
Zinc Oxide Nanoparticles and Zinc Sulfate Impact Physiological Parameters and Boosts Lipid Peroxidation in Soil Grown Coriander Plants (Coriandrum sativum)

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zinc-Based Materials and Coriander Seeds
Characteristics of the ZnO NPs Used in this Experiment
2.2. Preparation of Zinc-Based Solutions/Suspensions and Their Application to the Soil
2.3. Plant Growth Conditions
2.4. Plant Harvest
2.5. Elemental Quantification of Zn by Induction Plasma Coupled Atomic Emission Spectrometry (ICP-AES)
2.6. Chlorophyll and Carotenoid Pigment Content
2.7. H2O2 and MDA Content
2.8. Antioxidant Enzyme Activity
2.8.1. Enzyme Extraction
2.8.2. POD Activity
2.8.3. APX Activity
2.8.4. CAT Activity
2.9. Statistical Analysis
3. Results and Discussion
3.1. ZnO NPs Characterization
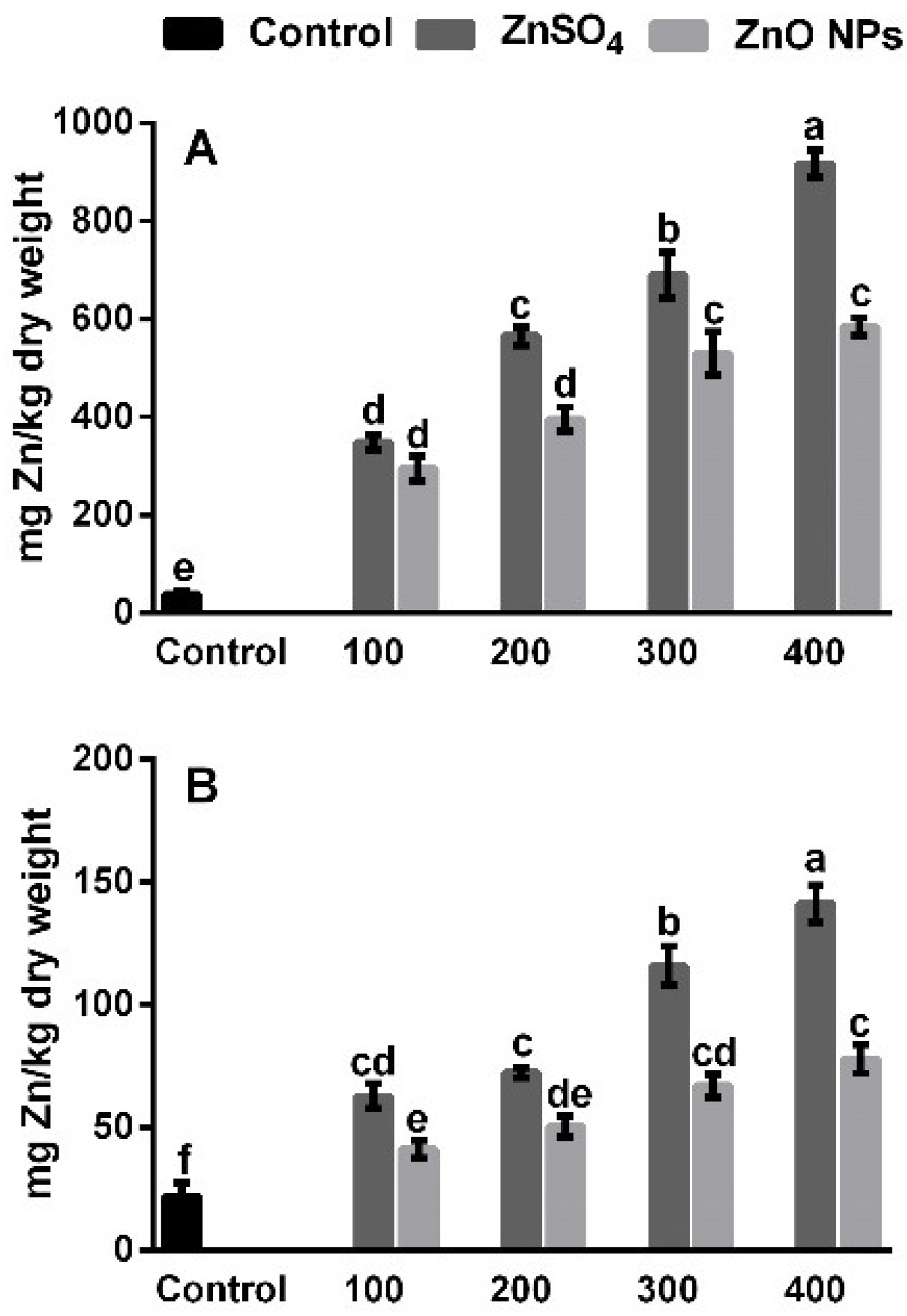
3.2. Zinc Uptake in Root and Shoot
3.3. Chlorophyll and Carotenoid Content
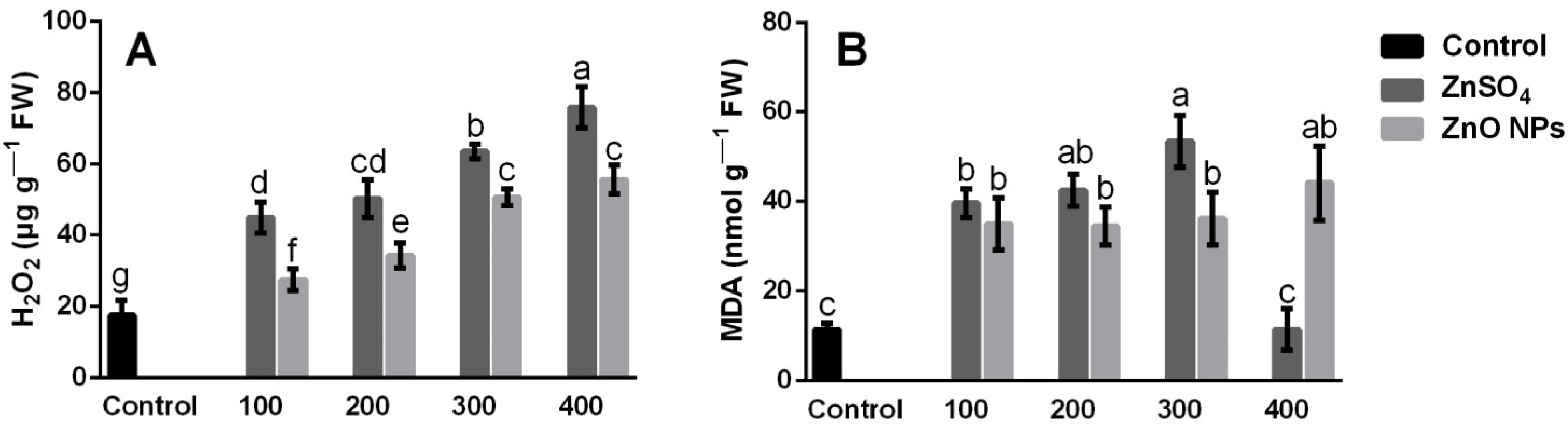
3.4. H2O2 and MDA Content
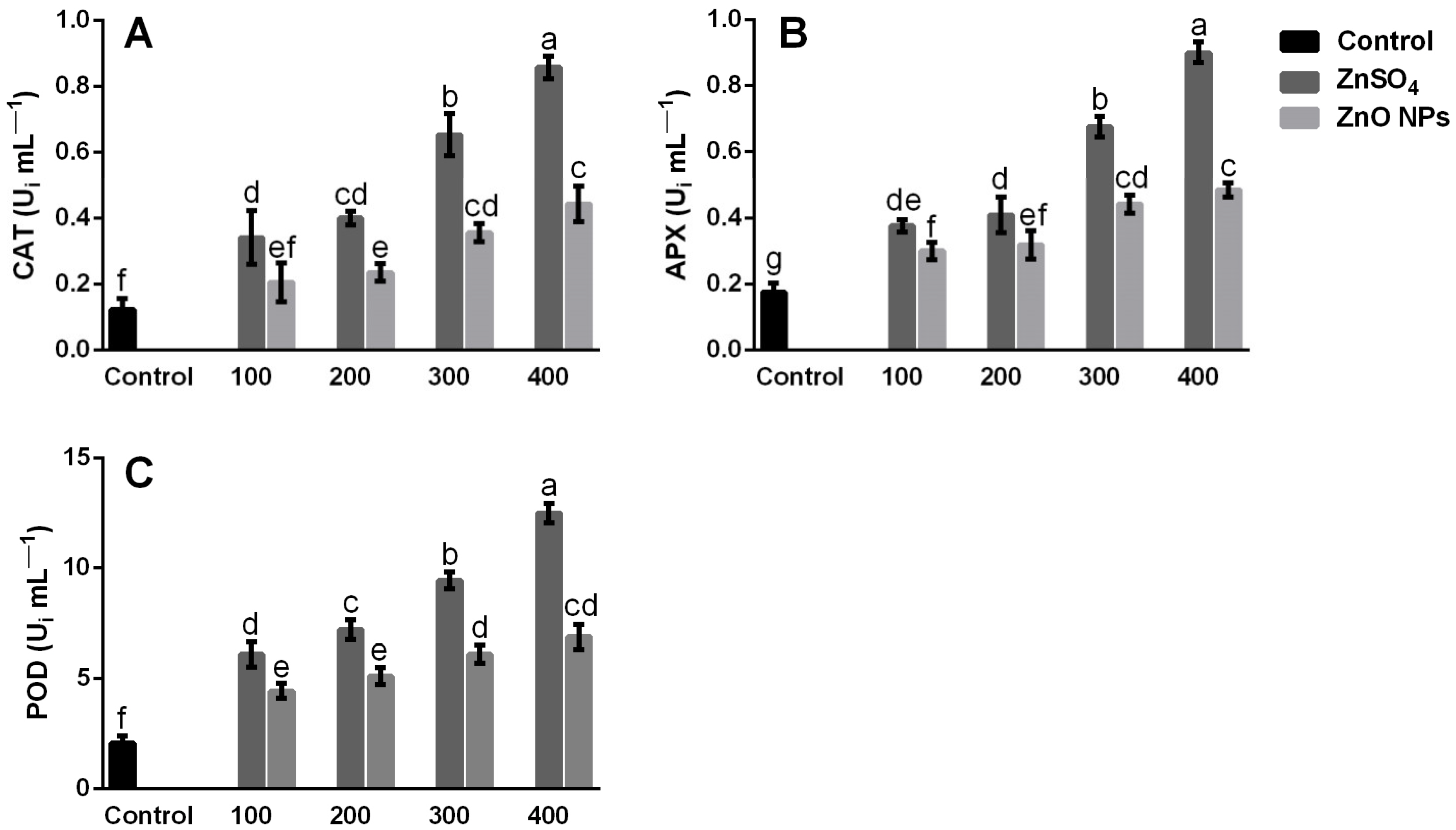
3.5. Antioxidant Enzyme Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Rossi, L.; Fedenia, L.N.; Sharifan, H.; Ma, X.; Lombardini, L. Effects of foliar application of zinc sulfate and zinc nanoparticles in coffee (Coffea arabica L.) plants. Plant Physiol. Biochem. 2019, 135, 160–166. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef] [Green Version]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.T.; Hayat, S. Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants. Photosynthetica 2017, 56, 678–686. [Google Scholar] [CrossRef]
- Rawat, P.S.; Kumar, R.; Ram, P.; Pandey, P. Effect of nanoparticles on wheat seed germination and seedling growth. Int. J. Agric. Biol. 2018, 12, 13–16. [Google Scholar] [CrossRef]
- Chikkanna, M.M.; Neelagund, S.E.; Rajashekarappa, K. Green synthesis of zinc oxide nanoparticles (ZnO NPs) and their biological activity. SN Appl. Sci. 2019, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gudkov, S.V.; Burmistrov, D.E.; Serov, D.A.; Rebezov, M.B.; Semenova, A.A.; Lisitsyn, A.B. A Mini Review of Antibacterial properties of ZnO nanoparticles. Front. Phys. 2021, 9, 641481. [Google Scholar] [CrossRef]
