
Crossing of the Cystic Barriers of Toxoplasma gondii by the Fluorescent Coumarin Tetra-Cyclopeptide

 , , and
, , and
Abstract
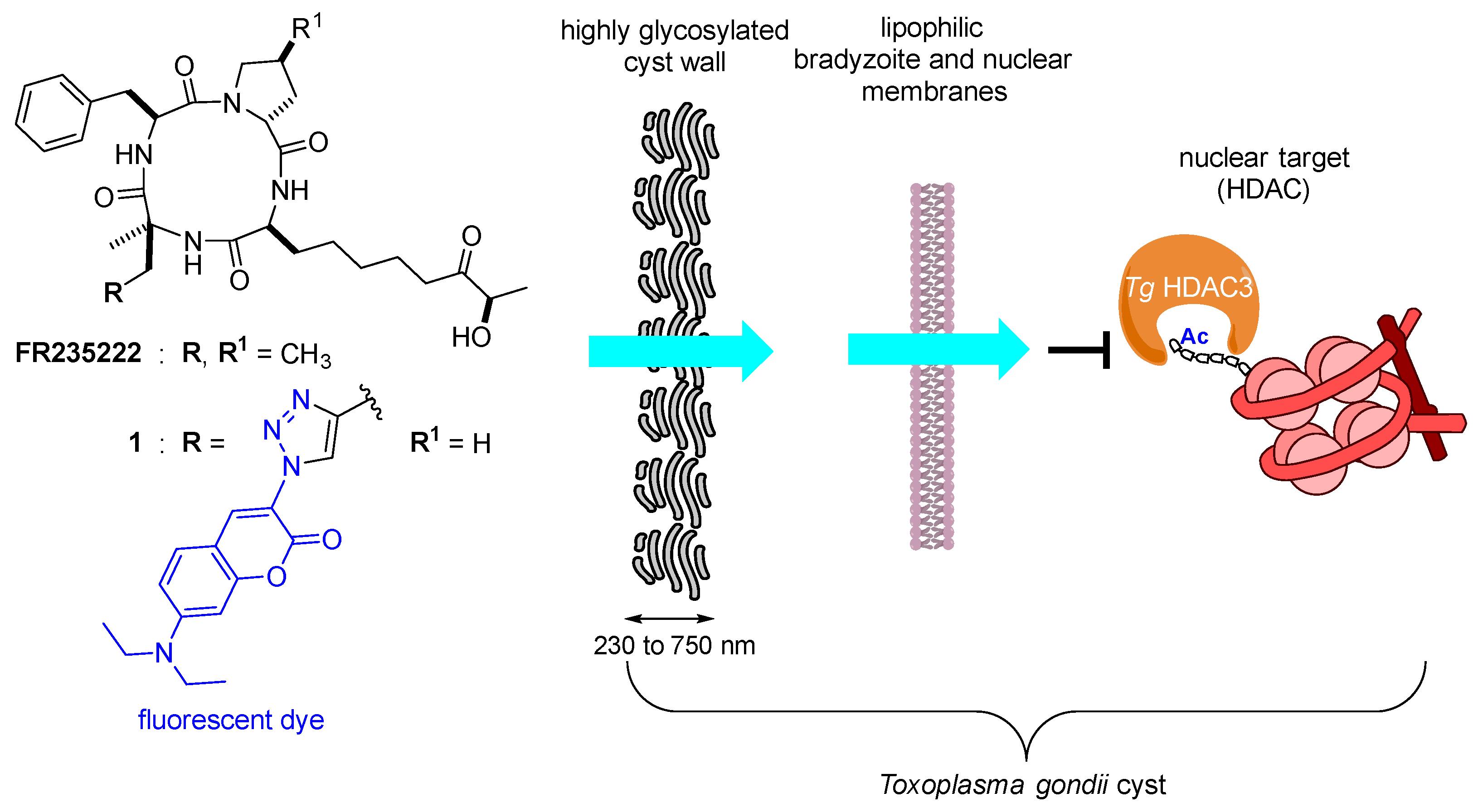
:1. Introduction
2. Results and Discussion
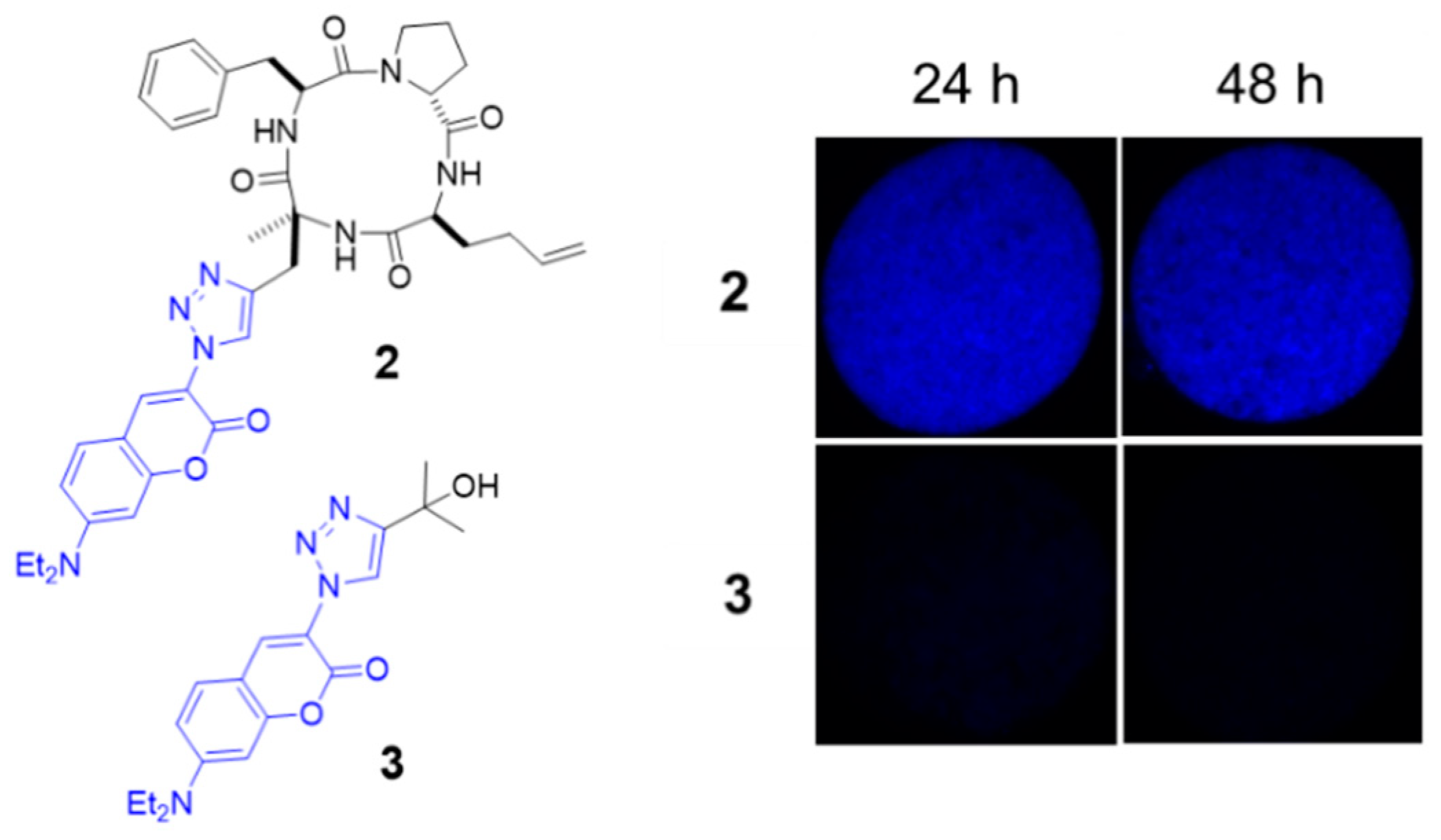
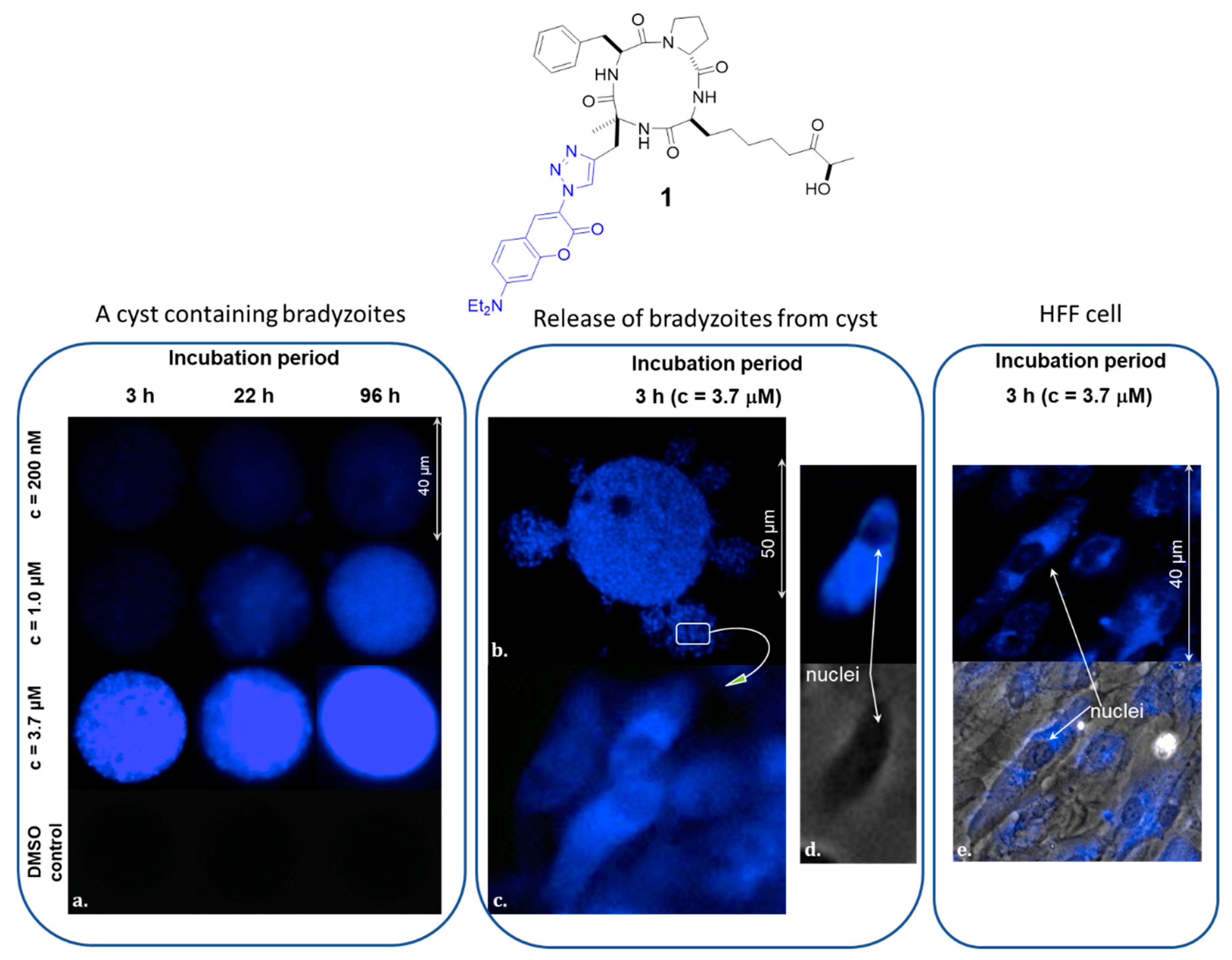
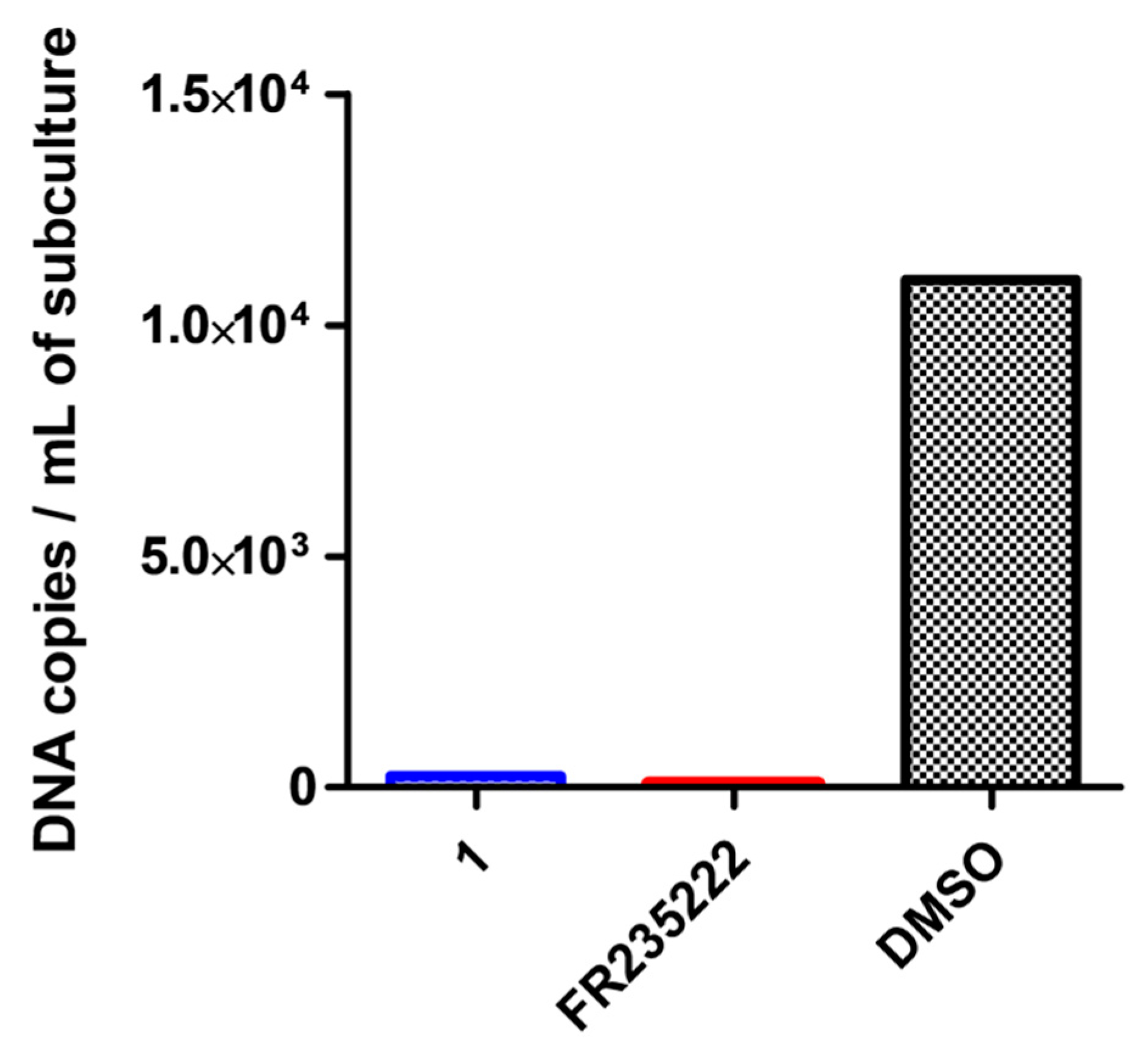
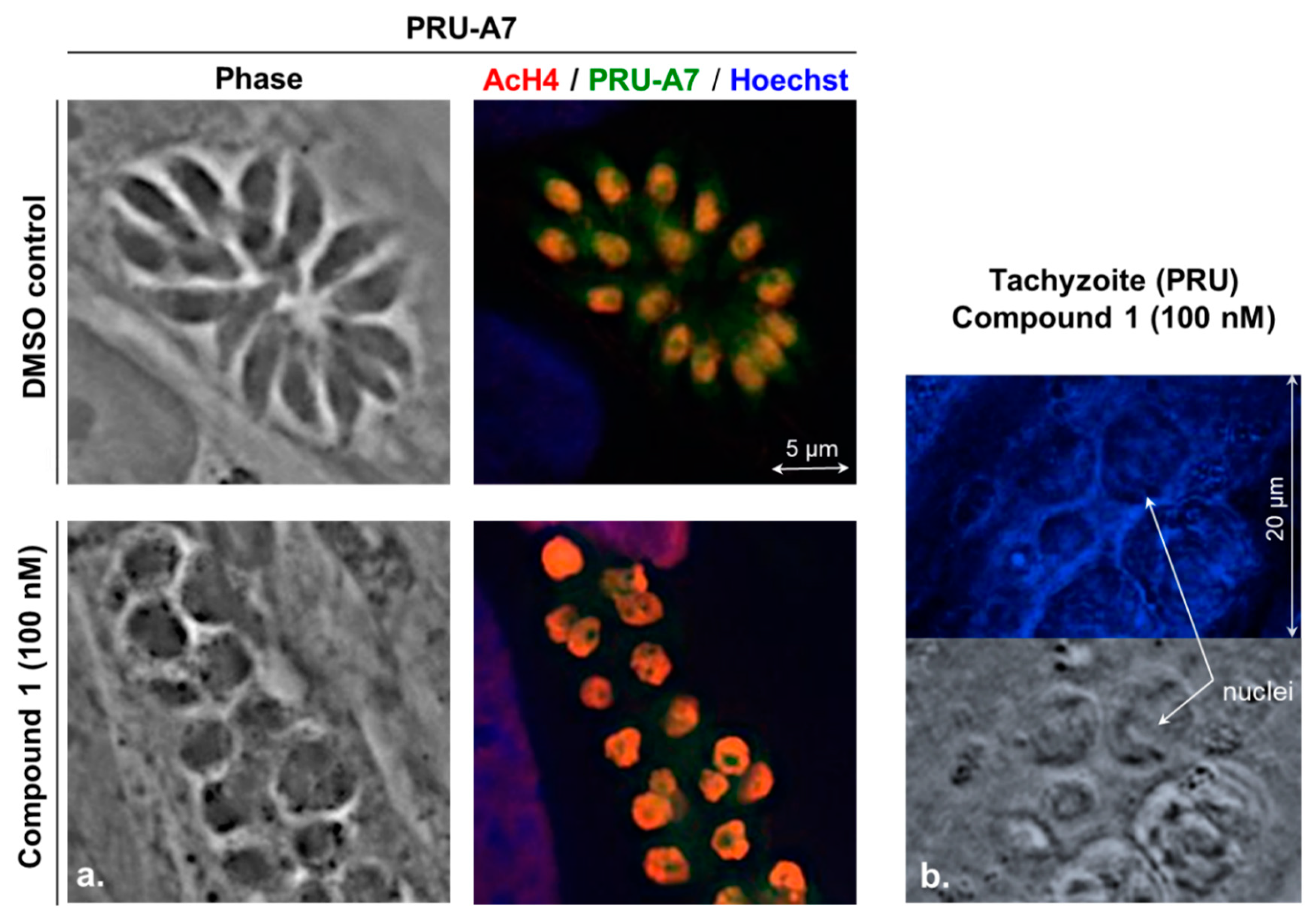
2.1. Evaluation of the Cyclic Tetrapeptide Penetration through the Cyst Wall and Bradyzoite Membranes
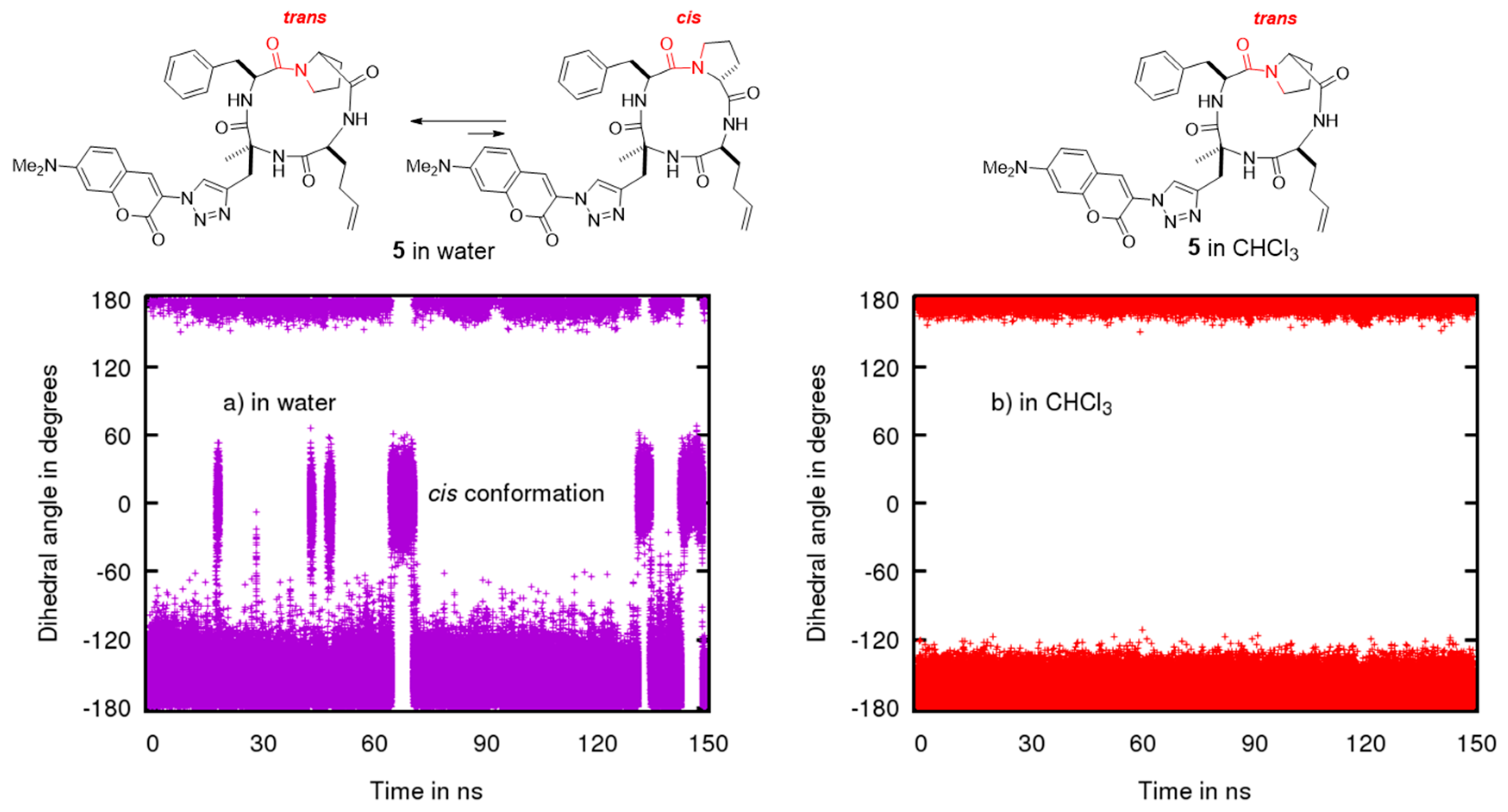
2.2. Conformational Studies by NMR
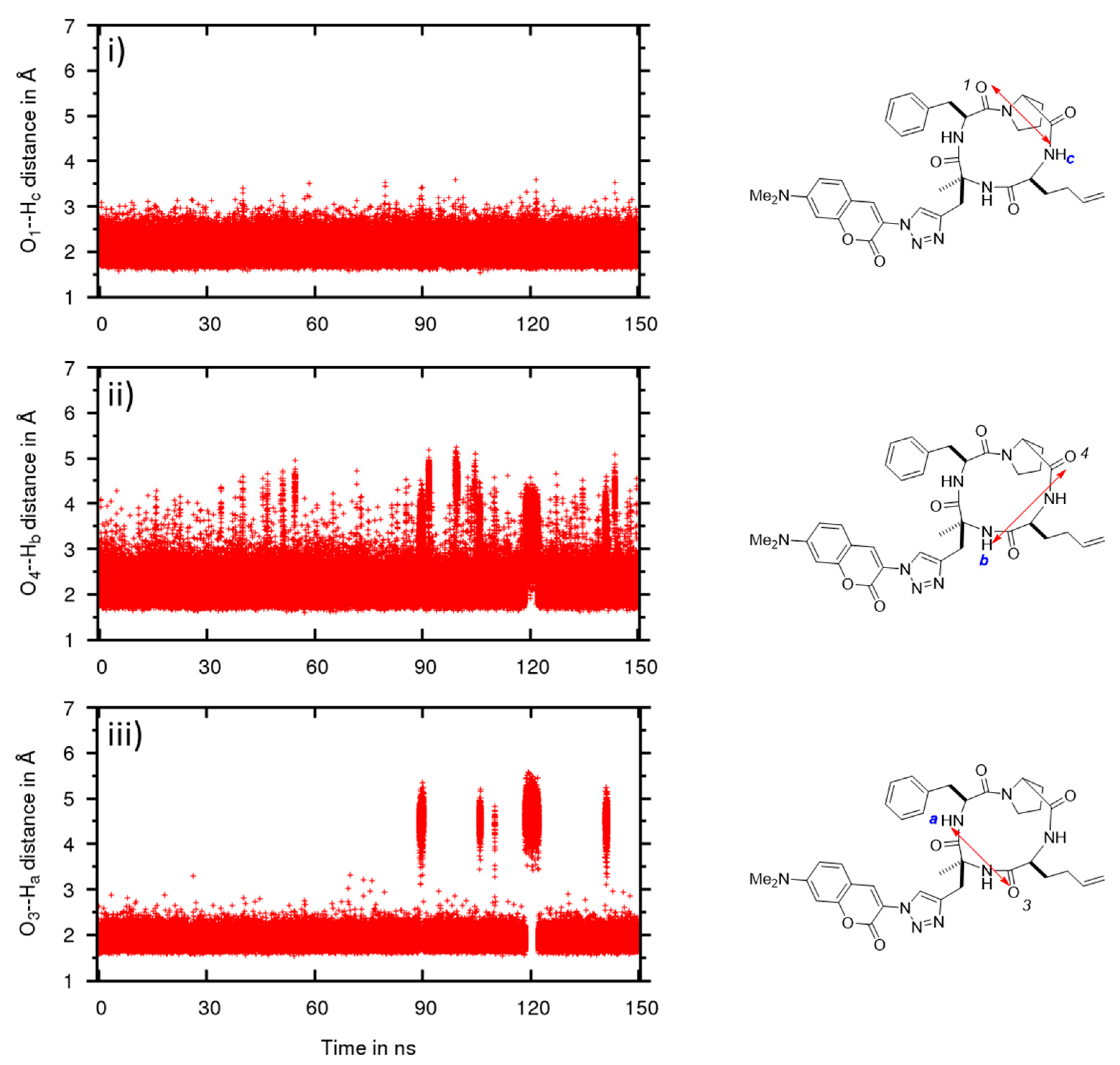
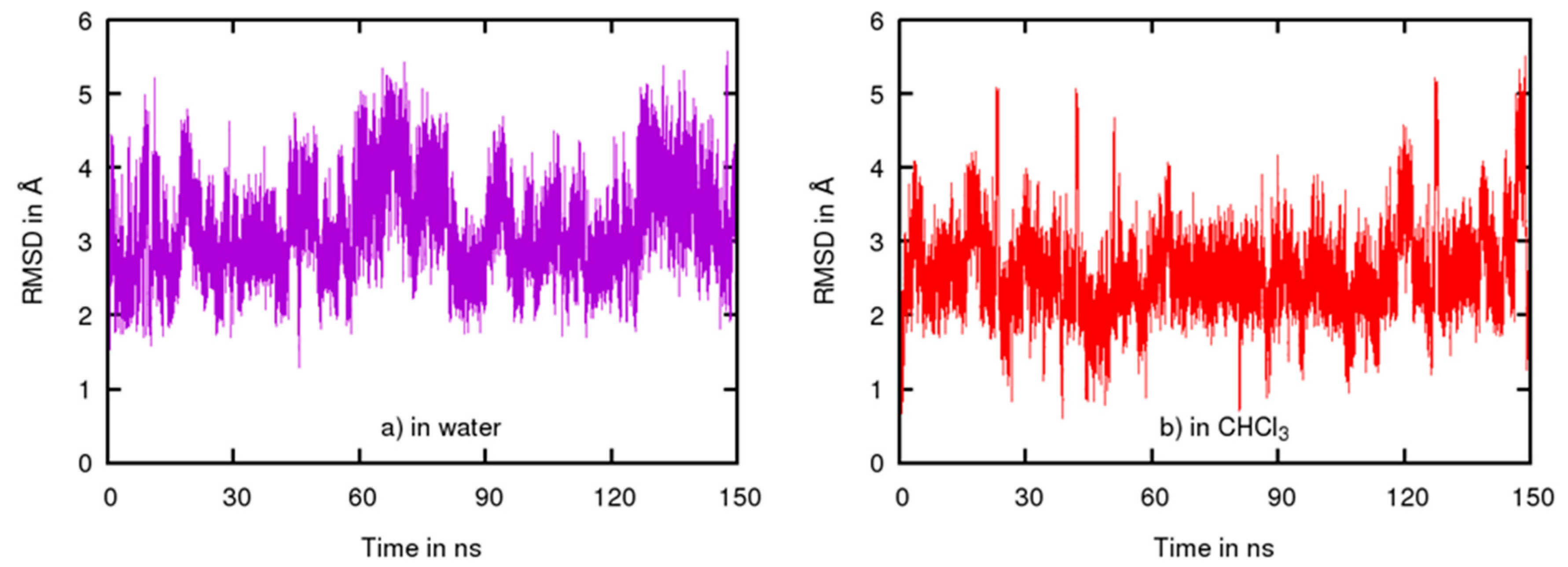
2.3. Conformational Studies by Molecular Modeling
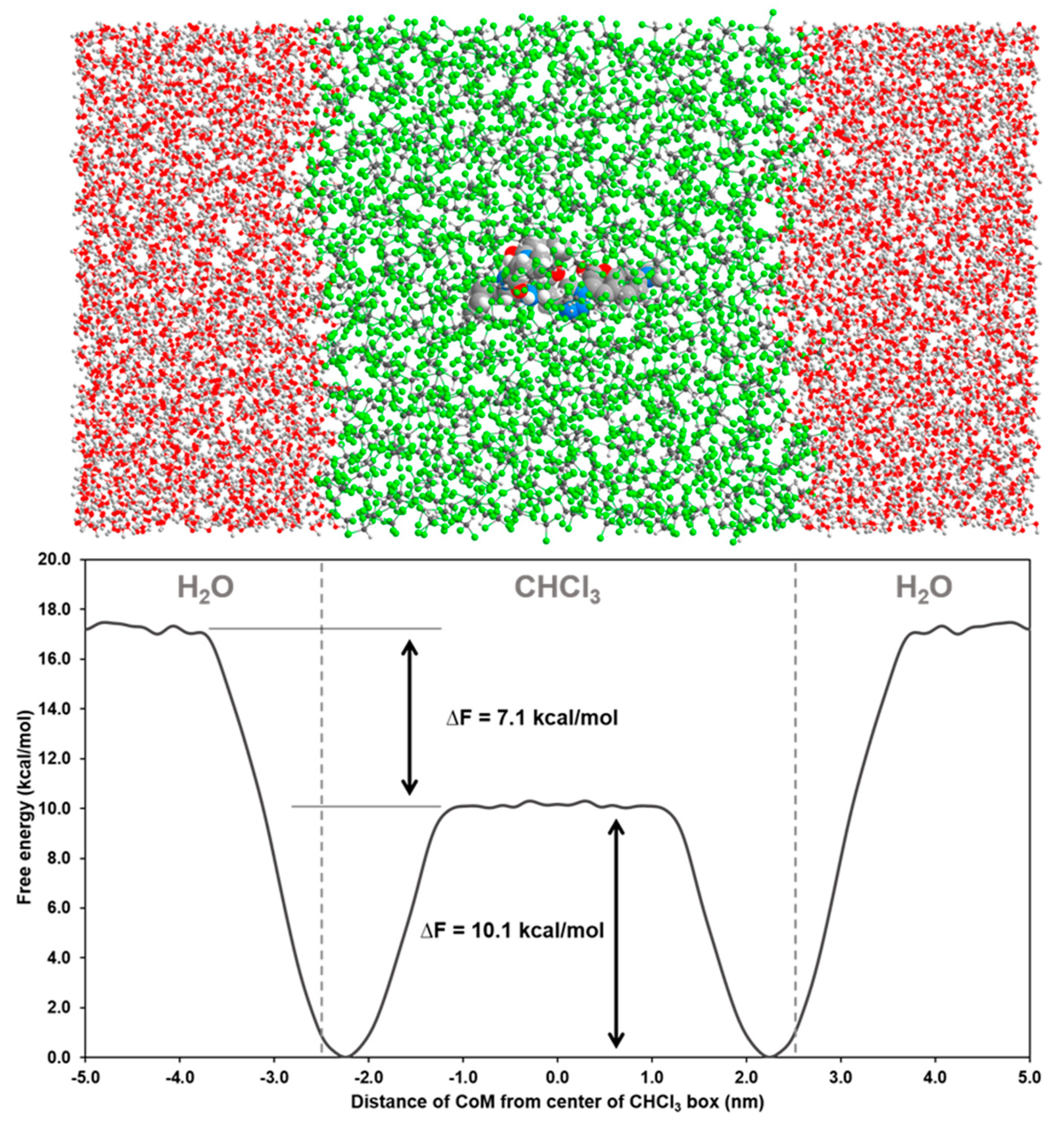
2.4. Permeability between Low and High Dielectric Environment
3. Conclusions
4. Materials and Methods
4.1. Biological Assay
4.1.1. Parasites Strains
4.1.2. In Vitro Parasites and Cell Culture
4.1.3. Mice
4.1.4. Mice Infection
4.1.5. Cysts Purification
4.1.6. Immunofluorescence Microscopy of Tachyzoites and Cysts
4.1.7. Drugs
4.2. Computation
4.2.1. Molecular Dynamics
4.2.2. Analysis Trajectories
4.2.3. Metadynamics in CHCl3/Water
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Lyons, R.E.; McLeod, R.; Craig, W.R. Toxoplasma Gondii Tachyzoite–Bradyzoite Interconversion. Trends Parasitol. 2002, 18, 198–201. [Google Scholar] [CrossRef]
- Lemgruber, L.; Lupetti, P.; Martins-Duarte, E.S.; De Souza, W.; Vommaro, R.C. The Organization of the Wall Filaments and Characterization of the Matrix Structures of Toxoplasma Gondii Cyst Form. Cell. Microbiol. 2011, 13, 1920–1932. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Blanchard, N.; Besteiro, S. The Bradyzoite: A Key Developmental Stage for the Persistence and Pathogenesis of Toxoplasmosis. Pathogens 2020, 9, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dard, C.; Marty, P.; Brenier-Pinchart, M.P.; Garnaud, C.; Fricker-Hidalgo, H.; Pelloux, H.; Pomares, C. Management