
The Hallmarks of Flavonoids in Cancer



 ,
,
Abstract
:1. Introduction
2. Flavonoids, Inflammation and Immunity
2.1. The NF-κB Pathway
2.2. The MAPK Pathway
2.3. The Inflammasome Pathway
2.4. The STAT Family Pathway
3. Flavonoids and Redox Metabolism
3.1. Metabolic Alterations in Cancer
3.2. Flavonoids and Oxidative Stress in Cancer
4. Flavonoids and Cell Growth Signaling
4.1. The AKT/mTOR Pathway
4.2. The Ras/ERK Pathway
5. Flavonoids and Autophagy
- -
- -
- Autophagy and sustaining proliferative signaling: the regulation of this process by some molecules such as the accumulation of Adenosine Monophosphate (AMP) and the depletion of Adenosine Triphosphate (ATP) leads to the activation of anti-proliferative pathways, for example, the Liver Kinase B1 (LKB1) and AMP-activated Kinase (AMPK) pathways. These proteins reduce the rate of cell proliferation and also intensify the autophagy process. In some types of cancer with driver mutation in PTEN and LKB1, there is a decrease of autophagy to maintain high levels of proliferation [288,289].
- -
- -
- Autophagy and resistance to cell death: Beclin1 protein interacts with Bcl-2 protein, inhibiting the autophagy process. Thus, when a cancer cell receives a death stimulus such as chemotherapy, the interaction between Beclin1 and Bcl-2 proteins is lost and the cell presents high levels of anti-apoptotic and autophagy activities, turning it resistant to apoptosis [292,293].
- -
- -
- Autophagy and activation of invasion and metastasis: low levels of autophagy are related to EMT with the association of inflammation and macrophages in the tumor. Cancer cells can then detach from the primary tumor region, promoting the metastasis process and producing an invasive tumoral colony. Tumoral colonies can benefit from autophagy since they can avoid the anoikis process, a type of apoptosis due to the loss of contact from the extracellular matrix. Autophagy also plays an important role to overcome starvation and hypoxia until the metastatic cells invade a new region [296,297].
- -
- Autophagy and chemoresistance: some types of cancer have mutations on proliferative pathways such as p53 and MAPK and are known as “autophagy addicted”. The scenario of intense proliferation can be associated with a cellular metabolic disturbance due to the preference for glycolytic metabolism. In this case, autophagy plays a crucial role in tumor progression, protecting the cell against the damage of super proliferation and generating energy substrates through the degradation of organelles and proteins. Thereby, when the autophagy pathway is activated, the cells often present anti-apoptotic activity caused by Beclin1 interaction with Bcl-2, through the BH3 domain, which can lead to chemotherapeutic resistance [292,298].
- -
- Autophagy and senescence: evidence points out that autophagy may correlate with the senescence process. CDKs (Cyclin-dependent-kinase) and cyclins control important pathways in proliferation, such as mTOR and AMPK, that can regulate autophagy. Thus, the deregulation of this system can slow the senescence process [299,300].
6. Flavonoids, Apoptosis, and Cell Cycle
6.1. Apoptotic Pathway
6.2. Cell Cycle Control
6.3. DNA Damage and Repair
6.4. Cell Senescence
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pallauf, K.; Duckstein, N.; Rimbach, G. A literature review of flavonoids and lifespan in model organisms. Proc. Nutr. Soc. 2017, 76, 145–162. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Jiang, Y.; Yang, J.; Zhao, Y.; Tian, M.; Yang, B. Structure, bioactivity, and synthesis of methylated flavonoids. Ann. N. Y. Acad. Sci. 2017, 1398, 120–129. [Google Scholar] [CrossRef]
- Rahaiee, S.; Assadpour, E.; Faridi Esfanjani, A.; Silva, A.S.; Jafari, S.M. Application of nano/microencapsulated phenolic compounds against cancer. Adv. Colloid Interface Sci. 2020, 279, 102153. [Google Scholar] [CrossRef]
- Rodríguez De Luna, S.L.; Ramírez-Garza, R.E.; Serna Saldívar, S.O. Environmentally Friendly Methods for Flavonoid Extraction from Plant Material: Impact of Their Operating Conditions on Yield and Antioxidant Properties. Sci. World J. 2020, 2020, 6792069. [Google Scholar] [CrossRef]
- Sotler, R.; Poljšak, B.; Dahmane, R.; Jukić, T.; Pavan Jukić, D.; Rotim, C.; Trebše, P.; Starc, A. Prooxidant activities of antioxidants and their impact on health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Tan, Y.; Chen, G.; Wang, G.; Sun, J.; Ou, S.; Chen, W.; Bai, W. Metabolism of anthocyanins and consequent effects on the gut microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 982–991. [Google Scholar] [CrossRef]
- Haskell-Ramsay, C.F.; Schmitt, J.; Actis-Goretta, L. The Impact of Epicatechin on Human Cognition: The Role of Cerebral Blood Flow. Nutrients 2018, 10, 986. [Google Scholar] [CrossRef] [Green Version]
- Tahiri, I.; Garro-Aguilar, Y.; Cayssials, V.; Achaintre, D.; Mancini, F.R.; Mahamat-Saleh, Y.; Boutron-Ruault, M.-C.; Kühn, T.; Katzke, V.; Boeing, H.; et al. Urinary flavanone concentrations as biomarkers of dietary flavanone intakes in the European Prospective Investigation into Cancer and Nutrition (EPIC) study. Br. J. Nutr. 2020, 123, 691–698. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Huang, S.; Su, Y. Dietary Flavonols Intake and Risk of Esophageal and Gastric Cancer: A Meta-Analysis of Epidemiological Studies. Nutrients 2016, 8, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, C.C.; Johnson, S.A.; Arjmandi, B.H. Soy and its isoflavones: The truth behind the science in breast cancer. Anticancer Agents Med. Chem. 2013, 13, 1178–1187. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Chahar, M.K.; Sharma, N.; Dobhal, M.P.; Joshi, Y.C. Flavonoids: A versatile source of anticancer drugs. Pharmacogn. Rev. 2011, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Joseph, S.V.; Edirisinghe, I.; Burton-Freeman, B.M. Fruit Polyphenols: A Review of Anti-inflammatory Effects in Humans. Crit. Rev. Food Sci. Nutr. 2016, 56, 419–444. [Google Scholar] [CrossRef]
- Ganai, S.A.; Sheikh, F.A.; Baba, Z.A.; Mir, M.A.; Mantoo, M.A.; Yatoo, M.A. Anticancer activity of the plant flavonoid luteolin against preclinical models of various cancers and insights on different signalling mechanisms modulated. Phytother. Res. 2021. [Google Scholar] [CrossRef]