- Pullagurala, V.L.R.; Adisa, I.O.; Rawat, S.; Kalagara, S.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. ZnO nanoparticles increase photosynthetic pigments and decrease lipid peroxidation in soil grown cilantro (Coriandrum sativum). Plant Physiol. Biochem. 2018, 132, 120–127. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; Wang, Y.; Gao, Y.; Zhang, Z.; Xu, J.; Xing, G. Comparative physiological and metabolomic analyses revealed that foliar spraying with zinc oxide and silica nanoparticles modulates metabolite profiles in cucumber (Cucumis sativus L.). Food Energy Secur. 2020, 10, 269. [Google Scholar] [CrossRef]
- Kalal, P.R.; Jajoo, A. Priming with zinc oxide nanoparticles improve germination and photosynthetic performance in wheat. Plant Physiol. Biochem. 2021, 160, 341–351. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Chen, S.; Li, Q.; Wang, W.; Hou, C.; Gao, X.; Wang, L.I.; Wang, S. Zinc oxide nanoparticles affect biomass accumulation and photosynthesis in Arabidopsis. Front. Plant Sci. 2016, 6, 1243. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, S.; Kim, S.; Lee, I. Assessment of phytotoxicity of ZnO NPs on a medicinal plant, Fagopyrum esculentum. Environ. Sci. Pollut. Res. 2013, 20, 848–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Gómez, C.; Obrador, A.; González, D.; Babín, M.; Fernández, M.D. Comparative effect of ZnO NPs, ZnO bulk and ZnSO4 in the antioxidant defences of two plant species growing in two agricultural soils under greenhouse conditions. Sci. Total Environ. 2017, 589, 11–24. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Mishra, R.K.; Singh, S.; Singh, S.; Singh, V.P.; Singh, P.K.; Pandey, A.C. Nitric oxide ameliorates zinc oxide nanoparticles phytotoxicity in wheat seedlings: Implication of the ascorbate-glutathione cycle. Front. Plant. Sci. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.I.; Lira-Saldivar, R.H.; Zavala-García, F.; Olivares-Sáenz, E.; Niño-Medina, G.; Angélica Ruiz-Torres, N.; Díaz-Barriga, E. Effects of zinc oxide nanoparticles on growth and antioxidant enzymes of Capsicum chinense. Toxicol. Environ. Chem. 2018, 100, 560–572. [Google Scholar] [CrossRef]
