
Effect-Directed Profiling of 17 Different Fortified Plant Extracts by High-Performance Thin-Layer Chromatography Combined with Six Planar Assays and High-Resolution Mass Spectrometry

Abstract
:
1. Introduction
2. Results and Discussion
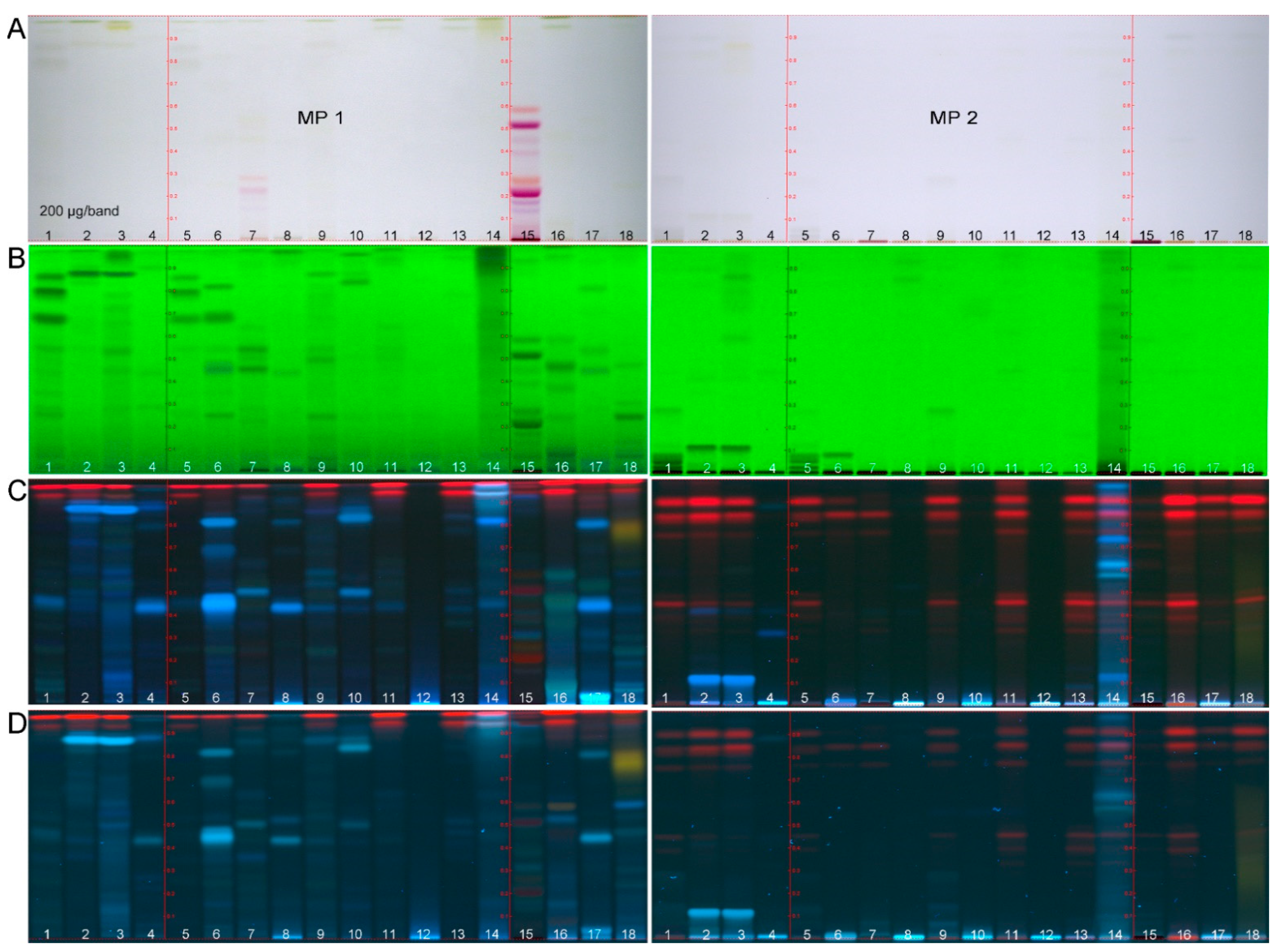
2.1. HPTLC-UV/Vis/FLD Method Development
2.2. Development of the Effect-Directed Profiling
2.2.1. Antibacterial Activity
2.2.2. Enzyme Inhibiting Activity
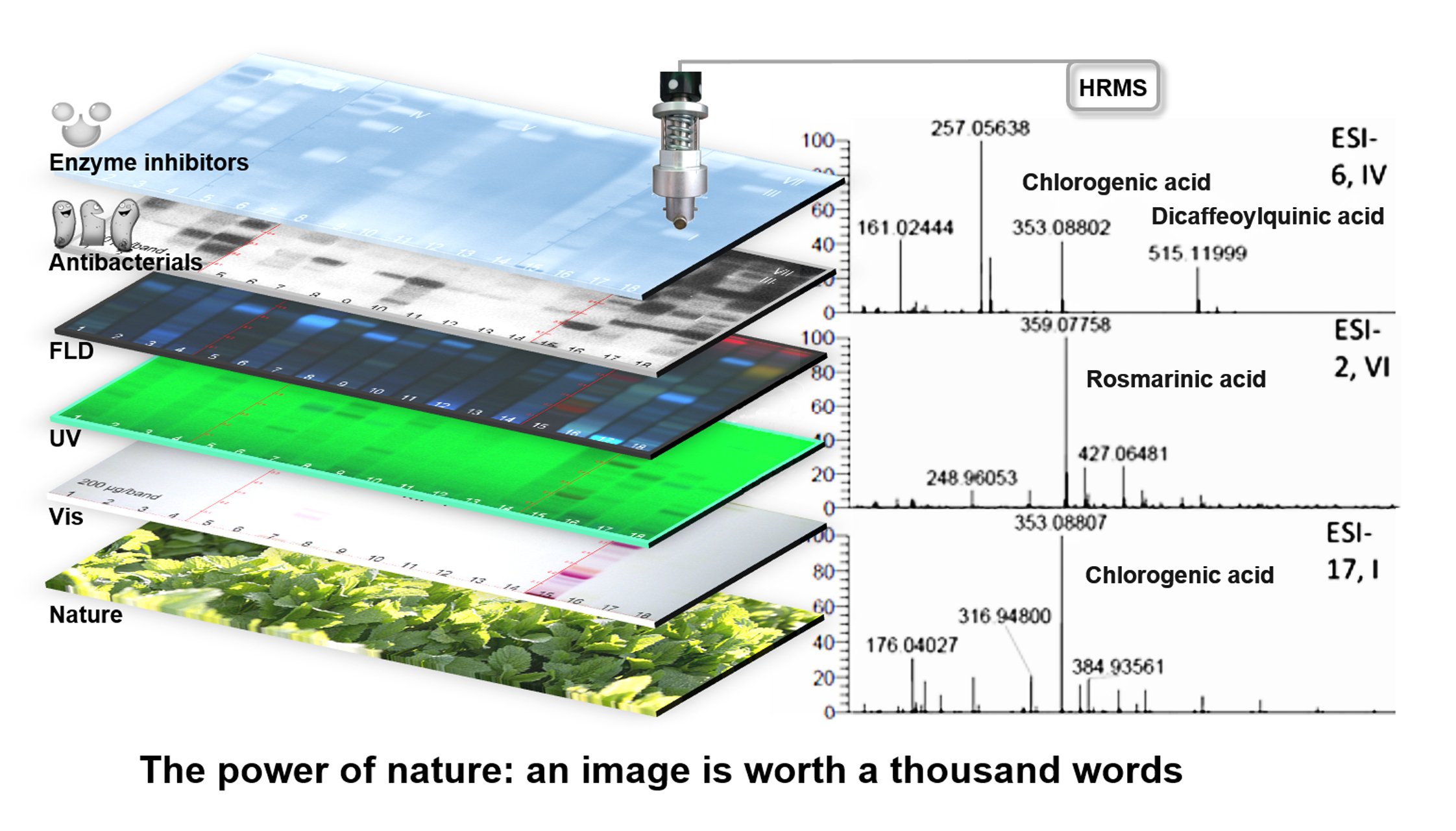
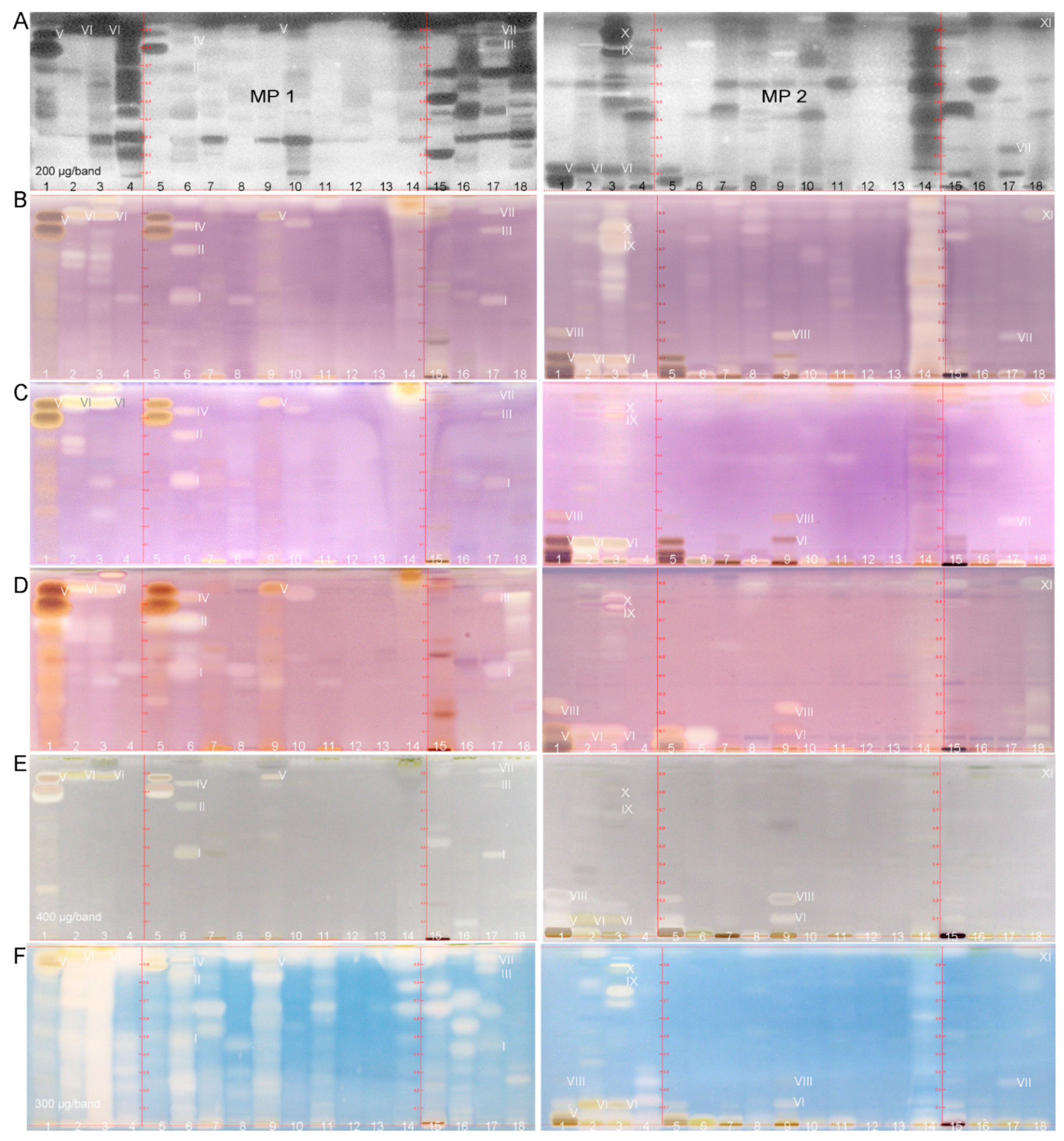
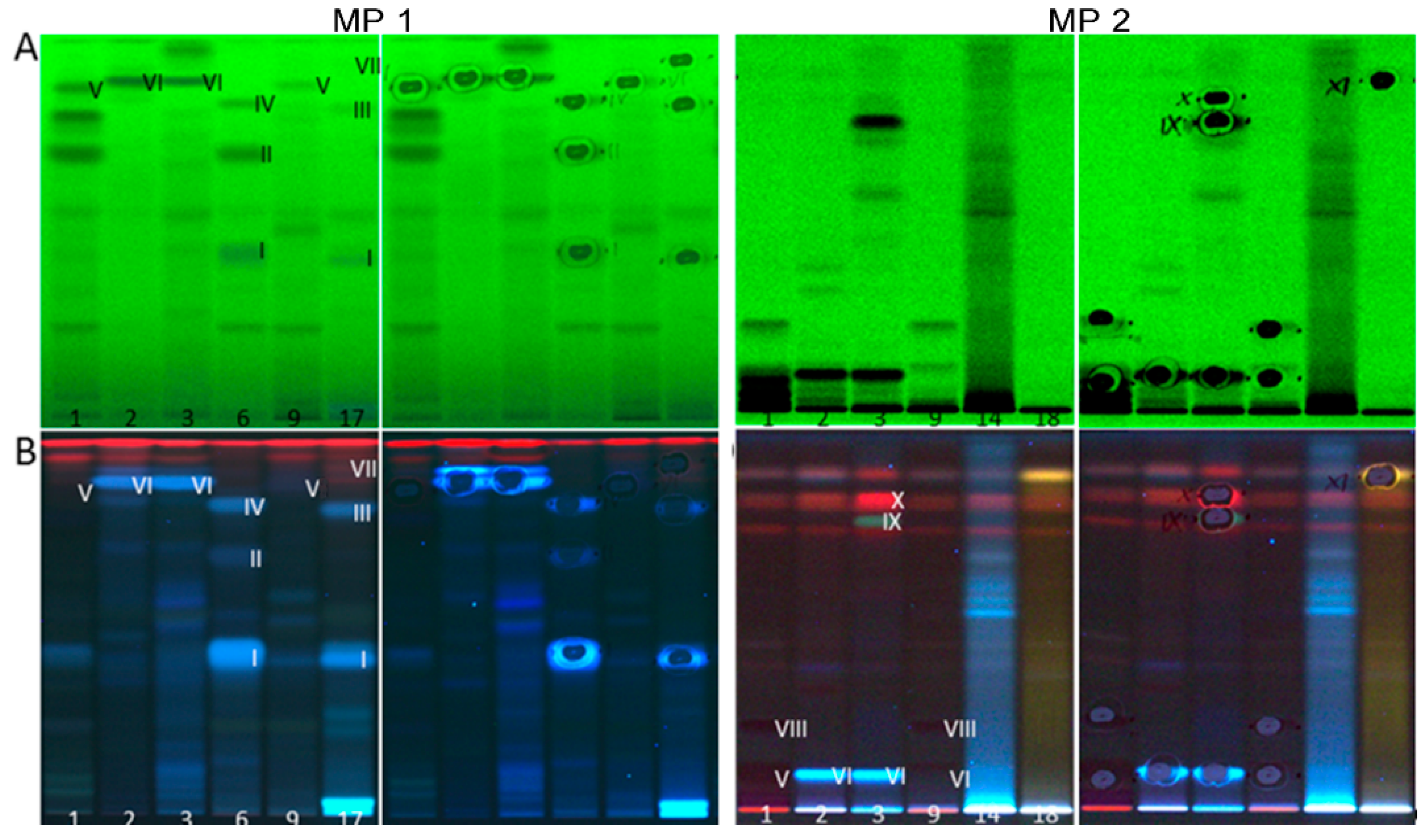
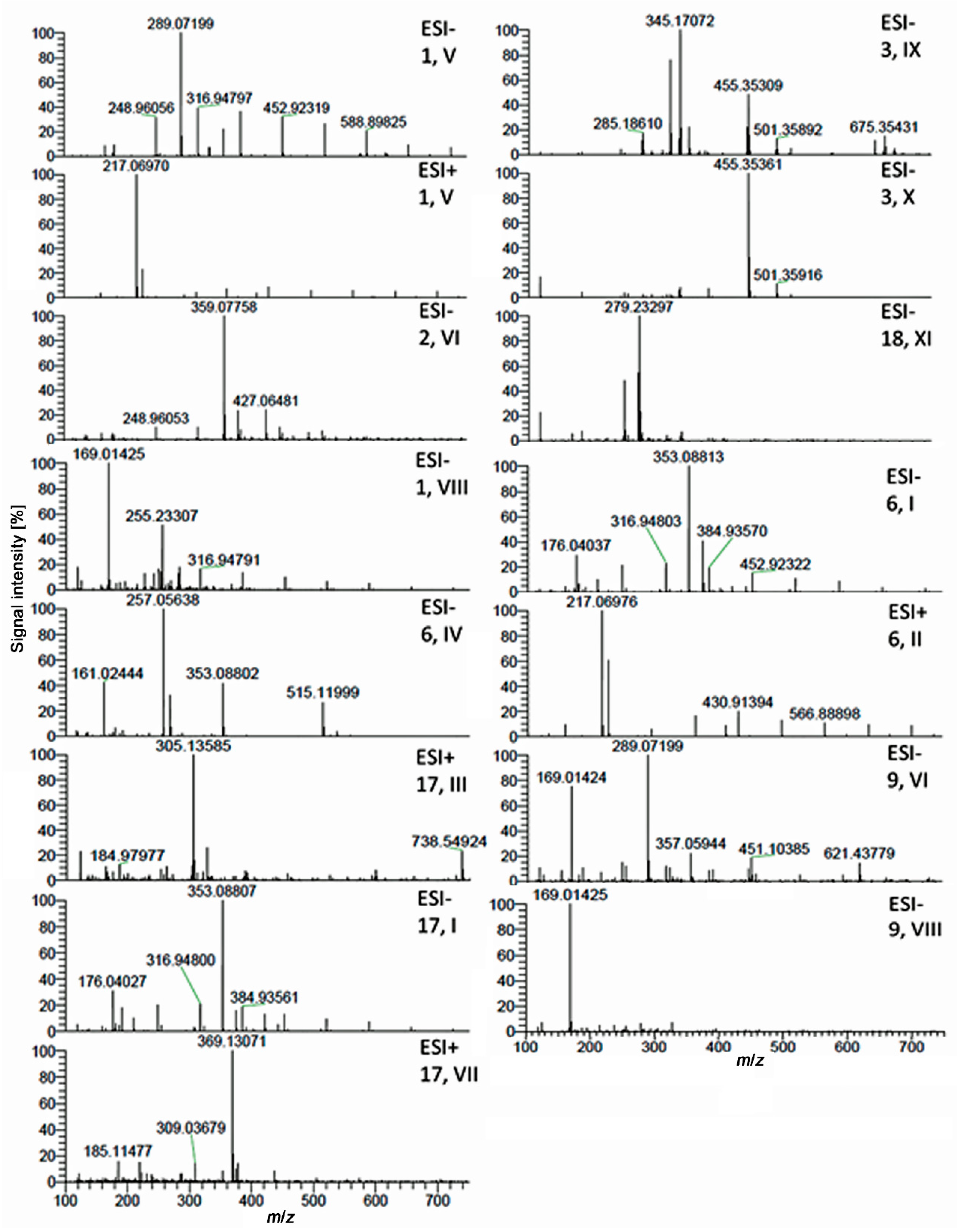
2.3. Characterization of Active Zones I–XI by HPTLC–HESI–HRMS
2.4. Outlook
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Production of Fortified Plant Powders
3.3. Extraction of the Plant Powders
3.4. HPTLC–UV/Vis/FLD Method
3.5. Neutralization of Acidic Traces
3.6. Effect-Directed Profiling
3.6.1. HPTLC–A. fischeri Bioassay
3.6.2. HPTLC–α-Glucosidase Inhibition Assay
3.6.3. HPTLC–β-Glucosidase Inhibition Assay