of toxoplasmosis in transplant recipients: An update. Expert Rev. Anti Infect Ther. 2018, 16, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Mori, H.; Urano, Y.; Kinoshita, T.; Yoshimura, S.; Takase, S.; Hino, M. FR235222, a Fungal Metabolite, is a Novel Immunosuppressant That Inhibits Mammalian Histone Deacetylase III. Structure Determination. J. Antibiot. 2003, 56, 181–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bougdour, A.; Maubon, D.; Baldacci, P.; Ortet, P.; Bastien, O.; Bouillon, A.; Barale, J.-C.; Pelloux, H.; Ménard, R.; Hakimi, M.-A. Drug inhibition of HDAC3 and epigenetic control of differentiation in Apicomplexa parasites. J. Exp. Med. 2009, 206, 953–966. [Google Scholar] [CrossRef]
- Saksouk, N.; Bhatti, M.M.; Kieffer, S.; Smith, A.T.; Musset, K.; Garin, J.; Sullivan, W.J., Jr.; Cesbron-Delauw, M.F.; Hakimi, M.A. Histone-modifying complexes regulate gene expression pertinent to the differentiation of the protozoan parasite Toxoplasma gondii. Mol. Cell Biol. 2005, 23, 10301–10314. [Google Scholar] [CrossRef] [Green Version]
- Farhat, D.C.; Swale, C.; Dard, C.; Cannella, D.; Ortet, P.; Barakat, M.; Sindikubwabo, F.; Belmudes, L.; De Bock, P.J.; Couté, Y.; et al. A MORC-driven transcriptional switch controls Toxoplasma developmental trajectories and sexual commitment. Nat. Microbiol. 2020, 4, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Maubon, D.; Bougdour, A.; Wong, Y.-S.; Brenier-Pinchart, M.-P.; Curt, A.; Hakimi, M.-A.; Pelloux, H. Activity of the Histone Deacetylase Inhibitor FR235222 on Toxoplasma gondii: Inhibition of Stage Conversion of the Parasite Cyst Form and Study of New Derivative Compounds. Antimicrob. Agents Chemother. 2010, 54, 4843–4850. [Google Scholar] [CrossRef] [Green Version]