- Wang, X.P.; Li, Q.Q.; Pei, Z.M.; Wang, S.C. Effects of zinc oxide nanoparticles on the growth, photosynthetic traits, and antioxidative enzymes in tomato plants. Biol. Plant 2018, 62, 801–808. [Google Scholar] [CrossRef]
- Farghaly, F.A.; Radi, A.A.; Al-Kahtany, F.A.; Hamada, A.M. Impacts of zinc oxide nano and bulk particles on redox-enzymes of the Punica granatum callus. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Wan, J.; Wang, R.; Bai, H.; Wang, Y.; Xu, J. Comparative physiological and metabolomics analysis reveals that single-walled carbon nanohorns and ZnO nanoparticles affect salt tolerance in Sophora alopecuroides. Environ. Sci. Nano 2020, 7, 2968–2981. [Google Scholar] [CrossRef]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. 2017, 482, 419–425. [Google Scholar] [CrossRef]
- Li, Y.L.; Zhan, Y.; Shi, D.Q.; Liu, X.J.; Ge, Q.; Xu, L.H.; Pan, X.L.; Li, W.; Qin, J.; Zhu, Y.Y.; et al. Spatial-temporal analysis of zinc homeostasis reveals the response mechanisms to zinc deficiency in Sorghum bicolor. New Phytol. 2013, 200, 1102–1115. [Google Scholar] [CrossRef]
- Khanm, H.; Vaishnavi, B.; Shankar, A. Rise of nano-fertilizer era: Effect of nano scale zinc oxide particles on the germination, growth and yield of tomato (Solanum lycopersicum). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1861–1871. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 5th ed.; International Potash Institute, Springer Science Business Media: Worblaufen, Switzerland, 1987; pp. 585–596. [Google Scholar]
- Latef, A.A.H.A.; Alhmad, M.F.A.; Abdelfattah, K.E. The possible roles of priming with ZnO nanoparticles in mitigation of salinity stress in lupine (Lupinus termis) plants. J. Plant Growth Regul. 2017, 36, 60–70. [Google Scholar] [CrossRef]