3.6.4. HPTLC–AChE Inhibition Assay
3.6.5. HPTLC–Tyrosinase Inhibition Assay
3.6.6. HPTLC–β-Glucuronidase Inhibition Assay
3.7. HPTLC–HRMS Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Williamson, E. Synergy and other interactions in phytomedicines. Phytomedicine 2001, 8, 401–409. [Google Scholar] [CrossRef]
- Verpoorte, R.; Kim, H.K.; Choi, Y.H. Synergy: Easier to say than to prove. Synergy 2018, 7, 34–35. [Google Scholar] [CrossRef]
- Morlock, G.E.; Heil, J. HI-HPTLC-UV/Vis/FLD-HESI-HRMS and bioprofiling of steviol glycosides, steviol, and isosteviol in Stevia leaves and foods. Anal. Bioanal. Chem. 2020, 412, 6431–6448. [Google Scholar] [CrossRef] [Green Version]
- Guinobert, I.; Bardot, V.; Dubourdeaux, M. De la plante aux effets biologiques de l’extrait: Quand la démarche scientifique éclaire les usages. Phytothérapie 2019, 17, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Bardot, V.; Escalon, A.; Ripoche, I.; Denis, S.; Alric, M.; Chalancon, S.; Chalard, P.; Cotte, C.; Berthomier, L.; Leremboure, M.; et al. Benefits of the ipowder® extraction process applied to Melissa officinalis L: Improvement of antioxidant activity and in vitro gastro-intestinal release profile of rosmarinic acid. Food Funct. 2020, 11, 722–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morlock, G.E.; Heil, J.; Inarejos-Garcia, A.M.; Maeder, J. Effect-directed profiling of powdered tea extracts for catechins, theaflavins, flavonols and caffeine. Antioxidants 2021, 10, 117. [Google Scholar] [CrossRef] [PubMed]
- Chandana, N.G.A.S.S.; Morlock, G.E. Eight different bioactivity profiles of 40 cinnamon extracts to discover multipotent compounds by multi-imaging planar chromatography hyphenated with effect-directed analysis and high-resolution mass spectrometry. Food Chem. 2021. in print. [Google Scholar]
- Dubourdeaux, M. Procédé de Préparation d’extraits Végétaux Permettant l’obtention d’une Nouvelle Forme Galénique. 14 January 2009. Available online: https://patents.google.com/patent/EP2080436A2/en (accessed on 21 January 2021).