- Hariton, E.; Locascio, J.J. Randomised controlled trials—The gold standard for effectiveness research: Study design: Randomised controlled trials. BJOG 2018, 125, 1716. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.M.; Ahmann, F.R.; Nagle, R.B.; Hsu, C.-H.; Tangrea, J.A.; Parnes, H.L.; Sokoloff, M.H.; Gretzer, M.B.; Chow, H.-H.S. Randomized, double-blind, placebo-controlled trial of polyphenon E in prostate cancer patients before prostatectomy: Evaluation of potential chemopreventive activities. Cancer Prev. Res. 2012, 5, 290–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crew, K.D.; Brown, P.; Greenlee, H.; Bevers, T.B.; Arun, B.; Hudis, C.; McArthur, H.L.; Chang, J.; Rimawi, M.; Vornik, L.; et al. Phase IB randomized, double-blinded, placebo-controlled, dose escalation study of polyphenon E in women with hormone receptor-negative breast cancer. Cancer Prev. Res. 2012, 5, 1144–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samavat, H.; Wu, A.H.; Ursin, G.; Torkelson, C.J.; Wang, R.; Yu, M.C.; Yee, D.; Kurzer, M.S.; Yuan, J.-M. Green Tea Catechin Extract Supplementation Does Not Influence Circulating Sex Hormones and Insulin-Like Growth Factor Axis Proteins in a Randomized Controlled Trial of Postmenopausal Women at High Risk of Breast Cancer. J. Nutr. 2019, 149, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Garcia, F.A.R.; Cornelison, T.; Nuño, T.; Greenspan, D.L.; Byron, J.W.; Hsu, C.-H.; Alberts, D.S.; Chow, H.-H.S. Results of a phase II randomized, double-blind, placebo-controlled trial of Polyphenon E in women with persistent high-risk HPV infection and low-grade cervical intraepithelial neoplasia. Gynecol. Oncol. 2014, 132, 377–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ávila-Gálvez, M.Á.; García-Villalba, R.; Martínez-Díaz, F.; Ocaña-Castillo, B.; Monedero-Saiz, T.; Torrecillas-Sánchez, A.; Abellán, B.; González-Sarrías, A.; Espín, J.C. Metabolic Profiling of Dietary Polyphenols and Methylxanthines in Normal and Malignant Mammary Tissues from Breast Cancer Patients. Mol. Nutr. Food Res. 2019, 63, 1801239. [Google Scholar] [CrossRef] [PubMed]
- Messing, E.; Gee, J.R.; Saltzstein, D.R.; Kim, K.; DiSant’Agnese, A.; Kolesar, J.; Harris, L.; Faerber, A.; Havighurst, T.; Young, J.M.; et al. A Phase 2 Cancer Chemoprevention Biomarker Trial of Isoflavone G-2535 (Genistein) in Presurgical Bladder Cancer Patients. Cancer Prev. Res. 2012, 5, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Pintova, S.; Dharmupari, S.; Moshier, E.; Zubizarreta, N.; Ang, C.; Holcombe, R.F. Genistein combined with FOLFOX or FOLFOX–Bevacizumab for the treatment of metastatic colorectal cancer: Phase I/II pilot study. Cancer Chemother. Pharmacol. 2019, 84, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Citrin, D.E.; Prasanna, P.G.S.; Walker, A.J.; Freeman, M.L.; Eke, I.; Barcellos-Hoff, M.H.; Arankalayil, M.J.; Cohen, E.P.; Wilkins, R.C.; Ahmed, M.M.; et al. Radiation-Induced Fibrosis: Mechanisms and Opportunities to Mitigate. Report of an NCI Workshop, September 19, 2016. Radiat. Res. 2017, 188, 1–20. [Google Scholar] [CrossRef]
- Helal, A.; Tagliazucchi, D.; Verzelloni, E.; Conte, A. Bioaccessibility of polyphenols and cinnamaldehyde in cinnamon beverages subjected to in vitro gastro-pancreatic digestion. J. Funct. Foods 2014, 7, 506–516. [Google Scholar] [CrossRef]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [Green Version]
- Velderrain-Rodríguez, G.R.; Palafox-Carlos, H.; Wall-Medrano, A.; Ayala-Zavala, J.F.; Chen, C.-Y.O.; Robles-Sánchez, M.; Astiazaran-García, H.; Alvarez-Parrilla, E.; González-Aguilar, G.A. Phenolic compounds: Their journey after intake. Food Funct. 2014, 5, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thilakarathna, S.H.; Rupasinghe, H.P.V. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef] [PubMed]
- Galati, G.; O’Brien, P.J. Potential toxicity of flavonoids and other dietary phenolics: Significance for their chemopreventive and anticancer properties. Free Radic. Biol. Med. 2004, 37, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Birt, D.F.; Jeffery, E. Flavonoids. Adv. Nutr. 2013, 4, 576–577. [Google Scholar] [CrossRef]
- Kashyap, D.; Tuli, H.S.; Yerer, M.B.; Sharma, A.; Sak, K.; Srivastava, S.; Pandey, A.; Garg, V.K.; Sethi, G.; Bishayee, A. Natural product-based nanoformulations for cancer therapy: Opportunities and challenges. Semin. Cancer Biol. 2021, 69, 5–23. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Park, M.J.; Lee, E.K.; Heo, H.S.; Kim, M.S.; Sung, B.; Kim, M.K.; Lee, J.; Kim, N.D.; Anton, S.; Choi, J.S.; et al. The anti-inflammatory effect of kaempferol in aged kidney tissues: The involvement of nuclear factor-κB via nuclear factor-inducing kinase/IκB kinase and mitogen-activated protein kinase pathways. J. Med. Food 2009, 12, 351–358. [Google Scholar] [CrossRef]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Brown, J.; Wang, H.; Hajishengallis, G.N.; Martin, M. TLR-signaling Networks. J. Dent. Res. 2011, 90, 417–427. [Google Scholar] [CrossRef]
- Au, P.Y.B.; Yeh, W.C. Physiological roles and mechanisms of signaling by TRAF2 and TRAF5. Adv. Exp. Med. Biol. 2007, 597, 32–47. [Google Scholar] [PubMed]
- Hoffmann, A. The Ikappa B-NF-kappa B Signaling Module: Temporal Control and Selective Gene Activation. Science 2002, 298, 1241–1245. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Hong, J. Roles of NF-κB in Cancer and Inflammatory Diseases and Their Therapeutic Approaches. Cells 2016, 5, 15. [Google Scholar] [CrossRef]
- Somade, O.T.; Ajayi, B.O.; Adeyi, O.E.; Aina, B.O.; David, B.O.; Sodiya, I.D. Activation of NF-kB mediates up-regulation of cerebellar and hypothalamic pro-inflammatory chemokines (RANTES and MCP-1) and cytokines (TNF-α, IL-1β, IL-6) in acute edible camphor administration. Sci. Afr. 2019, 5, e00114. [Google Scholar] [CrossRef]
- Richmond, A. NF-κB, chemokine gene transcription and tumour growth. Nat. Rev. Immunol. 2002, 2, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, R.; Ng, K.P.; Cicek, M.; Kelleher, C.; Niculaita, R.; Casey, G.; Sizemore, N. Role of IKK and oscillatory NFκB kinetics in MMP-9 gene expression and chemoresistance to 5-fluorouracil in RKO colorectal cancer cells. Mol. Carcinog. 2007, 46, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Guttridge, D.C.; Albanese, C.; Reuther, J.Y.; Pestell, R.G.; Baldwin, A.S. NF-κB Controls Cell Growth and Differentiation through Transcriptional Regulation of Cyclin D1. Mol. Cell. Biol. 1999, 19, 5785–5799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Rosa, F.A.; Pierce, J.W.; Sonenshein, G.E. Differential regulation of the c-myc oncogene promoter by the NF-kappa B rel family of transcription factors. Mol. Cell. Biol. 1994, 14, 1039–1044. [Google Scholar] [CrossRef]
- Heckman, C.A.; Mehew, J.W.; Boxer, L.M. NF-κB activates Bcl-2 expression in t(14;18) lymphoma cells. Oncogene 2002, 21, 3898–3908. [Google Scholar] [CrossRef] [Green Version]
- Khoshnan, A.; Tindell, C.; Laux, I.; Bae, D.; Bennett, B.; Nel, A.E. The NF-κB Cascade Is Important in Bcl-x L Expression and for the Anti-Apoptotic Effects of the CD28 Receptor in Primary Human CD4 + Lymphocytes. J. Immunol. 2000, 165, 1743–1754. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.X.; Xia, Z.; Zhang, N.; Gong, W.; Huang, S. Constitutive NF-$κ$B activity regulates the expression of VEGF and IL-8 and tumor angiogenesis of human glioblastoma. Oncol. Rep. 2010, 23, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, P.; Zhang, W.; Du, Q.; Tang, J.; Wang, H.; Lu, J.; Hu, R. GEN-27, a Newly Synthetic Isoflavonoid, Inhibits the Proliferation of Colon Cancer Cells in Inflammation Microenvironment by Suppressing NF- κ B Pathway. Mediat. Inflamm. 2016, 2016, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadioglu, O.; Nass, J.; Saeed, M.E.M.M.; Schuler, B.; Efferth, T. Kaempferol is an anti-inflammatory compound with activity towards NF-ℵB pathway proteins. Anticancer Res. 2015, 35, 2645–2650. [Google Scholar]
