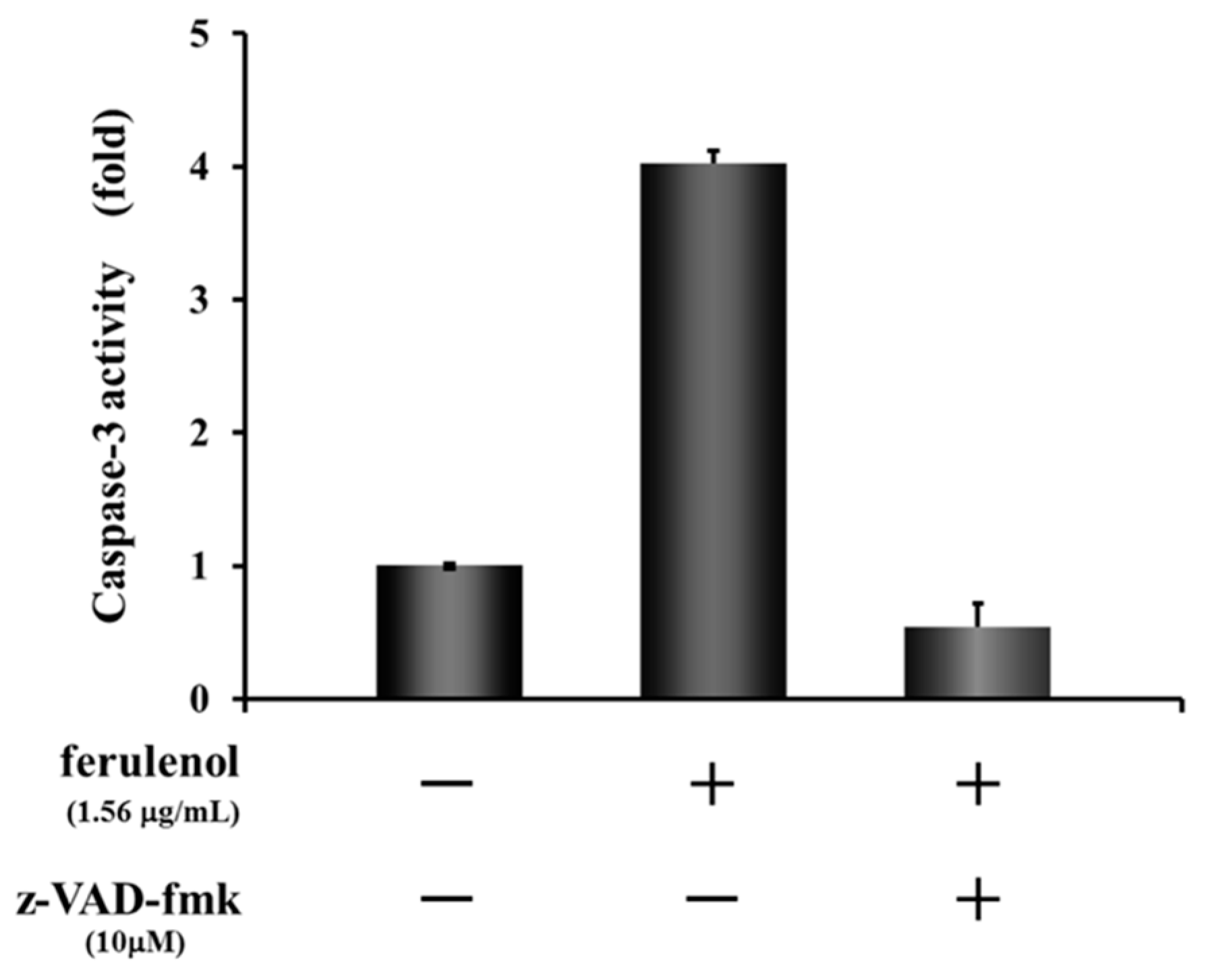
2.4. Apoptosis Induction of Ferulenol (2)
2.5. Identification of the Plant Origin of the Studied Propolis
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Equipment for the Isolation and Structure Analysis
4.3. Propolis Samples
4.4. Screening of the Four Algerian Propolis Samples and Isolation of Active Compounds (1 and 2)
4.5. Cell Culture
4.6. Tube Formation Assay
4.7. Caspase-Glo 3/7 Assay
4.8. Sampling of the Resin of Ferula Communis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| EEAP | ethanol extracts of Algerian propolis |
| HPLC | high-performance liquid chromatography |
| HUVECs | human umbilical vein endothelial cells |
| NMR | nuclear magnetic resonance |
| HR-ESIMS | high-resolution electrospray ionization mass spectroscopy |
| TMS | trimethylsilyl |
References
- Ghisalberti, E.L. Propolis: A review. Bee World 1979, 60, 59–84. [Google Scholar] [CrossRef]
- Bankova, V.S.; de Castro, S.L.; Marcucci, M.C. Propolis: Recent advances in chemistry and plant origin. Apidologie 2000, 31, 3–15. [Google Scholar] [CrossRef]
- Bankova, V. Chemical diversity of propolis makes it a valuable source of new biological active compounds. J. ApiProd. ApiMed. Sci. 2009, 1, 23–28. [Google Scholar] [CrossRef]
- Sforcin, J.M.; Bankova, V. Propolis: Is there a potential for the development of new drugs? J. Ethnopharmacol. 2011, 133, 253–260. [Google Scholar] [CrossRef]
- Sforcin, J.M. Biological properties and therapeutic applications of propolis. Phytother. Res. 2016, 30, 894–905. [Google Scholar] [CrossRef]
- Oryan, A.; Alemzadeh, E.; Moshiri, A. Potential role of propolis in wound healing: Biological properties and therapeutic activities. Biomed. Pharmacother. 2018, 98, 469–483. [Google Scholar] [CrossRef]
- Hamasaka, T.; Kumazawa, S.; Fujimoto, T.; Nakayama, T. Antioxidant activity and constituents of propolis collected in various areas of Japan. Food Sci. Technol. Res. 2004, 10, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant potential of propolis, bee pollen, and royal jelly: Possible medical application. Oxid. Med. Cell. Longev. 2018, 2018, 7074209. [Google Scholar] [CrossRef]
- Inui, S.; Hosoya, T.; Shimamura, Y.; Masuda, S.; Ogawa, T.; Kobayashi, H.; Shirafuji, K.; Moli, R.T.; Kozone, I.; Shin-ya, K.; et al. Solophenols B–D and solomonin: New prenylated polyphenols isolated from propolis collected from the Solomon Islands and their antibacterial activity. J. Agric. Food Chem. 2012, 60, 11765–11770. [Google Scholar] [CrossRef]
