
The Methionine 549 and Leucine 552 Residues of Friedelin Synthase from Maytenus ilicifolia Are Important for Substrate Binding Specificity

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Determination of Friedelin Synthase Mutants
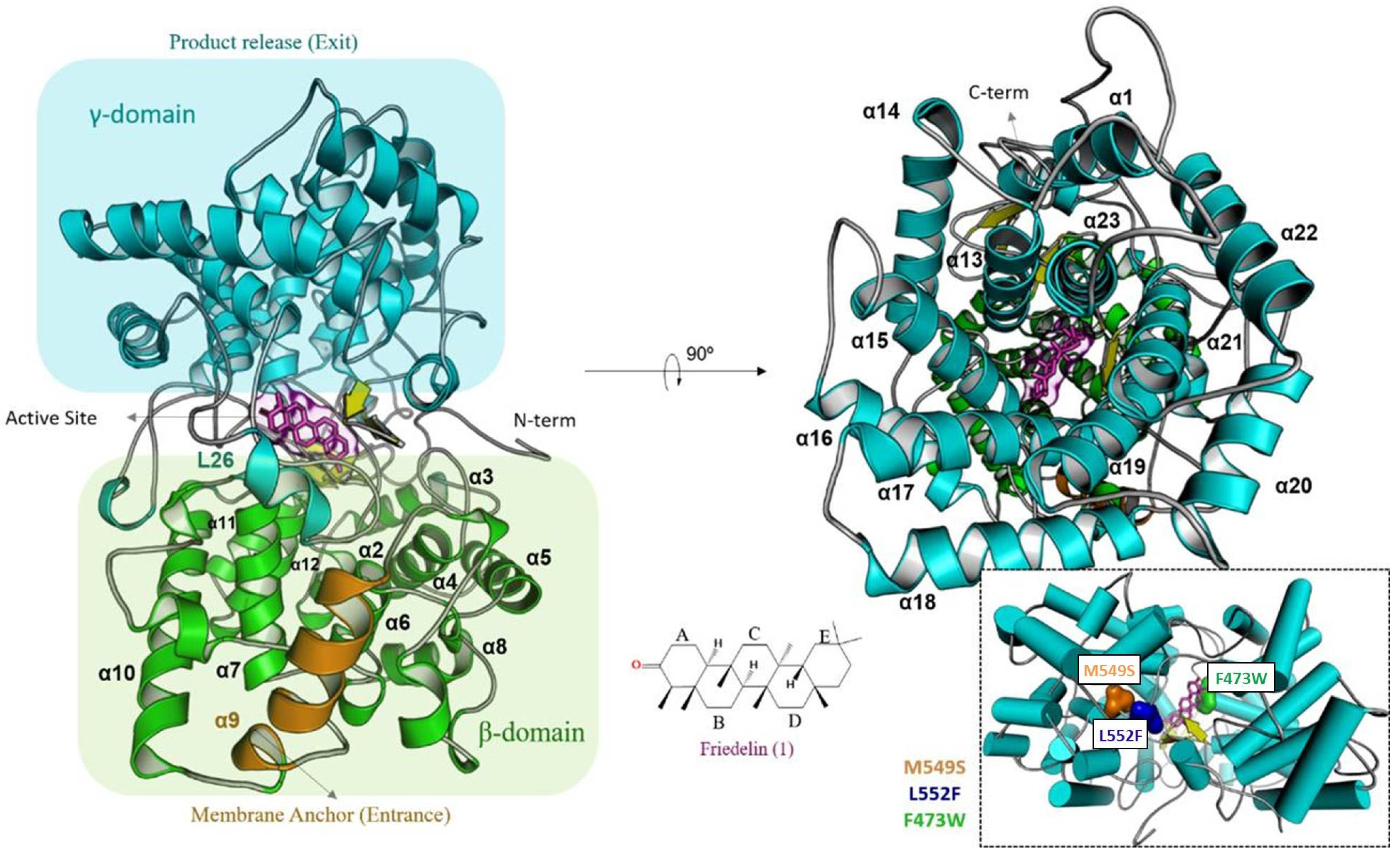
2.1.1. Phe473 Is Essential for Friedelin Activity
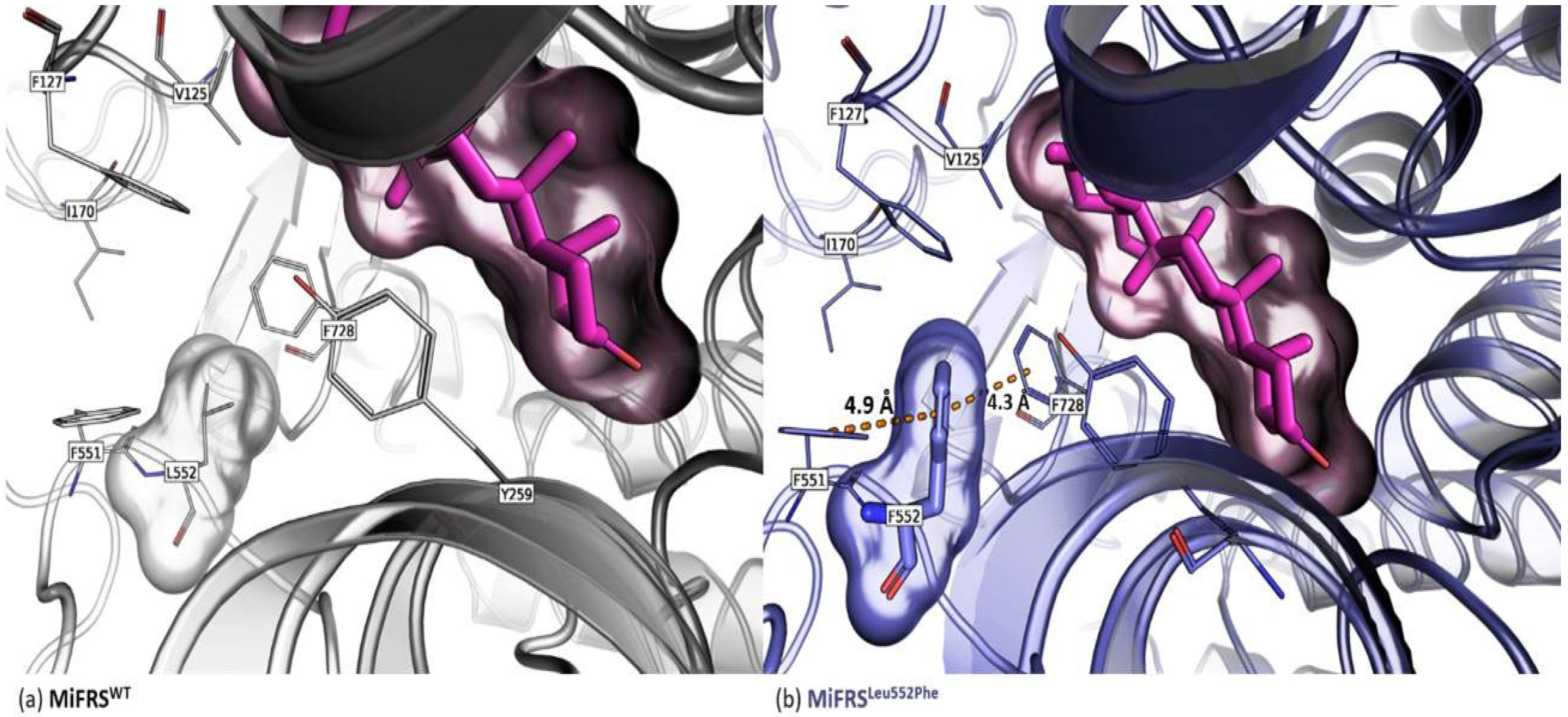
2.1.2. Met549 and Leu552 Influence the Specificity of MiFRS
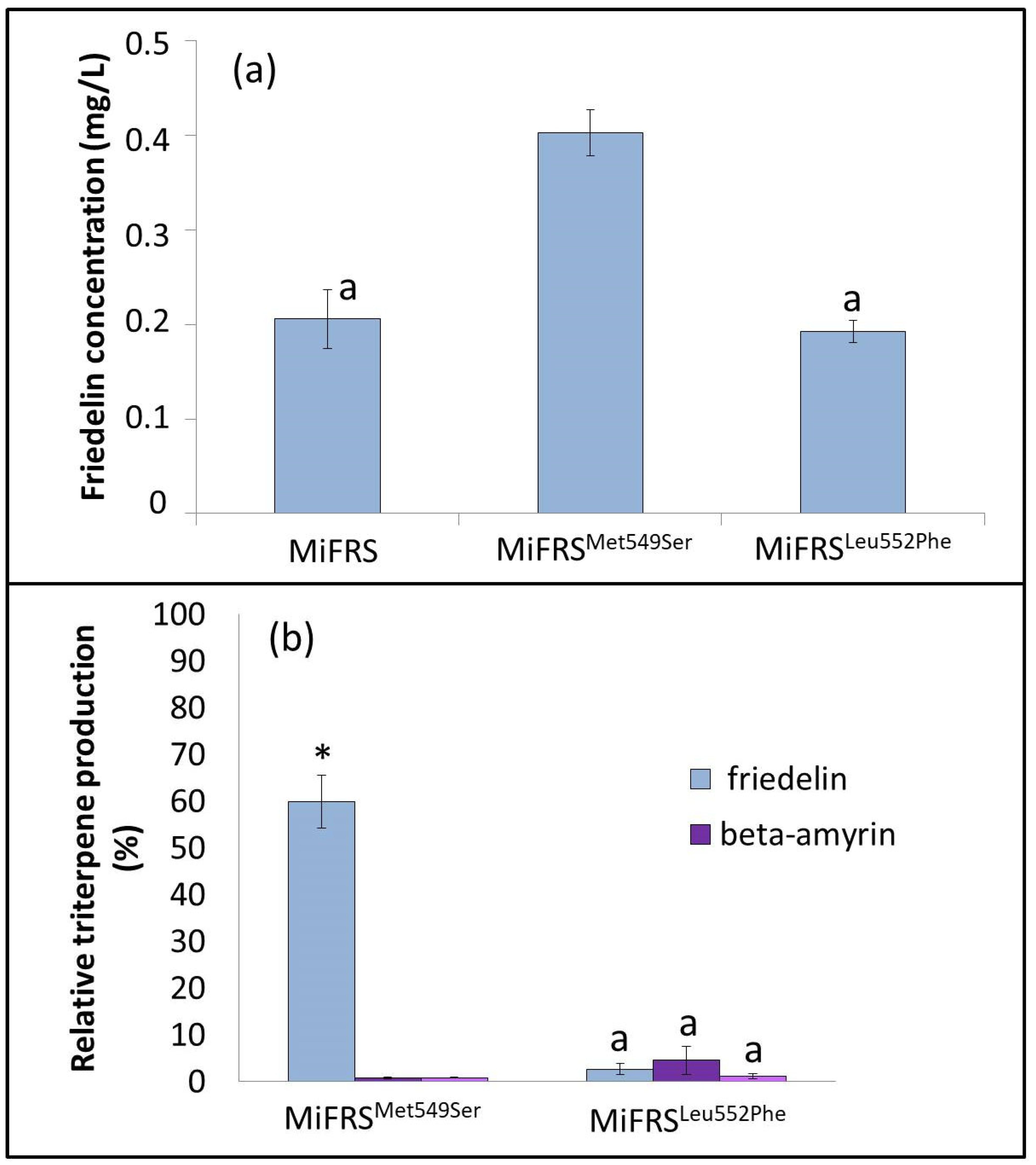
2.1.3. Met549Ser Is More Efficient in Friedelin Production
3. Discussion
4. Materials and Methods
4.1. Site-Directed Mutagenesis Reaction
4.2. Heterologous Expression of MiFRS Mutants in S. cerevisiae
4.3. Extraction of Biosynthesized Substances by the Heterologous System
4.4. Analysis of the Generated Products
4.5. Protein Homology Modelling
4.6. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Withers, S.T.; Keasling, J.D. Biosynthesis and engineering of isoprenoid small molecules. Appl. Microbiol. Biotechnol. 2007, 73, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.-A. Plant sterols and the membrane environment. Trends Plant Sci. 1998, 3, 170–175. [Google Scholar] [CrossRef]
- Phillips, D.R.; Rasbery, J.M.; Bartel, B.; Matsuda, S.P. Biosynthetic diversity in plant triterpene cyclization. Curr. Opin. Plant Biol. 2006, 9, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, K.; Suzuki, M.; Kikuchi, J.; Saito, K.; Muranaka, T. Dual biosynthetic pathways to phytosterol via cycloartenol and lanosterol in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Yeats, T.; Han, H.; Jetter, R. Cloning and characterization of oxidosqualene cyclases from Kalanchoe daigremontiana. J. Biol. Chem. 2019, 285, 29703–29712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene Biosynthesis in Plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef]