- Shukla, S.; Shankar, E.; Fu, P.; MacLennan, G.T.; Gupta, S. Suppression of NF-kB and NF-KB regulated gene expression by apigenin through IKKα and IKK pathway in TRAMP mice. PLoS ONE 2015, 10, e0138710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidya Priyadarsini, R.; Senthil Murugan, R.; Maitreyi, S.; Ramalingam, K.; Karunagaran, D.; Nagini, S. The flavonoid quercetin induces cell cycle arrest and mitochondria-mediated apoptosis in human cervical cancer (HeLa) cells through p53 induction and NF-κB inhibition. Eur. J. Pharmacol. 2010, 649, 84–91. [Google Scholar] [CrossRef]
- Wu, T.-C.; Chan, S.-T.; Chang, C.-N.; Yu, P.-S.; Chuang, C.-H.; Yeh, S.-L. Quercetin and chrysin inhibit nickel-induced invasion and migration by downregulation of TLR4/NF-κB signaling in A549 cells. Chem. Biol. Interact. 2018, 292, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Siddiqui, M.A.; Dayal, S.; Ayoub, Y.Z.; Malathi, K. Epigallocatechin-3-gallate suppresses proinflammatory cytokines and chemokines induced by Toll-like receptor 9 agonists in prostate cancer cells. J. Inflamm. Res. 2014, 7, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Pratheeshkumar, P.; Son, Y.-O.; Wang, X.; Divya, S.P.; Joseph, B.; Hitron, J.A.; Wang, L.; Kim, D.; Yin, Y.; Roy, R.V.; et al. Cyanidin-3-glucoside inhibits UVB-induced oxidative damage and inflammation by regulating MAP kinase and NF-κB signaling pathways in SKH-1 hairless mice skin. Toxicol. Appl. Pharmacol. 2014, 280, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Im, N.-K.; Jang, W.J.; Jeong, C.-H.; Jeong, G.-S. Delphinidin Suppresses PMA-Induced MMP-9 Expression by Blocking the NF-κB Activation through MAPK Signaling Pathways in MCF-7 Human Breast Carcinoma Cells. J. Med. Food 2014, 17, 855–861. [Google Scholar] [CrossRef]
- Lee, S.-B.; Shin, J.-S.; Han, H.-S.; Lee, H.-H.; Park, J.C.; Lee, K.-T. Kaempferol 7- O—β -D-glucoside isolated from the leaves of Cudrania tricuspidata inhibits LPS-induced expression of pro-inflammatory mediators through inactivation of NF-κB, AP-1, and JAK-STAT in RAW 264.7 macrophages. Chem. Biol. Interact. 2018, 284, 101–111. [Google Scholar] [CrossRef]
- Zhang, M.-J.; Su, H.; Yan, J.-Y.; Li, N.; Song, Z.-Y.; Wang, H.-J.; Huo, L.-G.; Wang, F.; Ji, W.-S.; Qu, X.-J.; et al. Chemopreventive effect of Myricetin, a natural occurring compound, on colonic chronic inflammation and inflammation-driven tumorigenesis in mice. Biomed. Pharmacother. 2018, 97, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Du, Q.; Li, X.; Zheng, X.; Lv, F.; Xi, X.; Huang, G.; Yang, J.; Liu, S. Eriodictyol Inhibits Proliferation, Metastasis and Induces Apoptosis of Glioma Cells via PI3K/Akt/NF-κB Signaling Pathway. Front. Pharmacol. 2020, 11, 114. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Choi, J.G.; Lee, J.H.; Kwon, D.Y. Luteolin isolated from the flowers of Lonicera japonica suppresses inflammatory mediator release by blocking NF-$κ$B and MAPKs activation pathways in HMC-1 cells. Molecules 2010, 15, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, G.; Gurley, E.C.; Zhou, H. Flavonoid apigenin inhibits lipopolysaccharide-induced inflammatory response through multiple mechanisms in macrophages. PLoS ONE 2014, 9, e107072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodduluru, L.N.; Kasala, E.R.; Madhana, R.M.; Barua, C.C.; Hussain, M.I.; Haloi, P.; Borah, P. Naringenin ameliorates inflammation and cell proliferation in benzo(a)pyrene induced pulmonary carcinogenesis by modulating CYP1A1, NFκB and PCNA expression. Int. Immunopharmacol. 2016, 30, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Manigandan, K.; Manimaran, D.; Jayaraj, R.L.; Elangovan, N.; Dhivya, V.; Kaphle, A. Taxifolin curbs NF-κB-mediated Wnt/β-catenin signaling via up-regulating Nrf2 pathway in experimental colon carcinogenesis. Biochimie 2015, 119, 103–112. [Google Scholar] [CrossRef]
- Sun, Z.-J.; Chen, G.; Hu, X.; Zhang, W.; Liu, Y.; Zhu, L.-X.; Zhou, Q.; Zhao, Y.-F. Activation of PI3K/Akt/IKK-α/NF-κB signaling pathway is required for the apoptosis-evasion in human salivary adenoid cystic carcinoma: Its inhibition by quercetin. Apoptosis 2010, 15, 850–863. [Google Scholar] [CrossRef]
- Huang, P.; Han, J.; Hui, L. MAPK signaling in inflammation-associated cancer development. Protein Cell 2010, 1, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Rauch, J.; Kolch, W. Targeting MAPK Signaling in Cancer: Mechanisms of Drug Resistance and Sensitivity. Int. J. Mol. Sci. 2020, 21, 1102. [Google Scholar] [CrossRef] [Green Version]
- Symons, A.; Beinke, S.; Ley, S.C. MAP kinase kinase kinases and innate immunity. Trends Immunol. 2006, 27, 40–48. [Google Scholar] [CrossRef]
- Chadee, D.N.; Kyriakis, J.M. Activation of SAPK/JNKs In Vitro. In MAP Kinase Signaling Protocols; Humana Press: Totowa, NJ, USA, 2010; pp. 59–73. [Google Scholar]
- Angel, P.; Karin, M. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim. Biophys. Acta Rev. Cancer 1991, 1072, 129–157. [Google Scholar] [CrossRef]
- Guinea-Viniegra, J.; Zenz, R.; Scheuch, H.; Hnisz, D.; Holcmann, M.; Bakiri, L.; Schonthaler, H.B.; Sibilia, M.; Wagner, E.F. TNFα shedding and epidermal inflammation are controlled by Jun proteins. Genes Dev. 2009, 23, 2663–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirum-Connolly, K.; Brinckerhoff, C.E. lnterleukin-1 or phorbol induction of the stromelysin promoter requires an element that cooperates with AP-1. Nucleic Acids Res. 1991, 19, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.K.; Bakiri, L.; Hasenfuss, S.C.; Hamacher, R.; Martinez, L.; Wagner, E.F. JUNB/AP-1 controls IFN-γ during inflammatory liver disease. J. Clin. Investig. 2013, 123, 5258–5268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Wenger, L.; Brinckerhoff, C.E.; Misra, R.R.; Cheung, H.S. Basic Calcium Phosphate Crystals Induce Matrix Metalloproteinase-1 through the Ras/Mitogen-activated Protein Kinase/c-Fos/AP-1/Metalloproteinase 1 Pathway. J. Biol. Chem. 2002, 277, 1544–1552. [Google Scholar] [CrossRef] [Green Version]
- Kaminska, B. Molecular Characterization of Inflammation-Induced JNK/c-Jun Signaling Pathway in Connection with Tumorigenesis. In Inflammation and Cancer; Humana Press: Totowa, NJ, USA, 2009; pp. 249–264. [Google Scholar]
- Papachristou, D.; Batistatou, A.; Sykiotis, G..; Varakis, I.; Papavassiliou, A. Activation of the JNK–AP-1 signal transduction pathway is associated with pathogenesis and progression of human osteosarcomas. Bone 2003, 32, 364–371. [Google Scholar] [CrossRef]
- Mendelson, K.G.; Contois, L.-R.; Tevosian, S.G.; Davis, R.J.; Paulson, K.E. Independent regulation of JNK/p38 mitogen-activated protein kinases by metabolic oxidative stress in the liver. Proc. Natl. Acad. Sci. USA 1996, 93, 12908–12913. [Google Scholar] [CrossRef] [Green Version]