- Zaeem, A.; Drouet, S.; Anjum, S.; Khurshid, R.; Younas, M.; Blondeau, J.P.; Abbasi, B.H. Effects of biogenic zinc oxide nanoparticles on growth and oxidative stress response in flax seedlings vs. In vitro cultures: A comparative analysis. Biomolecules 2020, 10, 918. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, A.P.; Bar-Tal, A.; Silber, A.; Saha, U.K.; Raviv, M.; Lieth, H. Inorganic and Synthetic Organic Components of Soilless Culture and Potting Mixes. Soilless Culture, Theory and Practices; Elsevier Science: Amsterdam, The Netherlands, 2008; pp. 505–543. [Google Scholar]
- Garza-Alonso, C.A.; Guillermo, N.M.; Gutiérrez-Diez, A.; García-López, J.I.; Vázquez-Alvarado, R.E.; López-Jiménez, A.; Olivares-Sáenz, E. Physicochemical characteristics, minerals, phenolic compounds, and antioxidant capacity in fig tree fruits with macronutrient deficiencies. Not. Bot. Horti Agrobot. Cluj Napoca 2020, 48, 1585–1599. [Google Scholar] [CrossRef]
- Alcántar, G.G.; Sandoval, V.M. Handbook Chemical Analysis of Vegetal Tissue; Especial Publication, No.10; Mexican Society of Soil Science: Chapingo, Mexico, 1999. (In Spanish) [Google Scholar]
- Rajput, R.D.; Patil, R.P. The comparative study on spectrophotometric analysis of chlorophyll and carotenoids pigments from non-leguminous fodder crops. Int. J. Innov. Res. Sci. Eng. Technol. 2017, 7, 140–148. [Google Scholar]
- Maclachalam, S.; Zalik, S. Plastid structure, chlorophyll concentration and free amino acid composition of a chlorophyll mutant of barely. Can. J. Bot. 1963, 41, 1053–1062. [Google Scholar] [CrossRef]
- Singh, N.; Ma, L.Q.; Srivastava, M.; Rathinasabapathi, B. Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L and Pteris ensiformis L. Plant Sci. 2006, 170, 274–282. [Google Scholar] [CrossRef]
- Wang, J.; Liu, S.; Li, C.; Wang, T.; Zhang, P.; Chen, K. PnLRR-RLK27, a novel leucine-rich repeats receptor-like protein kinase from the Antarctic moss Pohlia nutans, positively regulates salinity and oxidation-stress tolerance. PLoS ONE 2017, 12, e0172869. [Google Scholar] [CrossRef]
- Kwak, S.S.K.; Soo-Kyung, L.; Moon-Soon, J.; Kyung-Hee, P.; Jang-Ryol, L. Acidic peroxiases from suspension-cultures of sweet potato. Phytochemistry 1995, 39, 981–984. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Elavarthi, S.; Martin, B. Spectrophotometric assays for antioxidant enzymes in plants. Methods Mol. Biol. 2010, 639, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Franklin, N.M.; Rogers, N.J.; Apte, S.C.; Batley, G.E.; Gadd, G.E.; Casey, P.S. Comparative toxicity of nanoparticulate ZnO, bulk ZnO, and ZnCl2 to a freshwater microalga (Pseudokirchneriella subcapitata): The importance of particle solubility. Environ. Sci. Technol. 2007, 41, 8484–8490. [Google Scholar] [CrossRef]
- Reed, R.B.; Ladner, D.A.; Higgins, C.P.; Westerhoff, P.; Ranville, J.F. Solubility of nano-zinc oxide in environmentally and biologically important matrices. Environ. Toxicol. Chem. 2012, 31, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Medina-Velo, I.A.; Barrios, A.C.; Zuverza-Mena, N.; Hernandez-Viezcas, J.A.; Chang, C.H.; Ji, Z.; Gardea-Torresdey, J.L. Comparison of the effects of commercial coated and uncoated ZnO nanomaterials and Zn compounds in kidney bean (Phaseolus vulgaris) plants. J. Hazard. Mater. 2017, 332, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samreen, T.; Shah, H.U.; Ullah, S.; Javid, M. Zinc effect on growth rate, chlorophyll, protein and mineral contents of hydroponically grown mungbeans plant (Vigna radiata). Arab. J. Chem. 2017, 10, 1802–1807. [Google Scholar] [CrossRef] [Green Version]
- Samart, S.; Chutipaijit, S.; Phakamas, N. Evaluating the effect of zinc oxide nanoparticles on the physiological responses of nine non-photoperiod sensitive rice cultivars. Mater. Today 2017, 4, 6430–6435. [Google Scholar] [CrossRef]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Huang, X.; Zhou, Y.; Quan, Q.; Li, Y.; Zhu, X. Response of photosynthesis to different concentrations of heavy metals in Davidia involucrata. PLoS ONE 2020, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Ramel, F.; Birtic, S.; Cuiné, S.; Triantaphylidès, C.; Ravanat, J.L.; Havaux, M. Chemical quenching of singlet oxygen by carotenoids in plants. Plant Physiol. 2012, 158, 1267–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badger, M.R.; Price, G.D. The role of carbonic anhydrase in photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 369–392. [Google Scholar] [CrossRef]
