
Ginsenoside Compound K Assisted G-Quadruplex Folding and Regulated G-Quadruplex-Containing Transcription

Abstract
:1. Introduction
2. Results
2.1. G4 Characterization
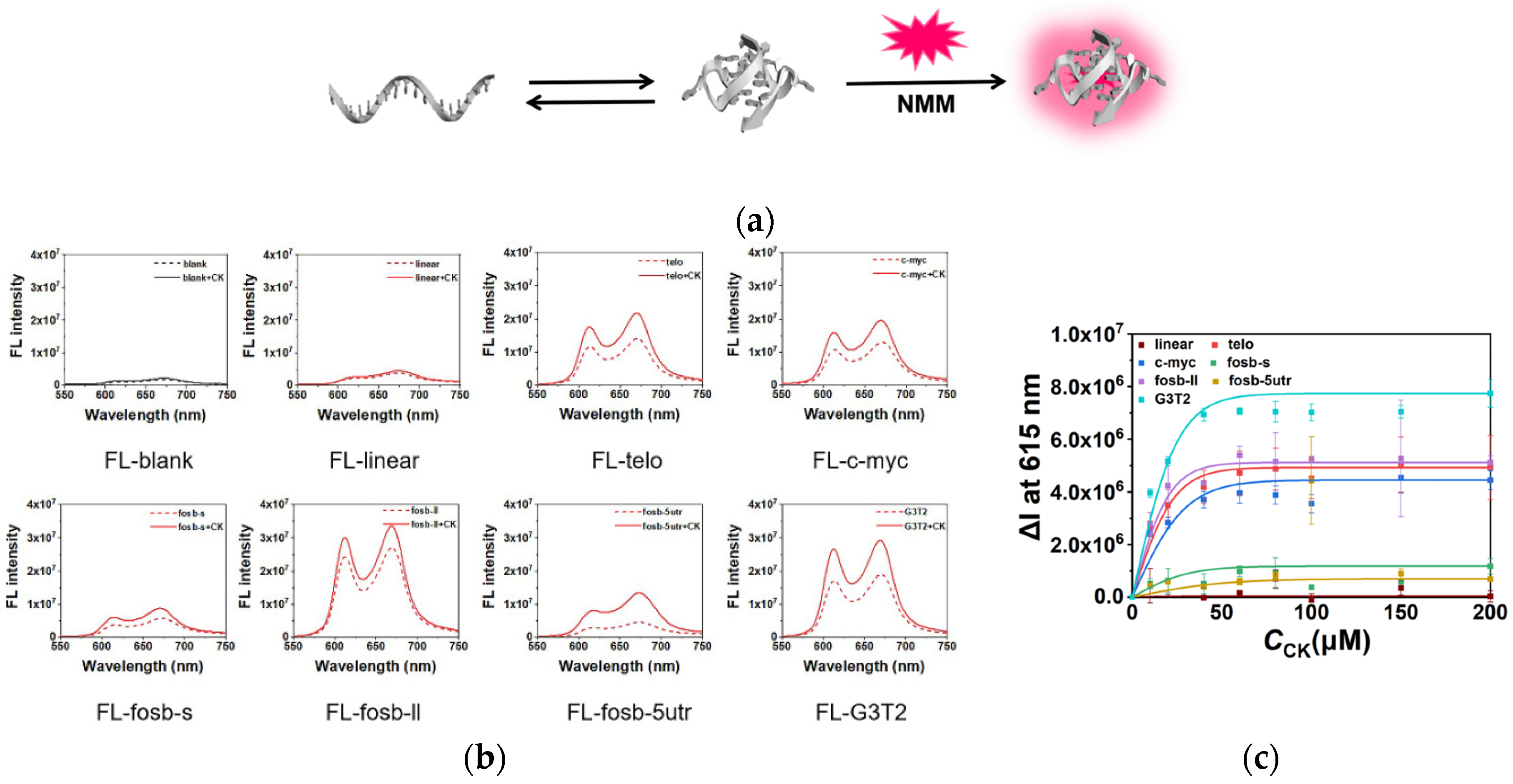
2.2. Stabilization of G4s by CK
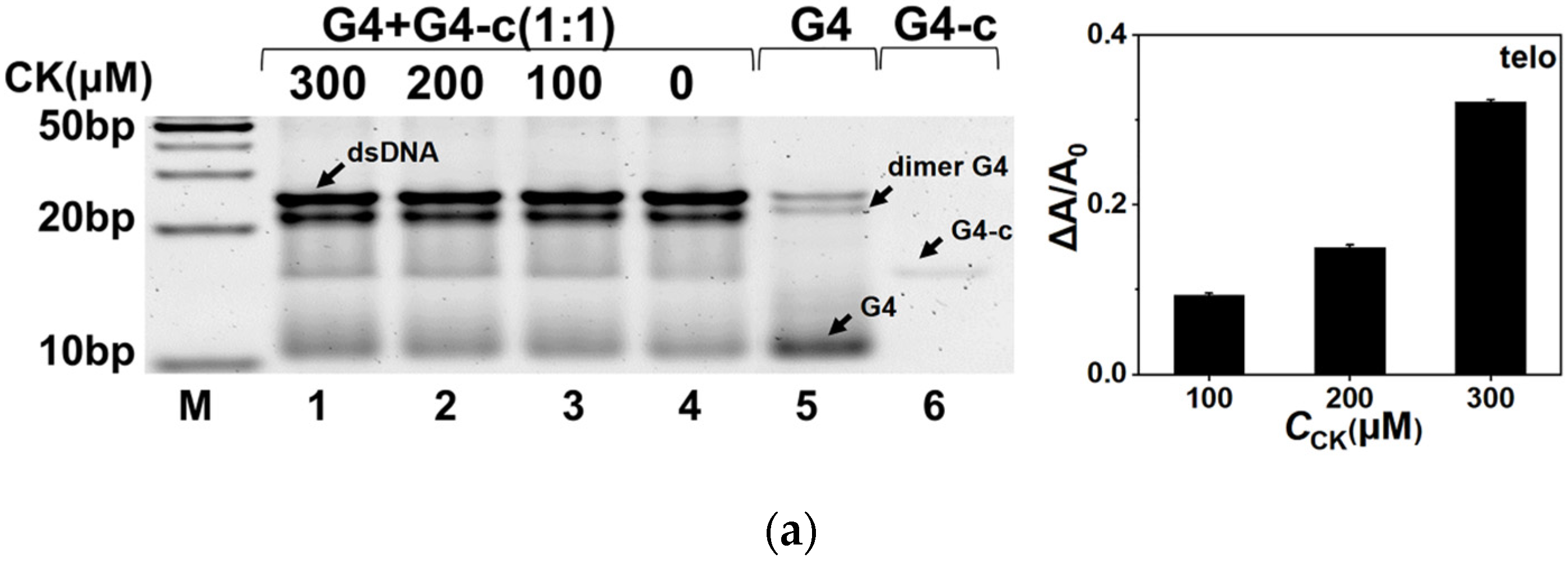
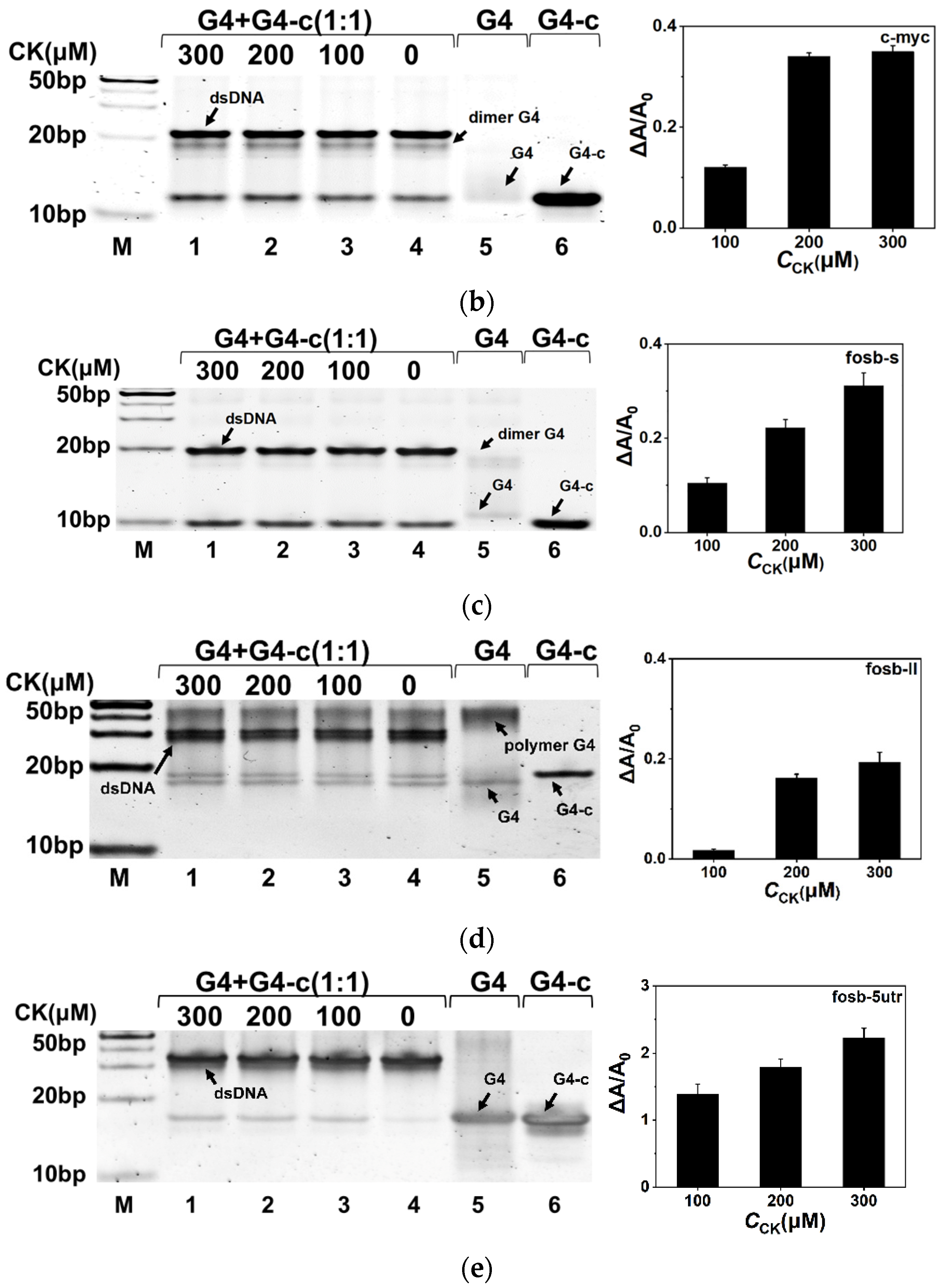
2.3. CK Facilitated G4 Folding in the Competition with Duplex
2.4. Quantitative Characterization of the Effect on G4 by CK in the Duplex System
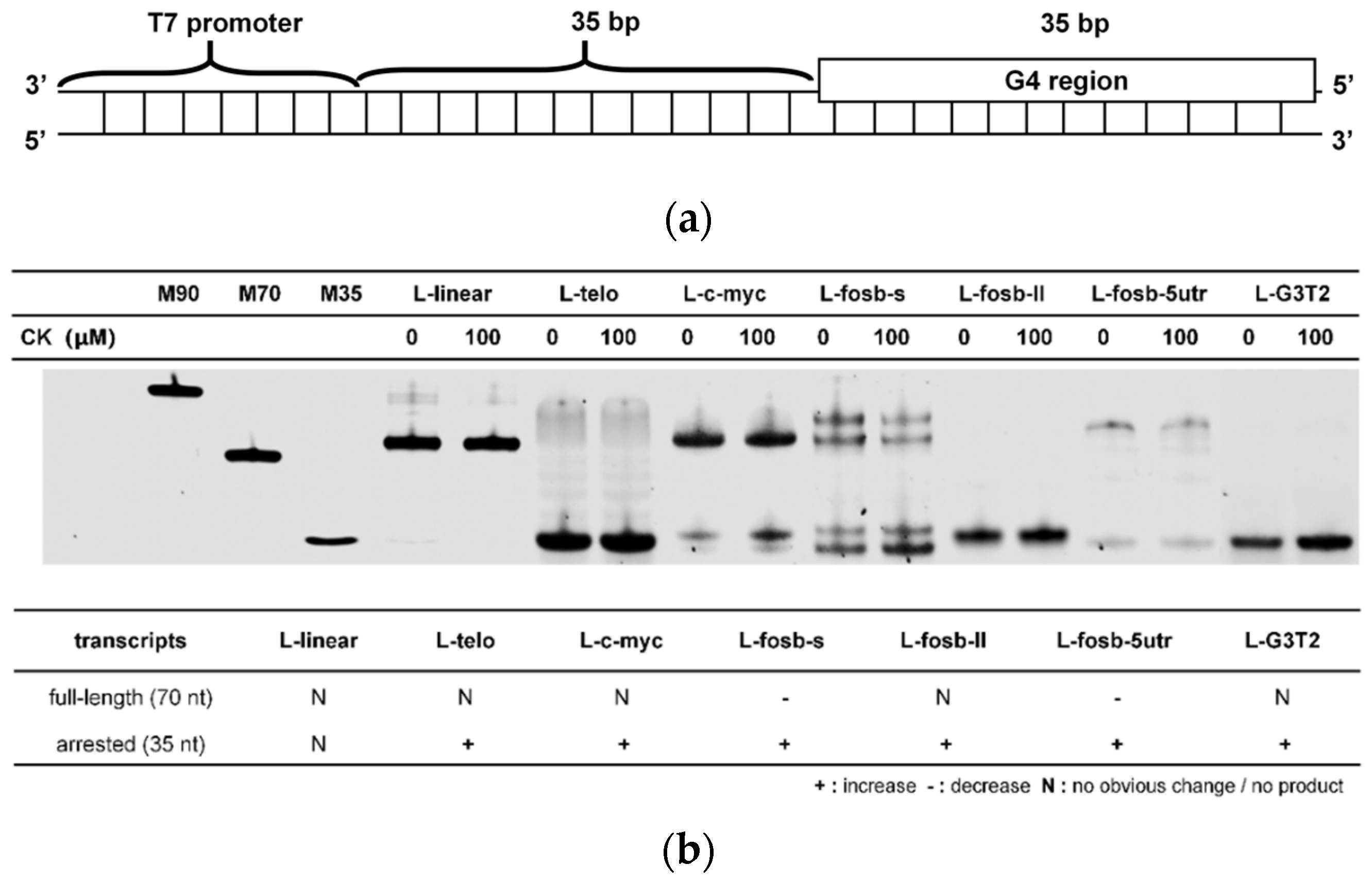
2.5. The Regulation of CK in G-Quadruplex-Containing Transcription
3. Discussion
4. Materials and Methods
4.1. Oligodeoxynucleotides and Materials
4.2. Circular Dichroism (CD) Measurement
4.3. Fluorescence Measurement
4.4. Native Polyacrylamide Gel Electrophoresis
4.5. Synthesis of DNA-AgNCs
4.6. Transcription Assays
4.7. Cell Counting Kit-8 (CCK-8)
4.8. Western Blot Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Yang, X.D.; Yang, Y.Y.; Ouyang, D.S.; Yang, G.P. A review of biotransformation and pharmacology of ginsenoside compound K. Fitoterapia 2015, 100, 208–220. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, M.; Chen, J.; Zhao, J.; Su, J.; Zhang, X. Ginsenoside Compound K Regulates HIF-1alpha-Mediated Glycolysis Through Bclaf1 to Inhibit the Proliferation of Human Liver Cancer Cells. Front. Pharmacol. 2020, 11, 2050. [Google Scholar] [CrossRef]
- Tian, F.; Wang, X.; Ni, H.; Feng, X.; Yuan, X.; Huang, Q. The ginsenoside metabolite compound K stimulates glucagon-like peptide-1 secretion in NCI-H716 cells by regulating the RhoA/ROCKs/YAP signaling pathway and cytoskeleton formation. J. Pharmacol. Sci. 2021, 145, 88–96. [Google Scholar] [CrossRef]