- Whitmarsh, A.J.; Davis, R.J. Transcription factor AP-1 regulation by mitogen-activated protein kinase signal transduction pathways. J. Mol. Med. 1996, 74, 589–607. [Google Scholar] [CrossRef]
- Martínez-Limón, A.; Joaquin, M.; Caballero, M.; Posas, F.; de Nadal, E. The p38 Pathway: From Biology to Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 1913. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Goepfert, H.; Boyd, D. Inhibition of the p38 mitogen-activated protein kinase by SB 203580 blocks PMA-induced Mr 92,000 type IV collagenase secretion and in vitro invasion. Cancer Res. 1998, 58, 1135–1139. [Google Scholar] [PubMed]
- Rousseau, S.; Houle, F.; Kotanides, H.; Witte, L.; Waltenberger, J.; Landry, J.; Huot, J. Vascular Endothelial Growth Factor (VEGF)-driven Actin-based Motility Is Mediated by VEGFR2 and Requires Concerted Activation of Stress-activated Protein Kinase 2 (SAPK2/p38) and Geldanamycin-sensitive Phosphorylation of Focal Adhesion Kinase. J. Biol. Chem. 2000, 275, 10661–10672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, N.; Qi, X.; Tsai, S.; Lu, Y.; Basir, Z.; Oshima, K.; Thomas, J.P.; Myers, C.R.; Stoner, G.; Chen, G. p38γ MAPK is required for inflammation-associated colon tumorigenesis. Oncogene 2016, 35, 1039–1048. [Google Scholar] [CrossRef]
- del Reino, P.; Alsina-Beauchamp, D.; Escós, A.; Cerezo-Guisado, M.I.; Risco, A.; Aparicio, N.; Zur, R.; Fernandez-Estévez, M.; Collantes, E.; Montans, J.; et al. Pro-Oncogenic Role of Alternative p38 Mitogen-Activated Protein Kinases p38γ and p38δ, Linking Inflammation and Cancer in Colitis-Associated Colon Cancer. Cancer Res. 2014, 74, 6150–6160. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-Y.; Chang, Y.-M.; Wang, K.-Y.; Chen, P.-N.; Hseu, Y.-C.; Chen, K.-M.; Yeh, K.-T.; Chen, C.-J.; Hsu, L.-S. Naringenin inhibited migration and invasion of glioblastoma cells through multiple mechanisms. Environ. Toxicol. 2019, 34, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Rosa, S.I.G.; Rios-Santos, F.; Balogun, S.O.; de Oliveira Martins, D.T. Vitexin reduces neutrophil migration to inflammatory focus by down-regulating pro-inflammatory mediators via inhibition of p38, ERK1/2 and JNK pathway. Phytomedicine 2016, 23, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, K.V.; Deng, M.; Ting, J.P.-Y. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Jo, E.-K.; Kim, J.K.; Shin, D.-M.; Sasakawa, C. Molecular mechanisms regulating NLRP3 inflammasome activation. Cell. Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Lamkanfi, M. Emerging inflammasome effector mechanisms. Nat. Rev. Immunol. 2011, 11, 213–220. [Google Scholar] [CrossRef]
- Hamarsheh, S.; Zeiser, R. NLRP3 Inflammasome Activation in Cancer: A Double-Edged Sword. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Okamoto, M.; Liu, W.; Luo, Y.; Tanaka, A.; Cai, X.; Norris, D.A.; Dinarello, C.A.; Fujita, M. Constitutively Active Inflammasome in Human Melanoma Cells Mediating Autoinflammation via Caspase-1 Processing and Secretion of Interleukin-1β. J. Biol. Chem. 2010, 285, 6477–6488. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.Y.; Lee, S.-W.; Shin, Y.-H.; Lee, J.-H.; Jahng, J.W.; Park, K. P2X7 receptor and NLRP3 inflammasome activation in head and neck cancer. Oncotarget 2017, 8, 48972–48982. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Du, H.-D.; Tang, D.; Zhang, D.; Zhou, J.; Zhai, C.-W.; Yuan, C.-C.; Hsueh, C.-Y.; Li, S.-J.; Heng, Y.; et al. Correlation Between the NLRP3 Inflammasome and the Prognosis of Patients With LSCC. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef]
- Daley, D.; Mani, V.R.; Mohan, N.; Akkad, N.; Pandian, G.S.D.B.; Savadkar, S.; Lee, K.B.; Torres-Hernandez, A.; Aykut, B.; Diskin, B.; et al. NLRP3 signaling drives macrophage-induced adaptive immune suppression in pancreatic carcinoma. J. Exp. Med. 2017, 214, 1711–1724. [Google Scholar] [CrossRef] [Green Version]
- Poli, G.; Brancorsini, S.; Cochetti, G.; Barillaro, F.; Egidi, M.G.; Mearini, E. Expression of inflammasome-related genes in bladder cancer and their association with cytokeratin 20 messenger RNA. Urol. Oncol. Semin. Orig. Investig. 2015, 33, 505.e1–505.e7. [Google Scholar] [CrossRef]
- Danese, S.; Malesci, A.; Vetrano, S. Colitis-associated cancer: The dark side of inflammatory bowel disease. Gut 2011, 60, 1609–1610. [Google Scholar] [CrossRef]
- Bauer, C.; Duewell, P.; Mayer, C.; Lehr, H.A.; Fitzgerald, K.A.; Dauer, M.; Tschopp, J.; Endres, S.; Latz, E.; Schnurr, M. Colitis induced in mice with dextran sulfate sodium (DSS) is mediated by the NLRP3 inflammasome. Gut 2010, 59, 1192–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Le, T.H.; Du, Q.; Zhao, Z.; Liu, Y.; Zou, J.; Hua, W.; Liu, C.; Zhu, Y. Genistein protects against DSS-induced colitis by inhibiting NLRP3 inflammasome via TGR5-cAMP signaling. Int. Immunopharmacol. 2019, 71, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Li, X.; Zhang, J.; Yuan, Z.; Fu, Y.; Ma, X.; Ren, N. GEN-27 exhibits anti-inflammatory effects by suppressing the activation of NLRP3 inflammasome and NF-κB pathway. Cell Biol. Int. 2019, 43, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.E.; Darnell, J.E. STATs: Transcriptional control and biological impact. Nat. Rev. Mol. Cell Biol. 2002, 3, 651–662. [Google Scholar] [CrossRef]
- Pfitzner, B.S.P.E.; Kliem, B.S.P.S.; Baus, B.S.P.D.; Litterst, B.S.P.M.C. The Role of STATs in Inflammation and Inflammatory Diseases. Curr. Pharm. Des. 2005, 10, 2839–2850. [Google Scholar] [CrossRef]
- Ernst, M.; Najdovska, M.; Grail, D.; Lundgren-May, T.; Buchert, M.; Tye, H.; Matthews, V.B.; Armes, J.; Bhathal, P.S.; Hughes, N.R.; et al. STAT3 and STAT1 mediate IL-11–dependent and inflammation-associated gastric tumorigenesis in gp130 receptor mutant mice. J. Clin. Investig. 2008, 118, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Kortylewski, M.; Xin, H.; Kujawski, M.; Lee, H.; Liu, Y.; Harris, T.; Drake, C.; Pardoll, D.; Yu, H. Regulation of the IL-23 and IL-12 Balance by Stat3 Signaling in the Tumor Microenvironment. Cancer Cell 2009, 15, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Caprioli, F.; Sarra, M.; Caruso, R.; Stolfi, C.; Fina, D.; Sica, G.; MacDonald, T.T.; Pallone, F.; Monteleone, G. Autocrine Regulation of IL-21 Production in Human T Lymphocytes. J. Immunol. 2008, 180, 1800–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Niu, G.; Kortylewski, M.; Burdelya, L.; Shain, K.; Zhang, S.; Bhattacharya, R.; Gabrilovich, D.; Heller, R.; Coppola, D.; et al. Regulation of the innate and adaptive immune responses by Stat-3 signaling in tumor cells. Nat. Med. 2004, 10, 48–54. [Google Scholar] [CrossRef]