- Lodeiro, S.; Xiong, Q.; Wilson, W.K.; Kolesnikova, M.D.; Onak, C.S.; Matsuda, S.P.T. An Oxidosqualene Cyclase Makes Numerous Products by Diverse Mechanisms: A Challenge to Prevailing Concepts of Triterpene Biosynthesis. J. Am. Chem. Soc. 2007, 129, 11213–11222. [Google Scholar] [CrossRef]
- Gas-Pascual, E.; Berna, A.; Bach, T.J.; Schaller, H. Plant Oxidosqualene Metabolism: Cycloartenol Synthase—Dependent Sterol Biosynthesis in Nicotiana benthamiana. PLoS ONE 2014, 9, e109156. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Duan, L.; Liu, D.; Guo, J.; Ge, S.; Dicks, J.; Ómáille, P.; Osbourn, A.; Qi, X. Divergent evolution of oxidosqualene cyclases in plants. New Phytol. 2011, 193, 1022–1038. [Google Scholar] [CrossRef]
- Han, J.Y.; Ahn, C.-H.; Adhikari, P.; Kondeti, S.; Choi, Y.E. Functional characterization of an oxidosqualene cyclase (PdFRS) encoding a monofunctional friedelin synthase in Populus davidiana. Planta 2018, 249, 95–111. [Google Scholar] [CrossRef]
- Bicalho, K.U.; Santoni, M.M.; Arendt, P.; Zanelli, C.F.; Furlan, M.; Goossens, A.; Pollier, J. CYP712K4 Catalyzes the C-29 Oxidation of Friedelin in the Maytenus ilicifolia Quinone Methide Triterpenoid Biosynthesis Pathway. Plant Cell Physiol. 2019, 60, 2510–2522. [Google Scholar] [CrossRef]
- Corsino, J.; de Carvalho, P.R.F.; Kato, M.; Latorre, L.R.; Oliveira, O.M.M.; Araujo, A.R.; Bolzani, V.; Franca, S.; Pereira, A.M.S.; Furlan, M. Biosynthesis of friedelane and quinonemethide triterpenoids is compartmentalized in Maytenus aquifolium and Salacia campestris. Phytochemistry 2000, 55, 741–748. [Google Scholar] [CrossRef]
- Tabach, R.; Duarte-Almeida, J.M.; Carlini, E. Pharmacological and toxicological study of Maytenus ilicifolia leaf extract. Part I—Preclinical studies. Phytother. Res. 2017, 31, 921–926. [Google Scholar] [CrossRef]
- Antonisamy, P.; Duraipandiyan, V.; Ignacimuthu, S. Anti-inflammatory, analgesic and antipyretic effects of friedelin isolated from Azima tetracantha Lam. in mouse and rat models. J. Pharm. Pharmacol. 2011, 63, 1070–1077. [Google Scholar] [CrossRef]