- Móricz, Á.M.; Ott, P.G.; Morlock, G.E. Discovered acetylcholinesterase inhibition and antibacterial activity of polyacetylenes in tansy root extract via effect-directed chromatographic fingerprints. J. Chromatogr. A 2018, 1543, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Krüger, S.; Bergin, A.; Morlock, G.E. Effect-directed analysis of ginger (Zingiber officinale) and its food products, and quantification of bioactive compounds via high-performance thin-layer chromatography and mass spectrometry. Food Chem. 2018, 243, 258–268. [Google Scholar] [CrossRef]
- Klöppel, A.; Grasse, W.; Brümmer, F.; Morlock, G. HPTLC coupled with bioluminescence and mass spectrometry for bioactivity-based analysis of secondary metabolites in marine sponges. J. Planar Chromatogr. Mod. TLC 2008, 21, 431–436. [Google Scholar] [CrossRef]
- Krüger, S.; Hüsken, L.; Fornasari, R.; Scainelli, I.; Morlock, G.E. Effect-directed fingerprints of 77 botanical extracts via a generic high-performance thin-layer chromatography method combined with assays and mass spectrometry. J. Chromatogr. A 2017, 1529, 93–106. [Google Scholar] [CrossRef]
- DIN EN ISO 11348-1. Water Quality—Determination of the Inhibitory Effect of Water Samples on the Light Emission of Vibrio fischeri (Luminescent bacteria test)—Part 1: Method Using Freshly Prepared Bacteria; Beuth Verlag: Berlin, Germany, 2009. [Google Scholar]
- Bartmańska, A.; Wałecka-Zacharska, E.; Tronina, T.; Popłoński, J.; Sordon, S.; Brzezowska, E.; Bania, J.; Huszcza, E. Antimicrobial Properties of Spent Hops Extracts, Flavonoids Isolated Therefrom, and Their Derivatives. Molecules 2018, 23, 2059. [Google Scholar] [CrossRef] [Green Version]
- Karabín, M.; Hudcová, T.; Jelínek, L.; Dostálek, P. Biologically Active Compounds from Hops and Prospects for Their Use. Compr. Rev. Food Sci. Food Saf. 2016, 15, 542–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Shin, D.-S.; Oh, K.-B.; Shin, K.H. Antibacterial compounds from the leaves of Acanthopanax senticosus. Arch. Pharm. Res. 2003, 26, 40–42. [Google Scholar] [CrossRef] [PubMed]
- Davydov, M.; Krikorian, A.D. Eleutherococcus senticosus (Rupr. & Maxim.) Maxim. (Araliaceae) as an adaptogen: A closer look. J. Ethnopharmacol. 2000, 72, 345–393. [Google Scholar] [CrossRef] [PubMed]
- Masadeh, M.M.; Alkofahi, A.S.; Alzoubi, K.H.; Tumah, H.N.; Bani-Hani, K. Anti-Helicobactor pylori activity of some Jordanian medicinal plants. Pharm. Biol. 2014, 52, 566–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Jain, L.; Pandey, M.B.; Singh, U.P.; Pandey, V.B. Antifungal activity of the alkaloids from Eschscholzia californica. Folia Microbiol. (Praha) 2009, 54, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Smither-Kopperl, M.L. Plant Guide for California Poppy (Eschscholzia californica), Lockeford, CA 95237. 2018. Available online: https://plants.usda.gov/plantguide/pdf/pg_esca2.pdf (accessed on 21 January 2021).
- Kuete, V. Health Effects of Alkaloids from African Medicinal Plants. In Toxicological Survey of African Medicinal Plants; Kuete, V., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 611–633. ISBN 9780128000182. [Google Scholar]
- Taylor, P.W. Interactions of Tea-Derived Catechin Gallates with Bacterial Pathogens. Molecules 2020, 25, 1986. [Google Scholar] [CrossRef]
- Gopal, J.; Muthu, M.; Paul, D.; Kim, D.-H.; Chun, S. Bactericidal activity of green tea extracts: The importance of catechin containing nano particles. Sci. Rep. 2016, 6, 266. [Google Scholar] [CrossRef]
- Oliveira, D.A.; Salvador, A.A.; Smânia, A.; Smânia, E.F.A.; Maraschin, M.; Ferreira, S.R.S. Antimicrobial activity and composition profile of grape (Vitis vinifera) pomace extracts obtained by supercritical fluids. J. Biotechnol. 2013, 164, 423–432. [Google Scholar] [CrossRef]
- Ehsani, A.; Alizadeh, O.; Hashemi, M.; Afshari, A.; Aminzare, M. Phytochemical, antioxidant and antibacterial properties of Melissa officinalis and Dracocephalum moldavica essential oils. Vet. Res. Forum 2017, 8, 223–229. [Google Scholar] [PubMed]
- Thompson, A.; Meah, D.; Ahmed, N.; Conniff-Jenkins, R.; Chileshe, E.; Phillips, C.O.; Claypole, T.C.; Forman, D.W.; Row, P.E. Comparison of the antibacterial activity of essential oils and extracts of medicinal and culinary herbs to investigate potential new treatments for irritable bowel syndrome. BMC Complement. Altern. Med. 2013, 13, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, C.; Barros, L.; José Alves, M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Artichoke and milk thistle pills and syrups as sources of phenolic compounds with antimicrobial activity. Food Funct. 2016, 7, 3083–3090. [Google Scholar] [CrossRef] [Green Version]
- Rauha, J.P.; Remes, S.; Heinonen, M.; Hopia, A.; Kähkönen, M.; Kujala, T.; Pihlaja, K.; Vuorela, H.; Vuorela, P. Antimicrobial effects of Finnish plant extracts containing flavonoids and other phenolic compounds. Int. J. Food Microbiol. 2000, 56, 3–12. [Google Scholar] [CrossRef]
- French, K.E.; Harvey, J.; McCullagh, J.S.O. Targeted and Untargeted Metabolic Profiling of Wild Grassland Plants identifies Antibiotic and Anthelmintic Compounds Targeting Pathogen Physiology, Metabolism and Reproduction. Sci. Rep. 2018, 8, 1695. [Google Scholar] [CrossRef] [Green Version]
- Deliorman Orhan, D.; Orhan, N.; Özçelik, B.; Ergun, F. Biological activities of Vitis vinifera L. leaves. Turk. J. Biol. 2009, 341–348. [Google Scholar]
- Sharifi-Rad, M.; Mnayer, D.; Morais-Braga, M.F.B.; Carneiro, J.N.P.; Bezerra, C.F.; Coutinho, H.D.M.; Salehi, B.; Martorell, M.; Del Mar Contreras, M.; Soltani-Nejad, A.; et al. Echinacea plants as antioxidant and antibacterial agents: From traditional medicine to biotechnological applications. Phytother. Res. 2018, 32, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Raudsepp, P.; Koskar, J.; Anton, D.; Meremäe, K.; Kapp, K.; Laurson, P.; Bleive, U.; Kaldmäe, H.; Roasto, M.; Püssa, T. Antibacterial and antioxidative properties of different parts of garden rhubarb, blackcurrant, chokeberry and blue honeysuckle. J. Sci. Food Agric. 2019, 99, 2311–2320. [Google Scholar] [CrossRef]
- Kungel, P.T.A.N.; Correa, V.G.; Corrêa, R.C.G.; Peralta, R.A.; Soković, M.; Calhelha, R.C.; Bracht, A.; Ferreira, I.C.F.R.; Peralta, R.M. Antioxidant and antimicrobial activities of a purified polysaccharide from yerba mate (Ilex paraguariensis). Int. J. Biol. Macromol. 2018, 114, 1161–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhao, J.; Liu, H.; Zhou, L.; Liu, Z.; Wang, J.; Han, J.; Yu, Z.; Yang, F. Chemical analysis and biological activity of the essential oils of two valerianaceous species from China: Nardostachys chinensis and Valeriana officinalis. Molecules 2010, 15, 6411–6422. [Google Scholar] [CrossRef] [Green Version]