- Ogura, H.; Murakami, M.; Okuyama, Y.; Tsuruoka, M.; Kitabayashi, C.; Kanamoto, M.; Nishihara, M.; Iwakura, Y.; Hirano, T. Interleukin-17 Promotes Autoimmunity by Triggering a Positive-Feedback Loop via Interleukin-6 Induction. Immunity 2008, 29, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Kortylewski, M.; Pardoll, D. Crosstalk between cancer and immune cells: Role of STAT3 in the tumour microenvironment. Nat. Rev. Immunol. 2007, 7, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Dai, S.; Dai, J.; Xiao, Y.; Bai, Y.; Chen, B.; Zhou, M. Luteolin decreases invasiveness, deactivates STAT3 signaling, and reverses interleukin-6 induced epithelial–mesenchymal transition and matrix metalloproteinase secretion of pancreatic cancer cells. Onco Targets Ther. 2015, 8, 2989–3001. [Google Scholar] [CrossRef] [Green Version]
- Riahi-Chebbi, I.; Souid, S.; Othman, H.; Haoues, M.; Karoui, H.; Morel, A.; Srairi-Abid, N.; Essafi, M.; Essafi-Benkhadir, K. The Phenolic compound Kaempferol overcomes 5-fluorouracil resistance in human resistant LS174 colon cancer cells. Sci. Rep. 2019, 9, 195. [Google Scholar] [CrossRef]
- Ruela-de-Sousa, R.R.; Fuhler, G.M.; Blom, N.; Ferreira, C.V.; Aoyama, H.; Peppelenbosch, M.P. Cytotoxicity of apigenin on leukemia cell lines: Implications for prevention and therapy. Cell Death Dis. 2010, 1, e19. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.-H.; Chu, J.-H.; Kwan, H.-Y.; Su, T.; Yu, H.; Cheng, C.-Y.; Fu, X.-Q.; Guo, H.; Li, T.; Tse, A.K.-W.; et al. Inhibition of the STAT3 signaling pathway contributes to apigenin-mediated anti-metastatic effect in melanoma. Sci. Rep. 2016, 6, 21731. [Google Scholar] [CrossRef] [Green Version]
- Senggunprai, L.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U. Quercetin and EGCG Exhibit Chemopreventive Effects in Cholangiocarcinoma Cells via Suppression of JAK/STAT Signaling Pathway. Phyther. Res. 2014, 28, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Mace, T.A.; King, S.A.; Ameen, Z.; Elnaggar, O.; Young, G.; Riedl, K.M.; Schwartz, S.J.; Clinton, S.K.; Knobloch, T.J.; Weghorst, C.M.; et al. Bioactive compounds or metabolites from black raspberries modulate T lymphocyte proliferation, myeloid cell differentiation and Jak/STAT signaling. Cancer Immunol. Immunother. 2014, 63, 889–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumamoto, T.; Fujii, M.; Hou, D.-X. Myricetin directly targets JAK1 to inhibit cell transformation. Cancer Lett. 2009, 275, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Wonganan, O.; He, Y.; Shen, X.; Wongkrajang, K.; Suksamrarn, A.; Zhang, G.; Wang, F. 6-Hydroxy-3- O -methyl-kaempferol 6- O -glucopyranoside potentiates the anti-proliferative effect of interferon α/β by promoting activation of the JAK/STAT signaling by inhibiting SOCS3 in hepatocellular carcinoma cells. Toxicol. Appl. Pharmacol. 2017, 336, 31–39. [Google Scholar] [CrossRef]
- Adham, A.N.A.; Abdelfatah, S.; Naqishbandi, A.M.; Mahmoud, N.; Efferth, T. Cytotoxicity of apigenin toward multiple myeloma cell lines and suppression of iNOS and COX-2 expression in STAT1-transfected HEK293 cells. Phytomedicine 2021, 80, 153371. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.-P. Epithelial-mesenchymal transitions in cancer onset and progression. Bull. Acad. Natl. Med. 2009, 193, 1969–1978. [Google Scholar]
- Sadikovic, B.; Al-Romaih, K.; Squire, J.; Zielenska, M. Cause and Consequences of Genetic and Epigenetic Alterations in Human Cancer. Curr. Genom. 2008, 9, 394–408. [Google Scholar] [CrossRef] [Green Version]
- Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative Stress and Oxidative Damage in Carcinogenesis. Toxicol. Pathol. 2010, 38, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. Oxidative stress and cancer: An overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Zheng, L.; Cardaci, S.; Jerby, L.; MacKenzie, E.D.; Sciacovelli, M.; Johnson, T.I.; Gaude, E.; King, A.; Leach, J.D.G.; Edrada-Ebel, R.; et al. Fumarate induces redox-dependent senescence by modifying glutathione metabolism. Nat. Commun. 2015, 6, 6001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrisic, L.; Dudzik, D.; Barbas, C.; Milkovic, L.; Grune, T.; Zarkovic, N. Short overview on metabolomics approach to study pathophysiology of oxidative stress in cancer. Redox Biol. 2018, 14, 47–58. [Google Scholar] [CrossRef]
- Iqbal, M.A.; Gupta, V.; Gopinath, P.; Mazurek, S.; Bamezai, R.N.K. Pyruvate kinase M2 and cancer: An updated assessment. FEBS Lett. 2014, 588, 2685–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiese, E.K.; Hitosugi, T. Tyrosine Kinase Signaling in Cancer Metabolism: PKM2 Paradox in the Warburg Effect. Front. Cell Dev. Biol. 2018, 6. [Google Scholar] [CrossRef]
- Dong, G.; Mao, Q.; Xia, W.; Xu, Y.; Wang, J.; Xu, L.; Jiang, F. PKM2 and cancer: The function of PKM2 beyond glycolysis. Oncol. Lett. 2016, 11, 1980–1986. [Google Scholar] [CrossRef] [Green Version]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of cancer cell metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New roles in redox signaling for an old antioxidant. Front. Pharmacol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sentellas, S.; Morales-Ibanez, O.; Zanuy, M.; Albertí, J.J. GSSG/GSH ratios in cryopreserved rat and human hepatocytes as a biomarker for drug induced oxidative stress. Toxicol. Vitr. 2014, 28, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Griess, B.; Tom, E.; Domann, F.; Teoh-Fitzgerald, M. Extracellular superoxide dismutase and its role in cancer. Free Radic. Biol. Med. 2017, 112, 464–479. [Google Scholar] [CrossRef] [PubMed]
- Parascandolo, A.; Laukkanen, M.O. Carcinogenesis and Reactive Oxygen Species Signaling: Interaction of the NADPH Oxidase NOX1-5 and Superoxide Dismutase 1-3 Signal Transduction Pathways. Antioxid. Redox Signal. 2019, 30, 443–486. [Google Scholar] [CrossRef]
- de la Vega, M.R.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 2, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-X.; Wen, J.-L.; Wang, L.; Wang, X.-P.; Chen, T.-S. Intracellular catalase activity instead of glutathione level dominates the resistance of cells to reactive oxygen species. Cell Stress Chaperones 2019, 24, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kang, N.J.; Rogozin, E.A.; Kim, H.-G.; Cho, Y.Y.; Bode, A.M.; Lee, H.J.; Surh, Y.-J.; Bowden, G.T.; Dong, Z. Myricetin is a novel natural inhibitor of neoplastic cell transformation and MEK1. Carcinogenesis 2007, 28, 1918–1927. [Google Scholar] [CrossRef] [PubMed]
- Bensaad, K.; Tsuruta, A.; Selak, M.A.; Vidal, M.N.C.; Nakano, K.; Bartrons, R.; Gottlieb, E.; Vousden, K.H. TIGAR, a p53-inducible regulator of glycolysis and apoptosis. Cell 2006, 126, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simabuco, F.M.; Morale, M.G.; Pavan, I.C.B.; Morelli, A.P.; Silva, F.R.; Tamura, R.E. p53 and metabolism: From mechanism to therapeutics. Oncotarget 2018, 9, 23780–23823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakya, A.; Cooksey, R.; Cox, J.E.; Wang, V.; McClain, D.A.; Tantin, D. Oct1 loss of function induces a coordinate metabolic shift that opposes tumorigenicity. Nat. Cell Biol. 2009, 11, 320–327. [Google Scholar] [CrossRef]