- Kuang, Y.Y.; Gao, X.; Niu, Y.J.; Shi, X.L.; Zhou, W. Polymorphic Characterization, Pharmacokinetics, and Anti-Inflammatory Activity of Ginsenoside Compound K Polymorphs. Molecules 2021, 26, 1983. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, P.; Shin, C.Y. A comprehensive review of the therapeutic and pharmacological effects of ginseng and ginsenosides in central nervous system. J. Ginseng. Res. 2013, 37, 8–29. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.-W.; Wang, C.-Z.; Yuan, C.-S. Ginsenosides from American ginseng: Chemical and pharmacological diversity. Phytochemistry 2011, 72, 689–699. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Lee, H.J. Ginsenoside Compound K: Insights into Recent Studies on Pharmacokinetics and Health-Promoting Activities. Biomolecules 2020, 10, 1028. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Jiang, Y.; Liu, T.; Luo, Y.; Diao, E.; Cao, Y.; Chen, L.; Zhang, L.; Gu, Q.; et al. Enhanced cytotoxic and apoptotic potential in hepatic carcinoma cells of chitosan nanoparticles loaded with ginsenoside compound K. Carbohydr. Polym. 2018, 198, 537–545. [Google Scholar] [CrossRef]
- Choi, E.; Kim, E.; Kim, J.H.; Yoon, K.; Kim, S.; Lee, J.; Cho, J.Y. AKT1-targeted proapoptotic activity of compound K in human breast cancer cells. J. Ginseng. Res. 2019, 43, 692–698. [Google Scholar] [CrossRef]
- Wu, Y.; Brosh, R.M., Jr. G-quadruplex nucleic acids and human disease. FEBS J. 2010, 277, 3470–3488. [Google Scholar] [CrossRef]