- Parizadeh, H.; Garampalli, R.H. Evaluation of Some Lichen Extracts for β-Glucosidase Inhibitory as a Possible Source of Herbal Anti-diabetic Drugs. Am. J. Biochem. 2016, 6, 46–50. [Google Scholar]
- Chokki, M.; Cudălbeanu, M.; Zongo, C.; Dah-Nouvlessounon, D.; Ghinea, I.O.; Furdui, B.; Raclea, R.; Savadogo, A.; Baba-Moussa, L.; Avamescu, S.M.; et al. Exploring Antioxidant and Enzymes (A-Amylase and B-Glucosidase) Inhibitory Activity of Morinda lucida and Momordica charantia Leaves from Benin. Foods 2020, 9, 434. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Kong, F. Evaluation of the in vitro α-glucosidase inhibitory activity of green tea polyphenols and different tea types. J. Sci. Food Agric. 2016, 96, 777–782. [Google Scholar] [CrossRef]
- Yilmazer-Musa, M.; Griffith, A.M.; Michels, A.J.; Schneider, E.; Frei, B. Grape seed and tea extracts and catechin 3-gallates are potent inhibitors of α-amylase and α-glucosidase activity. J. Agric. Food Chem. 2012, 60, 8924–8929. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.-C.; Shen, H.-D.; Fang, J.-J.; Ding, T.-M.; Ding, X.-P.; Liu, J.-F. On-line high-performance liquid chromatography coupled with biochemical detection method for screening of α-glucosidase inhibitors in green tea. Biomed. Chromatogr. 2018, 32, e4281. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. Meadowsweet Teas as New Functional Beverages: Comparative Analysis of Nutrients, Phytochemicals and Biological Effects of Four Filipendula Species. Molecules 2016, 22, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.I.; Vattem, D.A.; Shetty, K. Evaluation of clonal herbs of Lamiaceae species for management of diabetes and hypertension. Asia Pac. J. Clin. Nutr. 2006, 15, 107–118. [Google Scholar] [PubMed]
- Ranilla, L.G.; Kwon, Y.-I.; Apostolidis, E.; Shetty, K. Phenolic compounds, antioxidant activity and in vitro inhibitory potential against key enzymes relevant for hyperglycemia and hypertension of commonly used medicinal plants, herbs and spices in Latin America. Bioresour. Technol. 2010, 101, 4676–4689. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.S.; Escher, G.B.; Vieira do Carmo, M.; Azevedo, L.; Boscacci Marques, M.; Daguer, H.; Molognoni, L.; Inés Genovese, M.; Wen, M.; Zhang, L.; et al. A new analytical concept based on chemistry and toxicology for herbal extracts analysis: From phenolic composition to bioactivity. Food Res. Int. 2020, 132, 109090. [Google Scholar] [CrossRef]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P.; Hernández, F. Antidiabetic, Anticholinesterase and Antioxidant Activity vs. Terpenoids and Phenolic Compounds in Selected New Cultivars and Hybrids of Artichoke Cynara scolymus L. Molecules 2019, 24, 1222. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yin, H.; Liu, G.; Dong, J.; Qian, Z.; Miao, J. Xanthohumol, a prenylated chalcone from beer hops, acts as an α-glucosidase inhibitor in vitro. J. Agric. Food Chem. 2014, 62, 5548–5554. [Google Scholar] [CrossRef]
- Baranowska-Wójcik, E.; Szwajgier, D.; Winiarska-Mieczan, A. Regardless of the Brewing Conditions, Various Types of Tea are a Source of Acetylcholinesterase Inhibitors. Nutrients 2020, 12, 709. [Google Scholar] [CrossRef] [Green Version]
- Dastmalchi, K.; Ollilainen, V.; Lackman, P.; Boije af Gennäs, G.; Dorman, H.J.D.; Järvinen, P.P.; Yli-Kauhaluoma, J.; Hiltunen, R. Acetylcholinesterase inhibitory guided fractionation of Melissa officinalis L. Bioorg. Med. Chem. 2009, 17, 867–871. [Google Scholar] [CrossRef]
- Ozarowski, M.; Mikolajczak, P.L.; Bogacz, A.; Gryszczynska, A.; Kujawska, M.; Jodynis-Liebert, J.; Piasecka, A.; Napieczynska, H.; Szulc, M.; Kujawski, R.; et al. Rosmarinus officinalis L. leaf extract improves memory impairment and affects acetylcholinesterase and butyrylcholinesterase activities in rat brain. Fitoterapia 2013, 91, 261–271. [Google Scholar] [CrossRef]
- Santos, E.C.S.; Bicca, M.A.; Blum-Silva, C.H.; Costa, A.P.R.; Dos Santos, A.A.; Schenkel, E.P.; Farina, M.; Reginatto, F.H.; de Lima, T.C.M. Anxiolytic-like, stimulant and neuroprotective effects of Ilex paraguariensis extracts in mice. Neuroscience 2015, 292, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Neagu, E.; Paun, G.; Albu, C.; Radu, G.-L. Assessment of acetylcholinesterase and tyrosinase inhibitory and antioxidant activity of Alchemilla vulgaris and Filipendula ulmaria extracts. J. Taiwan Inst. Chem. Eng. 2015, 52, 1–6. [Google Scholar] [CrossRef]
- Salazar, P.B.; Minahk, C.; Rodriguez-Vaquero, M.J. Polyphenols from Cafayate Grape Pomace Display High Inhibitory Activity on Human Acetylcholinesterase. Available online: https://www.asev.org/abstract/polyphenols-cafayate-grape-pomace-display-high-inhibitory-activity-human (accessed on 16 December 2020).
- Cahlíková, L.; Macáková, K.; Kunes, J.; Kurfürst, M.; Opletal, L.; Cvacka, J.; Chlebek, J.; Blundene, G. Acetylcholinesterase and butyrylcholinesterase inhibitory compounds from Eschscholzia californica (Papaveraceae). Nat. Prod. Commun. 2010, 5, 1035–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- No, J.K.; Soung, D.Y.; Kim, Y.J.; Shim, K.H.; Jun, Y.S.; Rhee, S.H.; Yokozawa, T.; Chung, H.Y. Inhibition of tyrosinase by green tea components. Life Sci. 1999, 65, PL241-6. [Google Scholar] [CrossRef]
- Jun, H.-J.; Roh, M.; Kim, H.W.; Houng, S.-J.; Cho, B.; Yun, E.J.; Hossain, M.A.; Lee, H.; Kim, K.H.; Lee, S.-J. Dual inhibitions of lemon balm (Melissa officinalis) ethanolic extract on melanogenesis in B16-F1 murine melanocytes: Inhibition of tyrosinase activity and its gene expression. Food Sci. Biotechnol. 2011, 20, 1051–1059. [Google Scholar] [CrossRef]
- Al-Mamary, M. The antioxidant and tyrosinase inhibitory activities of some essential oils obtained from aromatic plants grown and used in Yemen. Sci. Res. Essays 2011, 6. [Google Scholar] [CrossRef]
- Ferri, M.; Rondini, G.; Calabretta, M.M.; Michelini, E.; Vallini, V.; Fava, F.; Roda, A.; Minnucci, G.; Tassoni, A. White grape pomace extracts, obtained by a sequential enzymatic plus ethanol-based extraction, exert antioxidant, anti-tyrosinase and anti-inflammatory activities. N. Biotechnol. 2017, 39, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Molan, A.-L.; Liu, Z.; Tiwari, R. The ability of green tea to positively modulate key markers of gastrointestinal function in rats. Phytother. Res. 2010, 24, 1614–1619. [Google Scholar] [CrossRef]