- Chen, W.; Sun, Z.; Wang, X.-J.; Jiang, T.; Huang, Z.; Fang, D.; Zhang, D.D. Direct interaction between Nrf2 and p21(Cip1/WAF1) upregulates the Nrf2-mediated antioxidant response. Mol. Cell 2009, 34, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Fahl, W.E. Functional Characterization of Transcription Regulators That Interact with the Electrophile Response Element. Biochem. Biophys. Res. Commun. 2001, 289, 212–219. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Koch, K.; Havermann, S.; Büchter, C.; Wätjen, W. Caenorhabditis elegans as model system in pharmacology and toxicology: Effects of flavonoids on redox-sensitive signalling pathways and ageing. Sci. World J. 2014, 2014, 920398. [Google Scholar] [CrossRef] [Green Version]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Brown, J.E.; Khodr, H.; Hider, R.C.; Rice-Evans, C.A. Structural dependence of flavonoid interactions with Cu2+ ions: Implications for their antioxidant properties. Biochem. J. 1998, 330 Pt 3, 1173–1178. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015; ISBN 9780198717478. [Google Scholar]
- Samec, M.; Liskova, A.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Buhrmann, C.; Varghese, E.; Abotaleb, M.; Qaradakhi, T.; Zulli, A.; et al. Flavonoids against the Warburg phenotype—concepts of predictive, preventive and personalised medicine to cut the Gordian knot of cancer cell metabolism. EPMA J. 2020, 11, 377–398. [Google Scholar] [CrossRef] [PubMed]
- Aslan, E.; Guler, C.; Adem, S. In vitro effects of some flavonoids and phenolic acids on human pyruvate kinase isoenzyme M2. J. Enzyme Inhib. Med. Chem. 2016, 31, 314–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, S.; Shi, J.; Yang, P.; Jia, B.; Wu, H.; Zhang, X.; Li, Z. Apigenin Restrains Colon Cancer Cell Proliferation via Targeted Blocking of Pyruvate Kinase M2-Dependent Glycolysis. J. Agric. Food Chem. 2017, 65, 8136–8144. [Google Scholar] [CrossRef]
- Jia, L.; Huang, S.; Yin, X.; Zan, Y.; Guo, Y.; Han, L. Quercetin suppresses the mobility of breast cancer by suppressing glycolysis through Akt-mTOR pathway mediated autophagy induction. Life Sci. 2018, 208, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Dihal, A.A.; van der Woude, H.; Hendriksen, P.J.M.; Charif, H.; Dekker, L.J.; IJsselstijn, L.; de Boer, V.C.J.; Alink, G.M.; Burgers, P.C.; Rietjens, I.M.C.M.; et al. Transcriptome and proteome profiling of colon mucosa from quercetin fed F344 rats point to tumor preventive mechanisms, increased mitochondrial fatty acid degradation and decreased glycolysis. Proteomics 2008, 8, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.; Correia-Branco, A.; Araújo, J.R.; Guimarães, J.T.; Keating, E.; Martel, F. The Chemopreventive Effect of the Dietary Compound Kaempferol on the MCF-7 Human Breast Cancer Cell Line Is Dependent on Inhibition of Glucose Cellular Uptake. Nutr. Cancer 2015, 67, 504–513. [Google Scholar] [CrossRef]
- Elbaz, H.A.; Lee, I.; Antwih, D.A.; Liu, J.; Hüttemann, M.; Zielske, S.P. Epicatechin Stimulates Mitochondrial Activity and Selectively Sensitizes Cancer Cells to Radiation. PLoS ONE 2014, 9, e88322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shay, J.; Elbaz, H.A.; Lee, I.; Zielske, S.P.; Malek, M.H.; Hüttemann, M. Molecular Mechanisms and Therapeutic Effects of (−)-Epicatechin and Other Polyphenols in Cancer, Inflammation, Diabetes, and Neurodegeneration. Oxid. Med. Cell. Longev. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Gao, J.; Liu, Y.; Shi, J.; Gong, Q. Icariside II alleviates oxygen-glucose deprivation and reoxygenation-induced PC12 cell oxidative injury by activating Nrf2/SIRT3 signaling pathway. Biomed. Pharmacother. 2018, 103, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Van Der Merwe, J.D.; De Beer, D.; Joubert, E.; Gelderblom, W.C.A. Short-term and sub-chronic dietary exposure to aspalathin-enriched green rooibos (Aspalathus linearis) extract affects rat liver function and antioxidant status. Molecules 2015, 20, 22674–22690. [Google Scholar] [CrossRef] [Green Version]
- Mimura, Y.; Yasujima, T.; Ohta, K.; Inoue, K.; Yuasa, H. Functional identification of organic cation transporter 1 as an atenolol transporter sensitive to flavonoids. Biochem. Biophys. Rep. 2015, 2, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wunpathe, C.; Potue, P.; Maneesai, P.; Bunbupha, S.; Prachaney, P.; Kukongviriyapan, U.; Kukongviriyapan, V.; Pakdeechote, P. Hesperidin Suppresses Renin-Angiotensin System Mediated NOX2 Over-Expression and Sympathoexcitation in 2K-1C Hypertensive Rats. Am. J. Chin. Med. 2018, 46, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhuang, M.; Zhong, C.; Peng, J.; Wang, X.; Li, J.; Chen, Z.; Huang, Y. Baicalein reverses hypoxia-induced 5-FU resistance in gastric cancer AGS cells through suppression of glycolysis and the PTEN/Akt/HIF-1α signaling pathway. Oncol. Rep. 2015, 33, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Q.; Wu, R.; Xiao, X.; Yang, C.; Yang, Y.; Wang, C.; Lin, L.; Kong, A.-N. The dietary flavone luteolin epigenetically activates the Nrf2 pathway and blocks cell transformation in human colorectal cancer HCT116 cells. J. Cell. Biochem. 2018, 119, 9573–9582. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cai, X.; Yang, J.; Sun, X.; Hu, C.; Yan, Z.; Xu, X.; Lu, W.; Wang, X.; Cao, P. Chemoprevention of dietary digitoflavone on colitis-associated colon tumorigenesis through inducing Nrf2 signaling pathway and inhibition of inflammation. Mol. Cancer 2014, 13, 48. [Google Scholar] [CrossRef] [Green Version]
- Paredes-Gonzalez, X.; Fuentes, F.; Jeffery, S.; Saw, C.L.-L.; Shu, L.; Su, Z.-Y.; Kong, A.-N.T. Induction of NRF2-mediated gene expression by dietary phytochemical flavones apigenin and luteolin. Biopharm. Drug Dispos. 2015, 36, 440–451. [Google Scholar] [CrossRef]
- Smirnova, N.A.; Kaidery, N.A.; Hushpulian, D.M.; Rakhman, I.I.; Poloznikov, A.A.; Tishkov, V.I.; Karuppagounder, S.S.; Gaisina, I.N.; Pekcec, A.; Van Leyen, K.; et al. Bioactive Flavonoids and Catechols as Hif1 and Nrf2 Protein Stabilizers—Implications for Parkinson’s Disease. Aging Dis. 2016, 7, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Yamamoto, M. The KEAP1–NRF2 System in Cancer. Front. Oncol. 2017, 7. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, H.; Chen, F.; Fu, J.; Xu, Y.; Hou, Y.; Kou, H.H.; Zhai, C.; Nelson, M.B.; Zhang, Q.; et al. An overview of chemical inhibitors of the Nrf2-ARE signaling pathway and their potential applications in cancer therapy. Free Radic. Biol. Med. 2016, 99, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Thiel, G.; Rössler, O.G. Resveratrol regulates gene transcription via activation of stimulus-responsive transcription factors. Pharmacol. Res. 2017, 117, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, M.A. Combination of siRNA-directed gene silencing with epigallocatechin-3-gallate (EGCG) reverses drug resistance in human breast cancer cells. J. Chem. Biol. 2016, 9, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilic, U.; Sahin, K.; Tuzcu, M.; Basak, N.; Orhan, C.; Elibol-Can, B.; Kilic, E.; Sahin, F.; Kucuk, O. Enhancement of Cisplatin Sensitivity in Human Cervical Cancer: Epigallocatechin-3-Gallate. Front. Nutr. 2015, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Wang, H.; Fan, L.; Wu, X.; Xin, A.; Ren, H.; Wang, X.J. Luteolin inhibits Nrf2 leading to negative regulation of the Nrf2/ARE pathway and sensitization of human lung carcinoma A549 cells to therapeutic drugs. Free Radic. Biol. Med. 2011, 50, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Tanigawa, S.; Fujii, M.; Hou, D. Action of Nrf2 and Keap1 in ARE-mediated NQO1 expression by quercetin. Free Radic. Biol. Med. 2007, 42, 1690–1703. [Google Scholar] [CrossRef]