- Traoré, M.D.M.; Zwick, V.; Simões-Pires, C.A.; Nurisso, A.; Issa, M.; Cuendet, M.; Maynadier, M.; Wein, S.; Vial, H.; Jamet, H.; et al. Hydroxyl Ketone-Based Histone Deacetylase Inhibitors To Gain Insight into Class I HDAC Selectivity versus That of HDAC6. ACS Omega 2017, 2, 1550–1562. [Google Scholar] [CrossRef] [PubMed]
- Francisco Hilário, F.; Traoré, M.D.M.; Zwick, V.; Berry, L.; Simões-Pires, C.A.; Cuendet, M.; Fantozzi, N.; Pereira de Freitas, R.; Maynadier, M.; Wein, S.; et al. Synthesis of an Uncharged Tetra-cyclopeptide Acting as a Transmembrane Carrier: Enhanced Cellular and Nuclear Uptake. Org. Lett. 2017, 19, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Sarojini, V.; Cameron, A.J.; Varnava, K.G.; Denny, W.A.; Sanjayan, G. Cyclic Tetrapeptides from Nature and Design: A Review of Synthetic Methodologies, Structure, and Function. Chem. Rev. 2019, 119, 10318–10359. [Google Scholar] [CrossRef] [PubMed]
- Ahlbach, C.L.; Lexa, K.W.; Bockus, A.T.; Chen, V.; Crews, P.; Jacobson, M.P.; Lokey, R.S. Beyond cyclosporine A: Conformation-dependent passive membrane permeabilities of cyclic peptide natural products. Future Med. Chem. 2015, 7, 2121–2130. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, P.G.; Sahni, A.; Pei, D. Understanding Cell Penetration of Cyclic Peptides. Chem. Rev. 2019, 119, 10241–10287. [Google Scholar] [CrossRef] [PubMed]
- Buckton, L.K.; Rahimi, M.N.; McAlpine, S.R. Cyclic Peptides as Drugs for Intracellular Targets: The Next Frontier in Peptide Therapeutic Development. Chem. Eur. J. 2021, 27, 1487–1513. [Google Scholar] [CrossRef]
- Di Micco, S.; Terracciano, S.; Bruno, I.; Rodriquez, M.; Riccio, R.; Taddei, M.; Bifulco, G. Molecular Modeling Studies toward the Structural Optimization of New Cyclopeptide-Based HDAC Inhibitors Modeled on the Natural Product FR235222. Bioorg. Med. Chem. 2008, 16, 8635–8642. [Google Scholar] [CrossRef]
- Matsson, P.; Doak, B.C.; Over, B.; Kihlberg, J. Cell permeability beyond the rule of 5. Adv. Drug Deliv. Rev. 2016, 101, 42–61. [Google Scholar] [CrossRef]