- Molan, A.-L.; Liu, Z.; Plimmer, G. Evaluation of the effect of blackcurrant products on gut microbiota and on markers of risk for colon cancer in humans. Phytother. Res. 2014, 28, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Shao, H.; Zhou, Z.; Wang, Q.; Zhao, L.; Yang, X. Non-extractable polyphenols of green tea and their antioxidant, anti-α-glucosidase capacity, and release during in vitro digestion. J. Funct. Foods 2018, 42, 129–136. [Google Scholar] [CrossRef]
- Guyot, S.; Pellerin, P.; Brillouet, J.M.; Cheynier, V. Inhibition of β-Glucosidase (Amygdalae dulces) by (+)-Catechin Oxidation Products and Procyanidin Dimers. Biosci. Biotechnol. Biochem. 1996, 60, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Fu, X.-W.; Dai, X.-L.; Hua, F.; Chu, G.-X.; Chu, M.-J.; Hu, F.-L.; Ling, T.-J.; Gao, L.-P.; Xie, Z.-W.; et al. Novel acetylcholinesterase inhibitors from Zijuan tea and biosynthetic pathway of caffeoylated catechin in tea plant. Food Chem. 2017, 237, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Kavak, D.D.; Altıok, E.; Bayraktar, O.; Ülkü, S. Pistacia terebinthus extract: As a potential antioxidant, antimicrobial and possible β-glucuronidase inhibitor. J. Mol. Catal. B Enzym. 2010, 64, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Oboh, G.; Ogunsuyi, O.B.; Ogunbadejo, M.D.; Adefegha, S.A. Influence of gallic acid on α-amylase and α-glucosidase inhibitory properties of acarbose. J. Food Drug Anal. 2016, 24, 627–634. [Google Scholar] [CrossRef]
- Li, P.-H.; Lin, Y.-W.; Lu, W.-C.; Hu, J.-M.; Huang, D.-W. In Vitro Hypoglycemic Activity of the Phenolic Compounds in Longan Fruit (Dimocarpus longan var. Fen Ke) Shell Against α-Glucosidase and β-Galactosidase. Int. J. Food Prop. 2015, 19, 1786–1797. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Randhawa, K.; Singh, V.; Shri, R. Bioactivity-Guided Isolation of Acetylcholinesterase Inhibitor from Ganoderma mediosinense (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 755–763. [Google Scholar] [CrossRef]
- Su, T.-R.; Lin, J.-J.; Tsai, C.-C.; Huang, T.-K.; Yang, Z.-Y.; Wu, M.-O.; Zheng, Y.-Q.; Su, C.-C.; Wu, Y.-J. Inhibition of Melanogenesis by Gallic Acid: Possible Involvement of the PI3K/Akt, MEK/ERK and Wnt/β-Catenin Signaling Pathways in B16F10 Cells. Int. J. Mol. Sci. 2013, 14, 20443–20458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Wang, J.; Takano, H.; Xu, Z.; Nishiwaki, H.; Yonekura, L.; Yang, R.; Tamura, H. Rosmarinic acid and its ester derivatives for enhancing antibacterial, α-glucosidase inhibitory, and lipid accumulation suppression activities. J. Food Biochem. 2019, 43, e12719. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.S.; Hossain, M.A.; Said, S.A. Isolation and characterization of antimicrobial compound from the stem-bark of the traditionally used medicinal plant Adenium obesum. J. Tradit. Complement. Med. 2017, 7, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Matejczyk, M.; Świsłocka, R.; Golonko, A.; Lewandowski, W.; Hawrylik, E. Cytotoxic, genotoxic and antimicrobial activity of caffeic and rosmarinic acids and their lithium, sodium and potassium salts as potential anticancer compounds. Adv. Med. Sci. 2018, 63, 14–21. [Google Scholar] [CrossRef]
- Abedini, A.; Roumy, V.; Mahieux, S.; Biabiany, M.; Standaert-Vitse, A.; Rivière, C.; Sahpaz, S.; Bailleul, F.; Neut, C.; Hennebelle, T. Rosmarinic Acid and Its Methyl Ester as Antimicrobial Components of the Hydromethanolic Extract of Hyptis atrorubens Poit. (Lamiaceae). Evid. Based Complement. Altern. Med. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jordán, M.J.; Lax, V.; Rota, M.C.; Lorán, S.; Sotomayor, J.A. Relevance of carnosic acid, carnosol, and rosmarinic acid concentrations in the in vitro antioxidant and antimicrobial activities of Rosmarinus officinalis (L.) methanolic extracts. J. Agric. Food Chem. 2012, 60, 9603–9608. [Google Scholar] [CrossRef]
- Gülçin, İ.; Scozzafava, A.; Supuran, C.T.; Koksal, Z.; Turkan, F.; Çetinkaya, S.; Bingöl, Z.; Huyut, Z.; Alwasel, S.H. Rosmarinic acid inhibits some metabolic enzymes including glutathione S-transferase, lactoperoxidase, acetylcholinesterase, butyrylcholinesterase and carbonic anhydrase isoenzymes. J. Enzyme Inhib. Med. Chem. 2016, 31, 1698–1702. [Google Scholar] [CrossRef] [Green Version]
- Zuo, A.-R.; Dong, H.-H.; Yu, Y.-Y.; Shu, Q.-L.; Zheng, L.-X.; Yu, X.-Y.; Cao, S.-W. The antityrosinase and antioxidant activities of flavonoids dominated by the number and location of phenolic hydroxyl groups. Chin. Med. 2018, 13, 51. [Google Scholar] [CrossRef] [Green Version]
- Bernardes, W.A.; Lucarini, R.; Tozatti, M.G.; Souza, M.G.M.; Silva, M.L.A.; Filho, A.A.d.S.; Martins, C.H.G.; Crotti, A.E.M.; Pauletti, P.M.; Groppo, M.; et al. Antimicrobial activity of Rosmarinus officinalis against oral pathogens: Relevance of carnosic acid and carnosol. Chem. Biodivers. 2010, 7, 1835–1840. [Google Scholar] [CrossRef]
- Collins, M.A.; Charles, H.P. Antimicrobial activity of Carnosol and Ursolic acid: Two anti-oxidant constituents of Rosmarinus officinalis L. Food Microbiol. 1987, 4, 311–315. [Google Scholar] [CrossRef]
- Ma, Y.-Y.; Zhao, D.-G.; Zhang, R.; He, X.; Li, B.Q.; Zhang, X.-Z.; Wang, Z.; Zhang, K. Identification of bioactive compounds that contribute to the α-glucosidase inhibitory activity of rosemary. Food Funct. 2020, 11, 1692–1701. [Google Scholar] [CrossRef]
- Shirasugi, I.; Sakakibara, Y.; Yamasaki, M.; Nishiyama, K.; Matsui, T.; Liu, M.-C.; Suiko, M. Novel screening method for potential skin-whitening compounds by a luciferase reporter assay. Biosci. Biotechnol. Biochem. 2010, 74, 2253–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesus, J.A.; Lago, J.H.G.; Laurenti, M.D.; Yamamoto, E.S.; Passero, L.F.D. Antimicrobial activity of oleanolic and ursolic acids: An update. Evid. Based Complement. Alternat. Med. 2015, 2015, 620472. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, H.; Zhang, P.; Gao, L.; Yan, N.; Li, P.; Liu, X.; Du, Y.; Shen, G. Chemical Composition, Antioxidant Activity and α-Glucosidase Inhibitory Activity of Chaenomeles Speciosa from Four Production Areas in China. Molecules 2018, 23, 2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Hu, X.; Xu, X.; Zhang, G.; Gong, D. Inhibitory mechanism of two allosteric inhibitors, oleanolic acid and ursolic acid on α-glucosidase. Int. J. Biol. Macromol. 2018, 107, 1844–1855. [Google Scholar] [CrossRef]
- Loesche, A.; Köwitsch, A.; Lucas, S.D.; Al-Halabi, Z.; Sippl, W.; Al-Harrasi, A.; Csuk, R. Ursolic and oleanolic acid derivatives with cholinesterase inhibiting potential. Bioorg. Chem. 2019, 85, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.K.; Heo, H.J.; Kim, E.K.; Kim, H.K.; Huh, T.L.; Lim, Y.; Kim, S.K.; Shin, D.H. Inhibitory effect of ursolic acid purified from Origanum majorana L. on the acetylcholinesterase. Mol. Cells 2001, 11, 137–143. [Google Scholar]