- Kerimi, A.; Williamson, G. Differential Impact of Flavonoids on Redox Modulation, Bioenergetics, and Cell Signaling in Normal and Tumor Cells: A Comprehensive Review. Antioxid. Redox Signal. 2018, 29, 1633–1659. [Google Scholar] [CrossRef]
- Valdameri, G.; Trombetta-Lima, M.; Worfel, P.R.; Pires, A.R.A.; Martinez, G.R.; Noleto, G.R.; Cadena, S.M.S.C.; Sogayar, M.C.; Winnischofer, S.M.B.; Rocha, M.E.M. Involvement of catalase in the apoptotic mechanism induced by apigenin in HepG2 human hepatoma cells. Chem. Biol. Interact. 2011, 193, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Gao, A.-M.; Ke, Z.-P.; Wang, J.-N.; Yang, J.-Y.; Chen, S.-Y.; Chen, H. Apigenin sensitizes doxorubicin-resistant hepatocellular carcinoma BEL-7402/ADM cells to doxorubicin via inhibiting PI3K/Akt/Nrf2 pathway. Carcinogenesis 2013, 34, 1806–1814. [Google Scholar] [CrossRef]
- Gao, A.-M.; Ke, Z.-P.; Shi, F.; Sun, G.-C.; Chen, H. Chrysin enhances sensitivity of BEL-7402/ADM cells to doxorubicin by suppressing PI3K/Akt/Nrf2 and ERK/Nrf2 pathway. Chem. Biol. Interact. 2013, 206, 100–108. [Google Scholar] [CrossRef]
- Qin, S.; Chen, J.; Tanigawa, S.; Hou, D.-X. Microarray and pathway analysis highlight Nrf2/ARE-mediated expression profiling by polyphenolic myricetin. Mol. Nutr. Food Res. 2013, 57, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Granado-Serrano, A.B.; Martín, M.A.; Haegeman, G.; Goya, L.; Bravo, L.; Ramos, S. Epicatechin induces NF-κB, activator protein-1 (AP-1) and nuclear transcription factor erythroid 2p45-related factor-2 (Nrf2) via phosphatidylinositol-3-kinase/protein kinase B (PI3K/AKT) and extracellular regulated kinase (ERK) signalling in HepG2 cells. Br. J. Nutr. 2010, 103, 168–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharjee, S.; Dashwood, R.H. Epigenetic Regulation of NRF2/KEAP1 by Phytochemicals. Antioxidants 2020, 9, 865. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, C.; Liu, B.; Jin, X.; Li, P.; Zheng, X.; Zhao, T.; Li, F.; Li, Q. Genistein mediates the selective radiosensitizing effect in NSCLC A549 cells via inhibiting methylation of the keap1 gene promoter region. Oncotarget 2016, 7, 27267–27279. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Wang, K.S.; Qi, M.; Zhou, Y.J.; Zeng, G.Y.; Tao, J.; Da Zhou, J.; Zhang, J.L.; Chen, X.; Peng, C. Vitexin compound 1, a novel extraction from a Chinese herb, suppresses melanoma cell growth through DNA damage by increasing ROS levels. J. Exp. Clin. Cancer Res. 2018, 37, 269. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Lee, Y.J. Quercetin suppresses hypoxia-induced accumulation of hypoxia-inducible factor-1α (HIF-1α) through inhibiting protein synthesis. J. Cell. Biochem. 2008, 105, 546–553. [Google Scholar] [CrossRef]
- Samec, M.; Liskova, A.; Koklesova, L.; Mersakova, S.; Strnadel, J.; Kajo, K.; Pec, M.; Zhai, K.; Smejkal, K.; Mirzaei, S.; et al. Flavonoids Targeting HIF-1: Implications on Cancer Metabolism. Cancers 2021, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Lin, H.; Wang, M.; Zhang, S.; Wu, X.; Lu, L.; Ji, L.; Yu, L. Quercetin greatly improved therapeutic index of doxorubicin against 4T1 breast cancer by its opposing effects on HIF-1α in tumor and normal cells. Cancer Chemother. Pharmacol. 2010, 65, 277–287. [Google Scholar] [CrossRef]
- Fu, J.-D.; Yao, J.-J.; Wang, H.; Cui, W.-G.; Leng, J.; Ding, L.-Y.; Fan, K.-Y. Effects of EGCG on proliferation and apoptosis of gastric cancer SGC7901 cells via down-regulation of HIF-1α and VEGF under a hypoxic state. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 155–161. [Google Scholar] [CrossRef]
- Huang, H.C.; Tao, M.H.; Hung, T.M.; Chen, J.C.; Lin, Z.J.; Huang, C. (-)-Epigallocatechin-3-gallate inhibits entry of hepatitis B virus into hepatocytes. Antivir. Res. 2014, 111, 100–111. [Google Scholar] [CrossRef]
- Fang, B.; Chen, X.; Wu, M.; Kong, H.; Chu, G.; Zhou, Z.; Zhang, C.; Chen, B. Luteolin inhibits angiogenesis of the M2-like TAMs via the downregulation of hypoxia inducible factor-1α and the STAT3 signalling pathway under hypoxia. Mol. Med. Rep. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonis, I.; Lakka, A.; Tsakalof, A.; Simos, G. The dietary flavonoid kaempferol effectively inhibits HIF-1 activity and hepatoma cancer cell viability under hypoxic conditions. Biochem. Biophys. Res. Commun. 2010, 398, 74–78. [Google Scholar] [CrossRef]
- Melstrom, L.G.; Salabat, M.R.; Ding, X.-Z.; Strouch, M.J.; Grippo, P.J.; Mirzoeva, S.; Pelling, J.C.; Bentrem, D.J. Apigenin Down-Regulates the Hypoxia Response Genes: HIF-1α, GLUT-1, and VEGF in Human Pancreatic Cancer Cells. J. Surg. Res. 2011, 167, 173–181. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Dai, W.; Zhang, Q.; Feng, J.; Wu, L.; Liu, T.; Yu, Q.; Xu, S.; Wang, W.; et al. Genistein suppresses aerobic glycolysis and induces hepatocellular carcinoma cell death. Br. J. Cancer 2017, 117, 1518–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanamala, J.; Leonardi, T.; Patil, B.S.; Taddeo, S.S.; Murphy, M.E.; Pike, L.M.; Chapkin, R.S.; Lupton, J.R.; Turner, N.D. Suppression of colon carcinogenesis by bioactive compounds in grapefruit. Carcinogenesis 2006, 27, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, T.; Vanamala, J.; Taddeo, S.S.; Davidson, L.A.; Murphy, M.E.; Patil, B.S.; Wang, N.; Carroll, R.J.; Chapkin, R.S.; Lupton, J.R.; et al. Apigenin and naringenin suppress colon carcinogenesis through the aberrant crypt stage in azoxymethane-treated rats. Exp. Biol. Med. 2010, 235, 710–717. [Google Scholar] [CrossRef] [Green Version]
- Pandurangan, A.K.; Kumar, S.A.S.; Dharmalingam, P.; Ganapasam, S. Luteolin, a bioflavonoid inhibits azoxymethane-induced colon carcinogenesis: Involvement of iNOS and COX-2. Pharmacogn. Mag. 2014, 10, S306–S310. [Google Scholar] [CrossRef] [Green Version]
- Froyen, E.B.; Steinberg, F.M. Soy isoflavones increase quinone reductase in hepa-1c1c7 cells via estrogen receptor beta and nuclear factor erythroid 2-related factor 2 binding to the antioxidant response element. J. Nutr. Biochem. 2011, 22, 843–848. [Google Scholar] [CrossRef]
- Bumke-Vogt, C.; Osterhoff, M.A.; Borchert, A.; Guzman-Perez, V.; Sarem, Z.; Birkenfeld, A.L.; Bähr, V.; Pfeiffer, A.F.H. The Flavones Apigenin and Luteolin Induce FOXO1 Translocation but Inhibit Gluconeogenic and Lipogenic Gene Expression in Human Cells. PLoS ONE 2014, 9, e104321. [Google Scholar] [CrossRef]