- Przybyłek, I.; Karpiński, T.M. Antibacterial properties of propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchin, M.; Freires, I.A.; Lazarini, J.G.; Nani, B.D.; da Cunha, M.G.; Colón, D.F.; de Alencar, S.M.; Rosalen, P.L. The use of Brazilian propolis for discovery and development of novel anti-inflammatory drugs. Eur. J. Med. Chem. 2018, 153, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Inui, S.; Hosoya, T.; Yoshizumi, K.; Sato, H.; Kumazawa, S. Phytochemical and anti-inflammatory properties of Senegalease propolis. Fitoterapia 2021, 151, 104861. [Google Scholar] [CrossRef]
- Ahn, M.R.; Kunimasa, K.; Ohta, T.; Kumazawa, S.; Kamihira, M.; Kaji, K.; Uto, Y.; Hori, H.; Nagasawa, H.; Nakayama, T. Suppression of tumor-induced angiogenesis by Brazilian propolis: Major component artepillin C inhibits in vitro tube formation and endothelial cell proliferation. Cancer Lett. 2007, 252, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, C.; Núñez, O.; Torres, A.; Guzmán, L.; Wehinger, S. Antitumor activity of propolis: Recent advances in cellular perspectives, animal models and possible applications. Food Rev. Int. 2020, 36, 429–455. [Google Scholar] [CrossRef]
- Pasupuleti, V.R.; Sammugam, L.; Ramesh, N.; Gan, S.H. Honey, propolis, and royal jelly: A comprehensive review of their biological actions and health benefits. Oxid. Med. Cell. Longev. 2017, 2017, 1259510. [Google Scholar] [CrossRef]
- Salatino, A.; Fernandes-Silva, C.C.; Righi, A.A.; Salatino, M.L.F. Propolis research and the chemistry of plant products. Nat. Prod. Rep. 2011, 28, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Bankova, V.; Popova, M.; Trusheva, B. The phytochemistry of the honeybee. Phytochemistry 2018, 155, 1–11. [Google Scholar] [CrossRef]
- Kumazawa, S.; Hamasaka, T.; Nakayama, T. Antioxidant activity of propolis of various geographic origins. Food Chem. 2004, 84, 329–339. [Google Scholar] [CrossRef]
- Kumazawa, S.; Yoneda, M.; Shibata, I.; Kanaeda, J.; Hamasaka, T.; Nakayama, T. Direct evidence for the plant origin of Brazilian propolis by the observation of honeybee behavior and phytochemical analysis. Chem. Pharm. Bull. 2003, 51, 740–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inui, S.; Hatano, A.; Yoshino, M.; Hosoya, T.; Shimamura, Y.; Masuda, S.; Ahn, M.R.; Tazawa, S.; Araki, Y.; Kumazawa, S. Identification of the phenolic compounds contributing to antibacterial activity in ethanol extracts of Brazilian red propolis. Nat. Prod. Res. 2014, 28, 1293–1296. [Google Scholar] [CrossRef]