- Mazaheri Tirani, M.; Madadkar-Haghjou, M.; Sulieman, S.; Ismaili, A. Comparative evaluation of zinc oxide effects on tobacco (Nicotiana tabacum L.) grown in different media. J. Agric. Sci. Tech. 2018, 20, 787–802. [Google Scholar]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Masayuki, F.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, C.; Savvides, A.; Christou, A.; Fotopoulos, V. Unravelling chemical priming machinery in plants: The role of reactive oxygen–nitrogen–sulfur species in abiotic stress tolerance enhancement. Curr. Opin. Plant Biol. 2016, 33, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Saeed, M.; Parveen, A.; Javed, M.T.; Zafar, S.; Iqbal, N. Modulation of growth and key physiobiochemical attributes after foliar application of zinc sulphate (ZnSO4) on wheat (Triticum aestivum L.) under cadmium (Cd) stress. Physiol. Mol. Biol. Plants 2020, 26, 1787–1797. [Google Scholar] [CrossRef]
- Nemček, L.; Šebesta, M.; Urík, M.; Bujdoš, M.; Dobročka, E.; Vávra, I. Impact of bulk ZnO, ZnO nanoparticles and dissolved Zn on early growth stages of barley-A pot experiment. Plants 2020, 9, 1365. [Google Scholar] [CrossRef]
- Mosavat, N.; Golkar, P.; Yousefifard, M.; Javed, R. Modulation of callus growth and secondary metabolites in different Thymus species and Zataria multiflora micropropagated under ZnO nanoparticles stress. Biotechnol. Appl. Biochem. 2019, 66, 316–322. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.I.; Zavala-García, F.; Olivares-Sáenz, E.; Lira-Saldívar, R.H.; Díaz Barriga-Castro, E.; Ruiz-Torres, N.A.; Niño-Medina, G. Zinc oxide nanoparticles boosts phenolic compounds and antioxidant activity of Capsicum annuum L. during germination. Agronomy 2018, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Gudkov, S.V.; Grinberg, M.A.; Sukhov, V.; Vodeneev, V. Effect of ionizing radiation on physiological and molecular processes in plants. J. Environ. Radioact. 2019, 202, 8–24. [Google Scholar] [CrossRef]