- Whitty, A.; Zhong, M.; Viarengo, L.; Beglov, D.; Hall, D.R.; Vajda, S. Quantifying the Chameleonic Properties of Macrocycles and Other High-Molecular-Weight Drugs. Drug Discov. Today 2016, 21, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Rossi Sebastiano, M.; Doak, B.C.; Backlund, M.; Poongavanam, V.; Over, B.; Ermondi, G.; Caron, G.; Matsson, P.; Kihlberg, J. Impact of Dynamically Exposed Polarity on Permeability and Solubility of Chameleonic Drugs Beyond the Rule of 5. J. Med. Chem. 2018, 61, 4189–4202. [Google Scholar] [CrossRef]
- Furukawa, A.; Schwochert, J.; Pye, C.R.; Asano, D.; Edmondson, Q.D.; Turmon, A.C.; Klein, V.G.; Ono, S.; Okada, O.; Lokey, R.S. Drug-Like Properties in Macrocycles above MW 1000: Backbone Rigidity versus Side-Chain Lipophilicity. Angew. Chem. Int. Ed. 2020, 48, 21571–21577. [Google Scholar] [CrossRef]
- Krishnamoorthy, S.; Xie, F.; Cash, B.M.; Long, S.; Barnhill, H.N.; Wang, Q. A Fluorogenic 1,3-Dipolar Cycloaddition Reaction of 3-Azidocoumarins and Acetylenes. Org. Lett. 2004, 6, 4603–4606. [Google Scholar]
- Brenier-Pinchart, M.P.; Morand-Bui, V.; Fricker-Hidalgo, H.; Equy, V.; Marlu, R.; Pelloux, H. Adapting a Conventional Pcr Assay for Toxoplasma gondii Detection to Real-Time Quantitative Pcr Including a Competitive Internal Control. Parasite 2007, 14, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Rezai, T.; Yu, B.; Millhauser, G.L.; Jacobson, M.P.; Lokey, R.S. Testing the Conformational Hypothesis of Passive Membrane Permeability Using Synthetic Cyclic Peptide Diastereomers. J. Am. Chem. Soc. 2006, 128, 2510–2511. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.N.; Hill, T.A.; Fairlie, D.P. Connecting Hydrophobic Surfaces in Cyclic Peptides Increases Membrane Permeability. Angew. Chem. Int. Ed. 2021, 60, 8385–8390. [Google Scholar] [CrossRef]
- Oakley, M.T.; Johnston, R.L. Exploring the Energy Landscapes of Cyclic Tetrapeptides with Discrete Path Sampling. J. Chem. Theory Comput. 2013, 9, 650–657. [Google Scholar] [CrossRef]