- Antonisamy, P.; Duraipandiyan, V.; Aravinthan, A.; Al-Dhabi, N.A.; Ignacimuthu, S.; Choi, K.C.; Kim, J.-H. Protective effects of friedelin isolated from Azima tetracantha Lam. against ethanol-induced gastric ulcer in rats and possible underlying mechanisms. Eur. J. Pharmacol. 2015, 750, 167–175. [Google Scholar] [CrossRef]
- Christopher, R.; Nyandoro, S.S.; Chacha, M.; Koning, C.B. A new cinnamoylglycoflavonoid, antimycobacterial and antioxidant constituents from Heritiera littoralis leaf extracts. Nat. Prod. Res. 2014, 28, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Mann, A.; Ibrahim, K.; Oyewale, A.O.; Amupitan, J.O.; Fatope, M.O.; Okogun, J.I. Antimycobacterial Friedelane-terpenoid from the Root Bark of Terminalia avicennioides. Am. J. Chem. 2011, 1, 52–55. [Google Scholar] [CrossRef] [Green Version]
- Duraipandiyan, V.; Al-Dhabi, N.A.; Irudayaraj, S.S.; Sunil, C. Hypolipidemic activity of friedelin isolated from Azima tetracantha in hyperlipidemic rats. Rev. Bras. Farm. 2016, 26, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Souza-Moreira, T.M.; Alves, T.B.; Pinheiro, K.; Felippe, L.G.; De Lima, G.M.A.; Watanabe, T.F.; Barbosa, C.C.; Santos, V.A.F.F.M.; Lopes, N.P.; Valentini, S.R.; et al. Friedelin Synthase from Maytenus ilicifolia: Leucine 482 Plays an Essential Role in the Production of the Most Rearranged Pentacyclic Triterpene. Sci. Rep. 2016, 6, 36858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, R.; Hashimoto, I.; Masukawa, Y.; Hoshino, T. Effect of Cation-π Interactions and Steric Bulk on the Catalytic Action of Oxidosqualene Cyclase: A Case Study of Phe728 of β-Amyrin Synthase fromEuphorbia tirucalli L. Chem. Eur. J. 2013, 19, 17150–17158. [Google Scholar] [CrossRef] [PubMed]