- Sengupta, A.; Roy, S.S.; Chowdhury, S. Non-duplex G-Quadruplex DNA Structure: A Developing Story from Predicted Sequences to DNA Structure-Dependent Epigenetics and Beyond. Acc. Chem. Res. 2021, 54, 46–56. [Google Scholar] [CrossRef]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The regulation and functions of DNA and RNA G-quadruplexes. Nat. Rev. Mol. Cell. Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Bochman, M.L.; Paeschke, K.; Zakian, V.A. DNA secondary structures: Stability and function of G-quadruplex structures. Nat. Rev. Genet. 2012, 13, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Endoh, T.; Sugimoto, N. Conformational Dynamics of the RNA G-Quadruplex and its Effect on Translation Efficiency. Molecules 2019, 2, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Meng, Z.; Guo, Q.; Wu, X.; Teulade-Fichou, M.P.; Yeow, E.K.L.; Shao, F. Fluorogenic Pt complexes distinguish the quantity and folding behavior of RNA G-quadruplexes between live cancerous and healthy cells. Chem. Commun. (Camb) 2020, 56, 14459–14462. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug. Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.H.; Wu, T.Y.; Huang, Q.; Jin, G. New substituted quinoxalines inhibit triple-negative breast cancer by specifically downregulating the c-MYC transcription. Nucleic Acids Res. 2019, 47, 10529–10542. [Google Scholar] [CrossRef]
- He, H.Z.; Li, K.; Yu, K.K.; Lu, P.L.; Feng, M.L.; Chen, S.Y.; Yu, X.Q. Additive- and column-free synthesis of rigid bis-coumarins as fluorescent dyes for G-quadruplex sensing via disaggregation-induced emission. Chem. Commun. (Camb) 2020, 56, 6870–6873. [Google Scholar] [CrossRef]