- Cierpicki, T.; Otlewski, J. Amide Proton Temperature Coefficients as Hydrogen Bond Indicators in Proteins. J. Biomol. NMR 2001, 21, 249–261. [Google Scholar] [CrossRef]
- Schwochert, J.; Lao, Y.; Pye, C.R.; Naylor, M.R.; Desai, P.V.; Gonzalez Valcarcel, I.C.; Barrett, J.A.; Sawada, G.; Blanco, M.-J.; Lokey, R.S. Stereochemistry Balances Cell Permeability and Solubility in the Naturally Derived Phepropeptin Cyclic Peptides. ACS Med. Chem. Lett. 2016, 7, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, D.A.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E.; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Ghoreishi, D.; Gilson, M.K.; et al. AMBER 2018; University of California: San Francisco, CA, USA, 2018. [Google Scholar]
- Leung, S.S.F.; Mijalkovic, J.; Borrelli, K.; Jacobson, M.P. Testing Physical Models of Passive Membrane Permeation. J. Chem. Inf. Model. 2012, 52, 1621–1636. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Xu, Z. Drug Permeability Prediction Using PMF Method. J. Mol. Model. 2013, 19, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Bennion, B.J.; Be, N.A.; McNerney, M.W.; Lao, V.; Carlson, E.M.; Valdez, C.A.; Malfatti, M.A.; Enright, H.A.; Nguyen, T.H.; Lightstone, F.C.; et al. Predicting a Drug’s Membrane Permeability: A Computational Model Validated With in Vitro Permeability Assay Data. J. Phys. Chem. B. 2017, 121, 5228–5237. [Google Scholar] [CrossRef] [Green Version]
- Neale, C.; Pomès, R. Sampling Errors in Free Energy Simulations of Small Molecules in Lipid Bilayers. Biochim. Biophys. Acta 2018, 1858, 2539–2548. [Google Scholar] [CrossRef] [PubMed]