- Islam, M.; Kim, J.W.; Begum, M.; Sohel, M.; Taher, A.; Lim, Y.S. Physiological and biochemical changes in sugar beet seedlings to confer stress adaptability under drought condition. Plants 2020, 9, 1511. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, N.Á.; Afzal, S.; Singh, T.; Hussain, I. Zinc oxide nanoparticles: A review of their biological synthesis, antimicrobial activity, uptake, translocation and biotransformation in plants. J. Mater. Sci. 2018, 53, 185–201. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Torres, N.; Flores-Naveda, A.; Barriga-Castro, E.D.; Camposeco-Montejo, N.; Ramírez-Barrón, S.; Borrego-Escalante, F.; Niño-Medina, G.; Hernández-Juárez, A.; Garza-Alonso, C.; Rodríguez-Salinas, P.; et al. Zinc Oxide Nanoparticles and Zinc Sulfate Impact Physiological Parameters and Boosts Lipid Peroxidation in Soil Grown Coriander Plants (Coriandrum sativum). Molecules 2021, 26, 1998. https://doi.org/10.3390/molecules26071998
Ruiz-Torres N, Flores-Naveda A, Barriga-Castro ED, Camposeco-Montejo N, Ramírez-Barrón S, Borrego-Escalante F, Niño-Medina G, Hernández-Juárez A, Garza-Alonso C, Rodríguez-Salinas P, et al. Zinc Oxide Nanoparticles and Zinc Sulfate Impact Physiological Parameters and Boosts Lipid Peroxidation in Soil Grown Coriander Plants (Coriandrum sativum). Molecules. 2021; 26(7):1998. https://doi.org/10.3390/molecules26071998
Chicago/Turabian StyleRuiz-Torres, Norma, Antonio Flores-Naveda, Enrique Díaz Barriga-Castro, Neymar Camposeco-Montejo, Sonia Ramírez-Barrón, Fernando Borrego-Escalante, Guillermo Niño-Medina, Agustín Hernández-Juárez, Carlos Garza-Alonso, Pablo Rodríguez-Salinas, and et al. 2021. "Zinc Oxide Nanoparticles and Zinc Sulfate Impact Physiological Parameters and Boosts Lipid Peroxidation in Soil Grown Coriander Plants (Coriandrum sativum)" Molecules 26, no. 7: 1998. https://doi.org/10.3390/molecules26071998
APA StyleRuiz-Torres, N., Flores-Naveda, A., Barriga-Castro, E. D., Camposeco-Montejo, N., Ramírez-Barrón, S., Borrego-Escalante, F., Niño-Medina, G., Hernández-Juárez, A., Garza-Alonso, C., Rodríguez-Salinas, P., & García-López, J. I. (2021). Zinc Oxide Nanoparticles and Zinc Sulfate Impact Physiological Parameters and Boosts Lipid Peroxidation in Soil Grown Coriander Plants (Coriandrum sativum). Molecules, 26(7), 1998. https://doi.org/10.3390/molecules26071998