- Tera, M.; Hirokawa, T.; Okabe, S.; Sugahara, K.; Seimiya, H.; Shimamoto, K. Design and synthesis of a berberine dimer: A fluorescent ligand with high affinity towards G-quadruplexes. Chem. Eur. J. 2015, 21, 14519–14528. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, D.; Nakamura, K.; Tateishi-Karimata, H.; Ohmichi, T.; Sugimoto, N. Hydration of Watson−Crick Base Pairs and Dehydration of Hoogsteen Base Pairs Inducing Structural Polymorphism under Molecular Crowding Conditions. J. Am. Chem. Soc. 2009, 131, 3522–3531. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.N.; Chaires, J.B.; Gray, R.D.; Trent, J.O. Stability and kinetics of G-quadruplex structures. Nucleic Acids Res. 2008, 36, 5482–5515. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Tateishi-Karimata, H.; Ohyama, T.; Sugimoto, N. Effect of Potassium Concentration on Triplex Stability under Molecular Crowding Conditions. Molecules 2020, 25, 387. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, J.; Kim, M.J.; Kim, Y.H.; Ko, S.R.; Park, H.W.; Kim, M.K. Enzymatic formation of compound-K from ginsenoside Rb1 by enzyme preparation from cultured mycelia of Armillaria mellea. J. Ginseng. Res. 2016, 40, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Fan, Y.; Wei, W.; Wang, P.P.; Liu, Q.F.; Wei, Y.J.; Zhang, L.; Zhao, G.P.; Yue, J.M.; Zhou, Z.H. Production of bioactive ginsenoside compound K in metabolically engineered yeast. Cell Res. 2014, 24, 770–773. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, H.; Zhang, Y.; Li, J.; Wang, C.; Zhou, L.; Jia, Y.; Li, X. Synthesis and Biological Evaluation of Ginsenoside Compound K Derivatives as a Novel Class of LXRalpha Activator. Molecules 2017, 22, 1232. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Miyoshi, D.; Sugimoto, N. Effects of molecular crowding on the structures, interactions, and functions of nucleic acids. Chem. Rev. 2014, 114, 2733–2758. [Google Scholar] [CrossRef] [PubMed]
- Ambrus, A.; Chen, D.; Dai, J.; Bialis, T.; Jones, R.A.; Yang, D. Human telomeric sequence forms a hybrid-type intramolecular G-quadruplex structure with mixed parallel/antiparallel strands in potassium solution. Nucleic Acids Res. 2006, 34, 2723–2735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, C.I.; Choi, S.; Hsiang, J.-C.; Antoku, Y.; Vosch, T.; Bongiorno, A.; Tzeng, Y.-L.; Dickson, R.M. Oligonucleotide-Stabilized Ag Nanocluster Fluorophores. J. Am. Chem. Soc. 2008, 130, 5038–5039. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Tateishi-Karimata, H.; Tsuruoka, T.; Sugimoto, N. A Turn-On Detection of DNA Sequences by Means of Fluorescence of DNA-Templated Silver Nanoclusters via Unique Interactions of a Hydrated Ionic Liquid. Molecules 2018, 23, 2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.; Muller, S.; Yeoman, J.; Trentesaux, C.; Riou, J.; Balasubramanian, S. A Novel Small Molecule That Alters Shelterin Integrity and Triggers a DNA-Damage Response at Telomeres. J. Am. Chem. Soc. 2008, 130, 15758–15759. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Tateishi-Karimata, H.; Sugimoto, N. C-Rich Sequence in a Non-Template DNA Strand Regulates Structure Change of G-Quadruplex in a Template Strand during Transcription. Bull. Chem. Soc. Jpn. 2019, 92, 572–577. [Google Scholar] [CrossRef]
- Kim, D.Y.; Park, M.W.; Yuan, H.D.; Lee, H.J.; Kim, S.H.; Chung, S.H. Compound K Induces Apoptosis via CAMK-IV/AMPK Pathways in HT-29 Colon Cancer Cells. J. Agr. Food Chem. 2009, 57, 10573–10578. [Google Scholar] [CrossRef] [PubMed]
- Han, H.G.; Lee, H.J.; Sim, D.Y.; Im, E.; Park, J.E.; Park, W.Y.; Kim, S.Y.; Khil, J.H.; Shim, B.S.; Kim, S.H. Suppression of phosphoinositide 3-kinase/phosphoinositide-dependent kinase-1/serum and glucocorticoid-induced protein kinase pathway. Phytother. Res. 2021, 35, 4547–4554. [Google Scholar] [CrossRef]
- Yan, C.Y.; Shi, W.G.; Gu, J.W.; Lee, R.J.; Zhang, Y. Design of a Novel Nucleus-Targeted NLS-KALA-SA Nanocarrier to Delivery Poorly Water-Soluble Anti-Tumor Drug for Lung Cancer Treatment. J. Pharm. Sci. 2021, 110, 2432–2441. [Google Scholar] [CrossRef]
- Zhang, Z.; Du, G.-J.; Wang, C.-Z.; Wen, X.-D.; Calway, T.; Li, Z.; He, T.-C.; Du, W.; Bissonnette, M.; Musch, M.W.; et al. Compound K, a Ginsenoside Metabolite, Inhibits Colon Cancer Growth via Multiple Pathways Including p53-p21 Interactions. Int. J. Mol. Sci. 2013, 14, 2980–2995. [Google Scholar] [CrossRef]
- Tateishi-Karimata, H.; Isono, N.; Sugimoto, N. New insights into transcription fidelity: Thermal stability of non-canonical structures in template DNA regulates transcriptional arrest, pause, and slippage. PLoS ONE 2014, 9, e90580. [Google Scholar] [CrossRef] [Green Version]