- Rezai, T.; Bock, J.E.; Zhou, M.V.; Kalyanaraman, C.; Lokey, R.S.; Jacobson, M.P. Conformational Flexibility, Internal Hydrogen Bonding, and Passive Membrane Permeability: Successful in Silico Prediction of the Relative Permeabilities of Cyclic Peptides. J. Am. Chem. Soc. 2006, 128, 14073–14080. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Zhang, H.; Ibrahim, H.M.; Ui, F.; Ogiso, A.; Xuan, X. Construction of Toxoplasme gondii Bradyzoite Expressing the Green Fluorescent Protein. Parasitol. Int. 2008, 57, 219–222. [Google Scholar] [CrossRef]
- Huynh, M.-H.; Carruthers, V.B. Tagging of Endogenous Genes in a Toxoplasma gondii Strain Lacking Ku80. Eukaryot. Cell 2009, 8, 530–539. [Google Scholar] [CrossRef] [Green Version]
- Fox, B.A.; Falla, A.; Rommereim, L.M.; Tomita, T.; Gigley, J.P.; Mercier, C.; Cesbron-Delauw, M.-F.; Weiss, L.M.; Bzik, D.J. Type II Toxoplasma gondii KU80 Knockout Strains enable Functional Analysis of Genes Required for Cyst Development and Latent Infection. Eukaryot. Cell 2011, 10, 1193–1206. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, A.W.; Overdulse, J.P.; Hoenderboom, J.M. Separation of Isospora (Toxoplasma) gondii cysts and cystozoites from mouse brain tissue by continuous density-gradient centrifugation. Parasitology 1981, 83, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Pracht, P.; Bohle, F.; Grimme, S. Automated Exploration of the Low-Energy Chemical Space with Fast Quantum Chemical Methods. Phys. Chem. Chem. Phys. 2020, 22, 7169–7192. [Google Scholar] [CrossRef]
- Bayly, C.I.; Cieplak, P.; Cornell, W.; Kollman, P.A. A Well-Behaved Electrostatic Potential Based Method Using Charge Restraints for Deriving Atomic Charges: The RESP Model. J. Phys. Chem. 1993, 97, 10269–10280. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Cieplak, P.; Caldwell, J.; Kollman, P. Molecular Mechanical Models for Organic and Biological Systems Going beyond the Atom Centered Two Body Additive Approximation: Aqueous Solution Free Energies of Methanol and N-Methyl Acetamide, Nucleic Acid Base, and Amide Hydrogen Bonding and Chloroform/Water Partition Coefficients of the Nucleic Acid Bases. J. Comput. Chem. 2001, 22, 1048–1057. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of Simple Potential Functions for Simulating Liquid Water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Martínez, L.; Andrade, R.; Birgin, E.G.; Martínez, J.M. Packmol: A Package for Building Initial Configurations for Molecular Dynamics Simulations. J. Comput. Chem. 2009, 30, 2157–2164. [Google Scholar] [CrossRef]
- Ryckaert, J.-P.; Ciccotti, G.; Berendsen, H.J.C. Numerical Integration of the Cartesian Equations of Motion of a System with Constraints: Molecular Dynamics of n-Alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Ren, P.; Ponder, J.W. Consistent treatment of inter-and Intreamolecular polarization in Molecular Mechanics Calculations. J. Comput. Chem. 2002, 23, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Bonomi, M.; Branduardi, D.; Bussi, G.; Camilloni, C.; Provasi, D.; Raiteri, P.; Donadio, D.; Marinelli, F.; Pietrucci, F.; Broglia, R.A.; et al. PLUMED: A Portable Plugin fot Free-Energy Calculations with Moleculars Dynamics. Comput. Phys. Commun. 2009, 180, 1961–1972. [Google Scholar] [CrossRef] [Green Version]
- Tribello, G.A.; Bonomi, M.; Branduardi, D.; Camilloni, C.; Bussi, G. PLUMED 2: New Feathers for an Old Bird. Comput. Phys. Commun. 2014, 185, 604–613. [Google Scholar] [CrossRef] [Green Version]
- Bonomi, M.; Bussi, G.; Camilloni, C.; Tribello, G.A.; Banáš, P.; Barducci, A.; Bernetti, M.; Bolhuis, P.G.; Bottaro, S.; Branduardi, D.; et al. Promting Transparency and Reproductibility in Enhanced Molecular Simulations. Nat. Methods 2019, 16, 670–673. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cyclopeptides | T. gondii EC50 (nM) | HFF EC50 (nM) |
|---|---|---|
| FR235222 | 7.6 ± 0.6 | 107 ± 18 |
| 1 | 43 ± 2.2 | 280 ± 20 |
| FR235222 (90 nM) | Compound 1 (90 nM) | DMSO | |
|---|---|---|---|
| RH T99A | + | + | + |
| RH Wild Type | − | − | + |

| Title 1 | Water | 0 Hbond | 1 Hbond | 2 Hbonds | 3 Hbonds | Total |