- Hart, E.A.; Hua, L.; Darr, L.B.; Wilson, W.K.; Pang, A.J.; Matsuda, S.P.T. Directed Evolution To Investigate Steric Control of Enzymatic Oxidosqualene Cyclization. An Isoleucine-to-Valine Mutation in Cycloartenol Synthase Allows Lanosterol and Parkeol Biosynthesis. J. Am. Chem. Soc. 1999, 121, 9887–9888. [Google Scholar] [CrossRef]
- Salmon, M.; Thimmappa, R.B.; Minto, R.; Melton, R.; Hughes, R.K.; O’Maille, P.E.; Hemmings, A.; Osbourn, A. A conserved amino acid residue for product and substrate specificity in plant triterpene synthases. Proc. Natl. Acad. Sci. USA 2016, 113, E4407–E4414. [Google Scholar] [CrossRef] [Green Version]
- Tetsuo, K.M.S.; Kazuo, M.Y.E. Mutational studies on triterpene synthases: Engineering lupeol synthase into β-amyrin synthase. J. Am. Chem. Soc. 2000, 122, 6816–6824. [Google Scholar]
- Zhou, J.; Hu, T.; Gao, L.; Su, P.; Zhang, Y.; Zhao, Y.; Chen, S.; Tu, L.; Song, Y.; Wang, X.; et al. Friedelane-type triterpene cyclase in celastrol biosynthesis from Tripterygium wilfordii and its application for triterpenes biosynthesis in yeast. New Phytol. 2019, 223, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Masukawa, Y.; Nakada, C.; Amari, K.; Nakano, C.; Hoshino, T. β-Amyrin synthase from Euphorbia tirucalli. Steric bulk, not the π-electrons of Phe, at position 474 has a key role in affording the correct folding of the substrate to complete the normal polycyclization cascade. Org. Biomol. Chem. 2014, 12, 3836–3846. [Google Scholar] [CrossRef] [PubMed]
- Anjana, R.; Vaishnavi, M.K.; Sherlin, D.; Kumar, S.P.; Naveen, K.; Kanth, P.S.; Sekar, K. Aromatic-aromatic interactions in structures of proteins and protein-DNA complexes: A study based on orientation and distance. Bioinformation 2012, 8, 1220–1224. [Google Scholar] [CrossRef]