- Tateishi-Karimata, H.; Ohyama, T.; Muraoka, T.; Tanaka, S.; Kinbara, K.; Sugimoto, N. New Modified Deoxythymine with Dibranched Tetraethylene Glycol Stabilizes G-Quadruplex Structures. Molecules 2020, 25, 705. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Structure |
|---|---|---|
| telo | GGGTTAGGGTTAGGGTTAGGGTTA | Hybrid |
| c-myc | GGCCGCGGGCGGGGTTCGGG | Parallel |
| fosb-s | GGCGCGGGCGGGGCGCGGG | Hybrid |
| fosb-ll | GGGGCGGGTGACGTAAGCAGGGGGGCGGG | Parallel |
| fosb-5utr | GAGGTACAGCGGCATCCTGTGGGGGCCTGGG | Hybrid |
| G3T2 | GGGTTGGGTGTGGGTTGGG | Hybrid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Qiu, Z.; Zhu, M.; Teng, Y. Ginsenoside Compound K Assisted G-Quadruplex Folding and Regulated G-Quadruplex-Containing Transcription. Molecules 2021, 26, 7339. https://doi.org/10.3390/molecules26237339
Zhang Y, Qiu Z, Zhu M, Teng Y. Ginsenoside Compound K Assisted G-Quadruplex Folding and Regulated G-Quadruplex-Containing Transcription. Molecules. 2021; 26(23):7339. https://doi.org/10.3390/molecules26237339
Chicago/Turabian StyleZhang, Yan, Zhidong Qiu, Ming Zhu, and Ye Teng. 2021. "Ginsenoside Compound K Assisted G-Quadruplex Folding and Regulated G-Quadruplex-Containing Transcription" Molecules 26, no. 23: 7339. https://doi.org/10.3390/molecules26237339
APA StyleZhang, Y., Qiu, Z., Zhu, M., & Teng, Y. (2021). Ginsenoside Compound K Assisted G-Quadruplex Folding and Regulated G-Quadruplex-Containing Transcription. Molecules, 26(23), 7339. https://doi.org/10.3390/molecules26237339