|---|---|---|---|---|---|---|
| O2 | H/donor | 6% | 51% | 42% | 1% | 138% |
| O3 | H/donor | 6% | 54% | 39% | 1% | 135% |
| O1 | H/donor | 12% | 65% | 22% | 1% | 112% |
| O5 | H/donor | 15% | 60% | 24% | 1% | 111% |
| O4 | H/donor | 13% | 69% | 18% | 0% | 105% |
| N-Ha | O/acceptor | 55% | 45% | 0% | 0% | 45% |
| NA | H/donor | 60% | 37% | 3% | 0% | 43% |
| NB | H/donor | 72% | 27% | 1% | 0% | 29% |
| N-Hc | O/acceptor | 76% | 24% | 0% | 0% | 24% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dard, C.; Leforestier, B.; Francisco Hilário, F.; Traoré, M.D.M.; Lespinasse, M.-A.; Pérès, B.; Molina, M.-C.; Pereira de Freitas, R.; Milet, A.; Maubon, D.; et al. Crossing of the Cystic Barriers of Toxoplasma gondii by the Fluorescent Coumarin Tetra-Cyclopeptide. Molecules 2021, 26, 7506. https://doi.org/10.3390/molecules26247506
Dard C, Leforestier B, Francisco Hilário F, Traoré MDM, Lespinasse M-A, Pérès B, Molina M-C, Pereira de Freitas R, Milet A, Maubon D, et al. Crossing of the Cystic Barriers of Toxoplasma gondii by the Fluorescent Coumarin Tetra-Cyclopeptide. Molecules. 2021; 26(24):7506. https://doi.org/10.3390/molecules26247506
Chicago/Turabian StyleDard, Céline, Baptiste Leforestier, Flaviane Francisco Hilário, Mohamed Dit Mady Traoré, Marie-Ange Lespinasse, Basile Pérès, Marie-Carmen Molina, Rossimiriam Pereira de Freitas, Anne Milet, Danièle Maubon, and et al. 2021. "Crossing of the Cystic Barriers of Toxoplasma gondii by the Fluorescent Coumarin Tetra-Cyclopeptide" Molecules 26, no. 24: 7506. https://doi.org/10.3390/molecules26247506
APA StyleDard, C., Leforestier, B., Francisco Hilário, F., Traoré, M. D. M., Lespinasse, M. -A., Pérès, B., Molina, M. -C., Pereira de Freitas, R., Milet, A., Maubon, D., & Wong, Y. -S. (2021). Crossing of the Cystic Barriers of Toxoplasma gondii by the Fluorescent Coumarin Tetra-Cyclopeptide. Molecules, 26(24), 7506. https://doi.org/10.3390/molecules26247506