- Edelheit, O.; Hanukoglu, A.; Hanukoglu, I. Simple and efficient site-directed mutagenesis using two single-primer reactions in parallel to generate mutants for protein structure-function studies. BMC Biotechnol. 2009, 9, e61. [Google Scholar] [CrossRef] [Green Version]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Souza-Moreira, T.M.; Navarrete, C.; Chen, X.; Zanelli, C.; Valentini, S.R.; Furlan, M.; Nielsen, J.; Krivoruchko, A. Screening of 2A peptides for polycistronic gene expression in yeast. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.S.; Srinivasan, N.; Sowdhamini, R.; Blundell, T.L. Knowledge-Based Protein Modeling. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 1–68. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzeu, B.F.; Souza-Moreira, T.M.; Oliveira, A.A.; Remlinger, M.; Felippe, L.G.; Valentini, S.R.; Guido, R.V.C.; Zanelli, C.F.; Furlan, M. The Methionine 549 and Leucine 552 Residues of Friedelin Synthase from Maytenus ilicifolia Are Important for Substrate Binding Specificity. Molecules 2021, 26, 6806. https://doi.org/10.3390/molecules26226806
Mazzeu BF, Souza-Moreira TM, Oliveira AA, Remlinger M, Felippe LG, Valentini SR, Guido RVC, Zanelli CF, Furlan M. The Methionine 549 and Leucine 552 Residues of Friedelin Synthase from Maytenus ilicifolia Are Important for Substrate Binding Specificity. Molecules. 2021; 26(22):6806. https://doi.org/10.3390/molecules26226806
Chicago/Turabian StyleMazzeu, Bruna F., Tatiana M. Souza-Moreira, Andrew A. Oliveira, Melissa Remlinger, Lidiane G. Felippe, Sandro R. Valentini, Rafael V. C. Guido, Cleslei F. Zanelli, and Maysa Furlan. 2021. "The Methionine 549 and Leucine 552 Residues of Friedelin Synthase from Maytenus ilicifolia Are Important for Substrate Binding Specificity" Molecules 26, no. 22: 6806. https://doi.org/10.3390/molecules26226806
APA StyleMazzeu, B. F., Souza-Moreira, T. M., Oliveira, A. A., Remlinger, M., Felippe, L. G., Valentini, S. R., Guido, R. V. C., Zanelli, C. F., & Furlan, M. (2021). The Methionine 549 and Leucine 552 Residues of Friedelin Synthase from Maytenus ilicifolia Are Important for Substrate Binding Specificity. Molecules, 26(22), 6806. https://doi.org/10.3390/molecules26226806