- Ccana-Ccapatinta, G.V.; Mejía, J.A.A.; Tanimoto, M.H.; Groppo, M.; de Carvalho, J.C.A.S.; Bastos, J.K. Dalbergia ecastaphyllum (L.) Taub. and Symphonia globulifera L.f.: The botanical sources of isoflavonoids and benzophenones in Brazilian red propolis. Molecules 2020, 25, 2060. [Google Scholar] [CrossRef] [PubMed]
- Kumazawa, S.; Nakamura, J.; Murase, M.; Miyagawa, M.; Ahn, M.-R.; Fukumoto, S. Plant origin of Okinawan propolis: Honeybee behavior observation and phytochemical analysis. Naturwissenschaften 2008, 95, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, K.; Inui, S.; Sugiyama, Y.; Kurosawa, M.; Nakamura, J.; Choi, S.J.; Ahn, M.-R.; Kumazawa, S. Identification of the plant origin of propolis from Jeju Island, Korea, by observation of honeybee behavior and phytochemical analysis. Biosci. Biotechnol. Biochem. 2012, 76, 2135–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, M.-R.; Kumazawa, S.; Hamasaka, T.; Bang, K.S.; Nakayama, T. Antioxidant activity and constituents of propolis collected in various areas of Korea. J. Agric. Food Chem. 2004, 52, 7286–7292. [Google Scholar] [CrossRef]
- Brihoum, H.; Maiza, M.; Sahali, H.; Boulmeltoiu, M.; Barratt, G.; Benguedouar, L.; Lahouel, M. Dual effect of Algerian propolis on lung cancer: Antitumor and chemopreventive effects involving antioxidant activity. Braz. J. Pharm. Sci. 2018, 54, e17396. [Google Scholar] [CrossRef]
- Bedaida, I.K.; Masry, S.H.D.; Mamache, B.; Shehata, M.G.; Benammar, L.; Ayachi, A. Ethanolic extract of Algerian propolis induced cell damage in Staphylococcus aureus: A promising alternative as a natural biopreservative in food products. Acta Aliment. 2020, 49, 505–514. [Google Scholar] [CrossRef]
- Segueni, N.; Keskin, M.; Keskin, Ş.; Kadour, B.; Kolayli, S.; Salah, A. Comparison between phenolic content, antioxidant, and antibacterial activity of Algerian and Turkish propolis. Comb. Chem. High Throughput Screen. 2021, 24, 1679–1687. [Google Scholar] [CrossRef]
- Tsuchiya, I.; Hosoya, T.; Ushida, M.; Kunimasa, K.; Ohta, T.; Kumazawa, S. Nymphaeol-A isolated from Okinawan propolis suppresses angiogenesis and induces caspase-dependent apoptosis via inactivation of survival signals. Evid. Based Complementary Alternat. Med. 2013, 2013, 826245. [Google Scholar] [CrossRef] [Green Version]