- Han, S.K.; Kim, Y.G.; Kang, H.C.; Huh, J.R.; Kim, J.Y.; Baek, N.-I.; Lee, D.-K.; Lee, D.-G. Oleanolic acid from Fragaria ananassa calyx leads to inhibition of α-MSH-induced melanogenesis in B16-F10 melanoma cells. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 735–742. [Google Scholar] [CrossRef]
- Muñoz, E.; Avila, J.G.; Alarcón, J.; Kubo, I.; Werner, E.; Céspedes, C.L. Tyrosinase inhibitors from Calceolaria integrifolia s.l.: Calceolaria talcana aerial parts. J. Agric. Food Chem. 2013, 61, 4336–4343. [Google Scholar] [CrossRef]
- Puupponen-Pimia, R.; Nohynek, L.; Meier, C.; Kahkonen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.-M. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef]
- Pérez-Nájera, V.C.; Gutiérrez-Uribe, J.A.; Antunes-Ricardo, M.; Hidalgo-Figueroa, S.; Del-Toro-Sánchez, C.L.; Salazar-Olivo, L.A.; Lugo-Cervantes, E. Smilax aristolochiifolia Root Extract and Its Compounds Chlorogenic Acid and Astilbin Inhibit the Activity of α-Amylase and α-Glucosidase Enzymes. Evid. Based Complement. Altern. Med. 2018, 2018, 6247306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oboh, G.; Agunloye, O.M.; Adefegha, S.A.; Akinyemi, A.J.; Ademiluyi, A.O. Caffeic and chlorogenic acids inhibit key enzymes linked to type 2 diabetes (in vitro): A comparative study. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.-H.; Lee, H.-K.; Kim, J.-A.; Hong, S.-I.; Kim, H.-C.; Jo, T.-H.; Park, Y.-I.; Lee, C.-K.; Kim, Y.-B.; Lee, S.-Y.; et al. Neuroprotective effects of chlorogenic acid on scopolamine-induced amnesia via anti-acetylcholinesterase and anti-oxidative activities in mice. Eur. J. Pharmacol. 2010, 649, 210–217. [Google Scholar] [CrossRef]
- Guo, Z.; Li, J. Chlorogenic Acid Prevents Alcohol-induced Brain Damage in Neonatal Rat. Transl. Neurosci. 2017, 8, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-R.; Habasi, M.; Xie, L.-Z.; Aisa, H.A. Effect of chlorogenic acid on melanogenesis of B16 melanoma cells. Molecules 2014, 19, 12940–12948. [Google Scholar] [CrossRef] [Green Version]
- Karak, S.; Das, S.; Biswas, M.; Choudhury, A.; Dutta, M.; Chaudhury, K.; De, B. Phytochemical composition, β-glucuronidase inhibition, and antioxidant properties of two fractions of Piper betle leaf aqueous extract. J. Food Biochem. 2019, 43, e13048. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M.; Dobes, P. Caffeine inhibits acetylcholinesterase, but not butyrylcholinesterase. Int. J. Mol. Sci. 2013, 14, 9873–9882. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, X.; Ni, H.; Du, X.; Chen, F.; Jiang, Z.; Li, Q. Identification and Characterization of the Tyrosinase Inhibitory Activity of Caffeine from Camellia Pollen. J. Agric. Food Chem. 2019, 67, 12741–12751. [Google Scholar] [CrossRef]
- Rempe, C.S.; Burris, K.P.; Woo, H.L.; Goodrich, B.; Gosnell, D.K.; Tschaplinski, T.J.; Stewart, C.N.; Schuch, R. Computational Ranking of Yerba Mate Small Molecules Based on Their Predicted Contribution to Antibacterial Activity against Methicillin-Resistant Staphylococcus aureus. PLoS ONE 2015, 10, e0123925. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Ma, B.; Ge, L.; Mo, Q.; Zhou, G.; He, J.; Wang, Y. Dicaffeoylquinic Acid-Enriched Fraction of Cichorium glandulosum Seeds Attenuates Experimental Type 1 Diabetes via Multipathway Protection. J. Agric. Food Chem. 2015, 63, 10791–10802. [Google Scholar] [CrossRef]
- Kang, J.Y.; Park, S.K.; Guo, T.J.; Ha, J.S.; Du Lee, S.; Kim, J.M.; Lee, U.; Kim, D.O.; Heo, H.J. Reversal of Trimethyltin-Induced Learning and Memory Deficits by 3,5-Dicaffeoylquinic Acid. Oxid. Med. Cell. Longev. 2016, 2016, 6981595. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K.; Kishimoto, N.; Kakino, Y.; Mochida, K.; Fujita, T. In vitro antioxidative effects and tyrosinase inhibitory activities of seven hydroxycinnamoyl derivatives in green coffee beans. J. Agric. Food Chem. 2004, 52, 4893–4898. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Sun, C.; Huang, H.; Lin, W.; Gao, J.; Lin, Y.; Zhang, Z.; Huo, X.; Tian, X.; Yu, Z.; et al. β-Glucuronidase- and OATP2B1-mediated drug interaction of scutellarin in Dengzhan Xixin Injection: A formulation aspect. Drug Dev. Res. 2020, 81, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Kusumah, D.; Wakui, M.; Murakami, M.; Xie, X.; Yukihito, K.; Maeda, I. Linoleic acid, α-linolenic acid, and monolinolenins as antibacterial substances in the heat-processed soybean fermented with Rhizopus oligosporus. Biosci. Biotechnol. Biochem. 2020, 84, 1285–1290. [Google Scholar] [CrossRef]
- Artanti, N.; Tachibana, S.; Kardono, L.B.S. Effect of Media Compositions on α-glucosidase Inhibitory Activity, Growth and Fatty Acid Content in Mycelium Extracts of Colletotrichum sp. TSC13 from Taxus Sumatrana (Miq.) de Laub. Pak. J. Biol. Sci. 2014, 17, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Cwikla, C.; Schmidt, K.; Matthias, A.; Bone, K.M.; Lehmann, R.; Tiralongo, E. Investigations into the antibacterial activities of phytotherapeutics against Helicobacter pylori and Campylobacter jejuni. Phytother. Res. 2010, 24, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Denev, P.; Kratchanova, M.; Ciz, M.; Lojek, A.; Vasicek, O.; Blazheva, D.; Nedelcheva, P.; Vojtek, L.; Hyrsl, P. Antioxidant, antimicrobial and neutrophil-modulating activities of herb extracts. Acta Biochim. Pol. 2014, 61, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants. J. Clin. Med. 2019, 9, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azadniya, E.; Goldoni, L.; Bandiera, T.; Morlock, G.E. Same analytical method for both (bio)assay and zone isolation to identify/quantify bioactive compounds by quantitative nuclear magnetic resonance spectroscopy. J. Chromatogr. A 2020, 1628, 461434. [Google Scholar] [CrossRef]
- El Sohaimy, S.A. Chemical Composition, Antioxidant and Antimicrobial Potential of Artichoke. Open Nutraceuticals J. 2014, 1, 15–20. [Google Scholar] [CrossRef]
- Jamshidi-Aidji, M.; Morlock, G.E. Fast equivalency estimation of unknown enzyme inhibitors in situ the effect-directed fingerprint, shown for Bacillus lipopeptide extracts. Anal. Chem. 2018, 90, 14260–14268. [Google Scholar] [CrossRef]
- Jamshidi-Aidji, M.; Morlock, G.E. From bioprofiling and characterization to bioquantification of natural antibiotics by direct bioautography linked to high-resolution mass spectrometry: Exemplarily shown for Salvia miltiorrhiza root. Anal. Chem. 2016, 88, 10979–10986. [Google Scholar] [CrossRef]
- Klingelhöfer, I.; Morlock, G.E. Bioprofiling of surface/wastewater and bioquantitation of discovered endocrine-active compounds by streamlined direct bioautography. Anal. Chem. 2015, 87, 11098–11104. [Google Scholar] [CrossRef] [PubMed]
- Morlock, G.E.; Klingelhöfer, I. Liquid chromatography-bioassay-mass spectrometry for profiling of physiologically active food. Anal. Chem. 2014, 86, 8289–8295. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Marin-Kuan, M.; Debon, E.; Serrant, P.; Cottet-Fontannaz, C.; Schilter, B.; Morlock, G.E. Detection of low levels of genotoxic compounds in food contact materials using an alternative HPTLC-SOS-Umu-C Assay. ALTEX Altern. Anim. Exp. 2020. [Google Scholar] [CrossRef]
- Azadniya, E.; Mollergues, J.; Stroheker, T.; Billerbeck, K.; Morlock, G.E. New incorporation of the S9 system into methods for detecting acetylcholinesterase inhibition, comparison and proof-of-concept using food contact materials. Anal. Chim. Acta 2020, 1129, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Klingelhöfer, I.; Hockamp, N.; Morlock, G.E. Non-targeted detection and differentiation of agonists versus antagonists, directly in bioprofiles of everyday products. Anal. Chim. Acta 2020, 1125, 288–298. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | A. fischeri | α-Glucosidase | β-Glucosidase | AChE | Tyrosinase | β-Glucuronidase | |
|---|---|---|---|---|---|---|---|
| ID | Plant | Amount applied (µg/band): 200 | 400 | 300 | |||
| 1 | Green tea | +++ | ++++ | +++ | +++ | ++ | +++ |
| 2 | Lemon balm | ++ | ++ | +++ | ++ | ++ | +++ |
| 3 | Rosemary | ++++ | +++ | +++ | ++ | ++ | ++++ |
| 4 | Eleutherococcus | ++++ | + | - | + | - | ++ |
| 5 | Green tea | ++ | ++ | ++ | ++ | ++ | ++ |
| 6 | Yerba mate | + | ++ | +++ | ++ | ++ | +++ |
| 7 | Red vine | ++ | + | - | - | + | ++ |
| 8 | Valerian | ++ | + | - | + | + | + |
| 9 | Meadowsweet | ++ | ++ | ++ | ++ | + | +++ |
| 10 | Echinacea | +++ | + | + | + | - | - |
| 11 | Blackcurrant | ++ | + | + | - | - | + |
| 12 | Black radish | - | - | - | - | + | - |
| 13 | Horse tail | - | - | - | - | - | + |
| 14 | Hops | ++++ | ++++ | ++ | - | - | ++ |
| 15 | Grape pomace | ++++ | + | + | + | + | ++ |
| 16 | Passiflora | ++++ | + | + | - | + | ++ |
| 17 | Artichoke | +++ | ++ | ++ | + | + | ++ |
| 18 | Eschscholzia | ++++ | + | + | ++ | + | + |
| ID | Plant Extract | Band | Mass Signal, m/z | Molecular Formula | Mass Error Δ ppm | Tentative Assignment | Compound Activity Found in Assay Literature | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a | b | c | d | e | f | |||||||||
| 1 + 5 | Green tea | V | 289.07199 | [M − H]− | C15H14O6 | 2.65 | (epi)catechin | + | + | + | + | + | + | [22,23,53,59,60,61,62] |
| VIII | 169.01419 | [M − H]− | C7H6O5 | 0.54 | gallic acid | - | + | + | + | + | + | [63,64,65,66] | ||
| 2 | Lemon balm | VI | 359.07754 | [M − H]− | C18H16O8 | 2.36 | rosmarinic acid | + | + | + | + | + | + | [12,40,41,67,68,69,70,71,72,73] |
| 383.07388 | [M + Na]+ | |||||||||||||
| 405.05585 | [M + 2Na − H]+ | |||||||||||||
| 381.05942 | [M + Na − 2H]− | |||||||||||||
| 3 | Rosemary | VI | 359.07754 | [M − H]− | C18H16O8 | rosmarinic acid | + | + | + | + | + | + | [12,40,41,67,68,69,70,71,72,73] | |
| 381.05942 | [M − 2H + Na]− | |||||||||||||
| IX | 329.17612 | [M − H]− | C20H26O4 | 0.81 | carnosol | + | + | + | + | + | + | [71,74,75,76,77] | ||
| 345.17087 | [M + O − H]− | |||||||||||||
| 359.18661 | [M + O + CH2 − H]− | |||||||||||||
| 455.35339 | coeluted | |||||||||||||
| X | 455.35339 | [M − H]− | C30H48O3 | 0.70 | oleanolic/ursolic acid | + | + | + | + | + | + | [75,78,79,80,81,82,83,84] | ||
| 6 | Yerba mate | I | 353.08807 | [M − H]− | C16H18O9 | 0.81 | chlorogenic acid | + | + | + | + | + | + | [85,86,87,88,89,90,91] |
| 375.06992 | [M − 2H + Na]− | |||||||||||||
| II | 217.06967 | [M + Na]+ | C8H10O2N4 | 0.48 | caffeine | + | + | + | + | + | + | [92,93] | ||
| 411.15007 | [2M + Na]+ | |||||||||||||
| IV | 257.05630 | [M − 2H]2− | C25H24O12 | |||||||||||
| 353.08801 | [M − C9H6O3 − H]− | C16H18O9 | fragment: chlorogenic acid | |||||||||||
| 515.11999 | [M − H]− | C25H24O12 | 1.11 | dicaffeoylquinic acid | + | + | + | + | + | + | [94,95,96,97,98] | |||
| 537.10193 | [M + Na − 2H]− | |||||||||||||
| 9 | Meadowsweet | V | 289.07199 | [M − H]− | C16H9O2N4 | 1.97 | ||||||||
| C15H14O6 | 2.65 | (epi)catechin | + | + | + | + | + | + | [22,23,53,59,60,61,62] | |||||
| VIII | 169.01425 | [M − H]− | C7H6O5 | gallic acid | - | + | + | + | + | + | [63,64,65,66] | |||
| 17 | Artichoke | I | 353.08807 | [M − H]− | C16H18O9 | 0.81 | chlorogenic acid | + | + | + | + | + | + | [85,86,87,88,89,90,91] |
| III | 307.06430 | [M − 2H]2- | ||||||||||||
| 615.13568 | [M − H]− | |||||||||||||
| 305.13589 | [M + Na]+ | C15H22O5 | 2.11 | cynaratriol | + | + | + | + | + | + | ||||
| VII | 369.13070 | [M + H]+ | C17H20O9 | 34 * | 3-O-feruloyl quinic acid | + | + | + | - | + | + | |||
| 18 | Eschscholzia | XI | 279.23296 277.21747 | [M − H]− [M − H]− | C18H32O2 C18H30O2 | 1.99 | linoleic acid and linolenic acid ** | + | + | + | + | + | + | [99,100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morlock, G.E.; Heil, J.; Bardot, V.; Lenoir, L.; Cotte, C.; Dubourdeaux, M. Effect-Directed Profiling of 17 Different Fortified Plant Extracts by High-Performance Thin-Layer Chromatography Combined with Six Planar Assays and High-Resolution Mass Spectrometry. Molecules 2021, 26, 1468. https://doi.org/10.3390/molecules26051468
Morlock GE, Heil J, Bardot V, Lenoir L, Cotte C, Dubourdeaux M. Effect-Directed Profiling of 17 Different Fortified Plant Extracts by High-Performance Thin-Layer Chromatography Combined with Six Planar Assays and High-Resolution Mass Spectrometry. Molecules. 2021; 26(5):1468. https://doi.org/10.3390/molecules26051468
Chicago/Turabian StyleMorlock, Gertrud E., Julia Heil, Valérie Bardot, Loïc Lenoir, César Cotte, and Michel Dubourdeaux. 2021. "Effect-Directed Profiling of 17 Different Fortified Plant Extracts by High-Performance Thin-Layer Chromatography Combined with Six Planar Assays and High-Resolution Mass Spectrometry" Molecules 26, no. 5: 1468. https://doi.org/10.3390/molecules26051468
APA StyleMorlock, G. E., Heil, J., Bardot, V., Lenoir, L., Cotte, C., & Dubourdeaux, M. (2021). Effect-Directed Profiling of 17 Different Fortified Plant Extracts by High-Performance Thin-Layer Chromatography Combined with Six Planar Assays and High-Resolution Mass Spectrometry. Molecules, 26(5), 1468. https://doi.org/10.3390/molecules26051468