- Gordon, M.S.; Mendelson, D.S.; Kato, G. Tumor angiogenesis and novel antiangiogenic strategies. Int. J. Cancer 2010, 126, 1777–1787. [Google Scholar] [CrossRef]
- Abbaszadeh, H.; Keikhaei, B.; Mottaghi, S. A review of molecular mechanisms involved in anticancer and antiangiogenic effects of natural polyphenolic compounds. Phytother. Res. 2019, 33, 2002–2014. [Google Scholar] [CrossRef]
- Valle, M.G.; Appendino, G.; Nano, G.M.; Picci, V. Prenylated coumarins and sesquiterpenoids from Ferula communis. Phytochemistry 1986, 26, 253–256. [Google Scholar] [CrossRef]
- Appendino, G.; Mercalli, E.; Fuzzati, N.; Arnoldi, L.; Stavri, M.; Gibbons, S.; Ballero, M.; Maxia, A. Antimycobacterial coumarins from the sardinian giant fennel (Ferula communis). J. Nat. Prod. 2004, 67, 2108–2110. [Google Scholar] [CrossRef] [PubMed]
- Malika, B.; Amar, Z.; Lamia, B.; Gillian, B.; Mesbah, L. Cytotoxicity of sesquiterpenes ferulenol and coladin on liver FAO and B16F melanoma cells. Pharmacogen. Mag. 2018, 14, 333–337. [Google Scholar]
- Bocca, C.; Gabriel, L.; Bozzo, F.; Miglietta, A. Microtubule-interacting activity and cytotoxitity of the prenylated coumarin ferulenol. Planta Med. 2002, 68, 1135–1137. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Voucher Specimen | Area | Color and Texture |
|---|---|---|---|
| 1 | TIA-1 | Tiaret | Dark brown and rigid |
| 2 | TIA-2 | Tiaret | Dark brown and rigid |
| 3 | NED-TL | Tlemcen | Dark brown and waxy |
| 4 | SFS-SBA | Sidi bel Abbés | Brown, waxy, and sticky |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosoya, T.; Tsuchiya, I.; Ohta, T.; Benhanifia, M.; Kumazawa, S. Composition of Algerian Propolis, Plant Origin, and Its Antiangiogenic Activity In Vitro. Molecules 2021, 26, 6510. https://doi.org/10.3390/molecules26216510
Hosoya T, Tsuchiya I, Ohta T, Benhanifia M, Kumazawa S. Composition of Algerian Propolis, Plant Origin, and Its Antiangiogenic Activity In Vitro. Molecules. 2021; 26(21):6510. https://doi.org/10.3390/molecules26216510
Chicago/Turabian StyleHosoya, Takahiro, Ikumi Tsuchiya, Toshiro Ohta, Mokhtar Benhanifia, and Shigenori Kumazawa. 2021. "Composition of Algerian Propolis, Plant Origin, and Its Antiangiogenic Activity In Vitro" Molecules 26, no. 21: 6510. https://doi.org/10.3390/molecules26216510
APA StyleHosoya, T., Tsuchiya, I., Ohta, T., Benhanifia, M., & Kumazawa, S. (2021). Composition of Algerian Propolis, Plant Origin, and Its Antiangiogenic Activity In Vitro. Molecules, 26(21), 6510. https://doi.org/10.3390/molecules26216510