- Silva, L.G.S.; Morelli, A.P.; Pavan, I.C.B.; Tavares, M.R.; Pestana, N.F.; Rostagno, M.A.; Simabuco, F.M.; Bezerra, R.M.N. Protective effects of beet (Beta vulgaris) leaves extract against oxidative stress in endothelial cells in vitro. Phyther. Res. 2020, 34, 1385–1396. [Google Scholar] [CrossRef]
- Sharma, V.; Joseph, C.; Ghosh, S.; Agarwal, A.; Mishra, M.K.; Sen, E. Kaempferol induces apoptosis in glioblastoma cells through oxidative stress. Mol. Cancer Ther. 2007, 6, 2544–2553. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-B.; Kim, J.-H.; Lee, H.; Pak, J.-N.; Shim, B.S.; Kim, S.-H. Reactive Oxygen Species and p53 Mediated Activation of p38 and Caspases is Critically Involved in Kaempferol Induced Apoptosis in Colorectal Cancer Cells. J. Agric. Food Chem. 2018, 66, 9960–9967. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Park, J.H.Y. Kaempferol Induces Cell Cycle Arrest in HT-29 Human Colon Cancer Cells. J. Cancer Prev. 2013, 18, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Bao, J.; Wei, Y.; Chen, Y.; Mao, X.; Li, J.; Yang, Z.; Xue, Y. Kaempferol inhibits gastric cancer tumor growth: An in vitro and in vivo study. Oncol. Rep. 2015, 33, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-W.; Lee, W.; Shin, S.; Kim, G.-Y.; Choi, B.; Choi, Y. Anthocyanins Downregulate Lipopolysaccharide-Induced Inflammatory Responses in BV2 Microglial Cells by Suppressing the NF-κB and Akt/MAPKs Signaling Pathways. Int. J. Mol. Sci. 2013, 14, 1502–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haseeb, A.; Chen, D.; Haqqi, T.M. Delphinidin inhibits IL-1 -induced activation of NF- B by modulating the phosphorylation of IRAK-1Ser376 in human articular chondrocytes. Rheumatology 2013, 52, 998–1008. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhang, B.; Shan, S.; Zhao, X. Neuroprotective effects of vitexin against isoflurane-induced neurotoxicity by targeting the TRPV1 and NR2B signaling pathways. Mol. Med. Rep. 2016, 14, 5607–5613. [Google Scholar] [CrossRef]
- Wing-Cheung Leung, H.; Kuo, C.-L.; Yang, W.-H.; Lin, C.-H.; Lee, H.-Z. Antioxidant enzymes activity involvement in luteolin-induced human lung squamous carcinoma CH27 cell apoptosis. Eur. J. Pharmacol. 2006, 534, 12–18. [Google Scholar] [CrossRef]
- Chen, K.-C.; Hsu, W.-H.; Ho, J.-Y.; Lin, C.-W.; Chu, C.-Y.; Kandaswami, C.C.; Lee, M.-T.; Cheng, C.-H. Flavonoids Luteolin and Quercetin Inhibit RPS19 and contributes to metastasis of cancer cells through c-Myc reduction. J. Food Drug Anal. 2018, 26, 1180–1191. [Google Scholar] [CrossRef] [Green Version]
- DeBerardinis, R.J.; Lum, J.J.; Hatzivassiliou, G.; Thompson, C.B. The Biology of Cancer: Metabolic Reprogramming Fuels Cell Growth and Proliferation. Cell Metab. 2008, 7, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.; Dai, Q.; Su, X.; Fu, J.; Feng, X.; Peng, J. Role of PI3K/AKT pathway in cancer: The framework of malignant behavior. Mol. Biol. Rep. 2020, 47, 4587–4629. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, Q.; Li, H.; Yang, B.; Wang, M. Vitexin suppresses renal cell carcinoma by regulating mTOR pathways. Transl. Androl. Urol. 2020, 9, 1700–1711. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jiang, Q.; Liu, H.; Luo, S. Vitexin induces apoptosis through mitochondrial pathway and PI3K/Akt/mTOR signaling in human non-small cell lung cancer A549 cells. Biol. Res. 2019, 52, 7. [Google Scholar] [CrossRef]
- Wang, R.; Zhu, X.; Wang, Q.; Li, X.; Wang, E.; Zhao, Q.; Wang, Q.; Cao, H. The anti-tumor effect of taxifolin on lung cancer via suppressing stemness and epithelial-mesenchymal transition in vitro and oncogenesis in nude mice. Ann. Transl. Med. 2020, 8, 590. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, R.; Ni, H. Eriodictyol exerts potent anticancer activity against A549 human lung cancer cell line by inducing mitochondrial-mediated apoptosis, G2/M cell cycle arrest and inhibition of m-TOR/PI3K/Akt signalling pathway. Arch. Med. Sci. 2020, 16, 446–452. [Google Scholar] [CrossRef]
- Kaushik, S.; Shyam, H.; Sharma, R.; Balapure, A.K. Dietary isoflavone daidzein synergizes centchroman action via induction of apoptosis and inhibition of PI3K/Akt pathway in MCF-7/MDA MB-231 human breast cancer cells. Phytomedicine 2018, 40, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.K.L.; Siu, M.K.Y.; Jiang, Y.-X.; Wang, J.-J.; Leung, T.H.Y.; Ngan, H.Y.S. Estrogen receptor modulators genistein, daidzein and ERB-041 inhibit cell migration, invasion, proliferation and sphere formation via modulation of FAK and PI3K/AKT signaling in ovarian cancer. Cancer Cell Int. 2018, 18, 65. [Google Scholar] [CrossRef] [Green Version]
- Pal, H.C.; Sharma, S.; Strickland, L.R.; Agarwal, J.; Athar, M.; Elmets, C.A.; Afaq, F. Delphinidin reduces cell proliferation and induces apoptosis of non-small-cell lung cancer cells by targeting EGFR/VEGFR2 signaling pathways. PLoS ONE 2013, 8, e77270. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Liu, X.; Li, H.; Yan, Y.; Hong, X.; Lin, Z. Kaempferol inhibits proliferation, migration, and invasion of liver cancer HepG2 cells by down-regulation of microRNA-21. Int. J. Immunopathol. Pharmacol. 2018, 32. [Google Scholar] [CrossRef] [Green Version]
- Kashafi, E.; Moradzadeh, M.; Mohamadkhani, A.; Erfanian, S. Kaempferol increases apoptosis in human cervical cancer HeLa cells via PI3K/AKT and telomerase pathways. Biomed. Pharmacother. 2017, 89, 573–577. [Google Scholar] [CrossRef]
- Liu, S.; Xu, Z.-L.; Sun, L.; Liu, Y.; Li, C.-C.; Li, H.-M.; Zhang, W.; Li, C.-J.; Qin, W. (-)-Epigallocatechin-3-gallate induces apoptosis in human pancreatic cancer cells via PTEN. Mol. Med. Rep. 2016, 14, 599–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oi, N.; Chen, H.; Ok Kim, M.; Lubet, R.A.; Bode, A.M.; Dong, Z. Taxifolin Suppresses UV-Induced Skin Carcinogenesis by Targeting EGFR and PI3K. Cancer Prev. Res. 2012, 5, 1103–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Bjornsti, M.-A.; Houghton, P.J. Rapamycins: Mechanism of action and cellular resistance. Cancer Biol. Ther. 2003, 2, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Amaral, C.L.; Freitas, L.B.; Tamura, R.E.; Tavares, M.R.; Pavan, I.C.B.; Bajgelman, M.C.; Simabuco, F.M. S6Ks isoforms contribute to viability, migration, docetaxel resistance and tumor formation of prostate cancer cells. BMC Cancer 2016, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hirashima, K.; Baba, Y.; Watanabe, M.; Karashima, R.; Sato, N.; Imamura, Y.; Hiyoshi, Y.; Nagai, Y.; Hayashi, N.; Iyama, K.; et al. Phosphorylated mTOR expression is associated with poor prognosis for patients with esophageal squamous cell carcinoma. Ann. Surg. Oncol. 2010, 17, 2486–2493. [Google Scholar] [CrossRef]
- Sun, S.-Y.; Rosenberg, L.M.; Wang, X.; Zhou, Z.; Yue, P.; Fu, H.; Khuri, F.R. Activation of Akt and eIF4E Survival Pathways by Rapamycin-Mediated Mammalian Target of Rapamycin Inhibition. Cancer Res. 2005, 65, 7052–7058. [Google Scholar] [CrossRef] [Green Version]
- Magnuson, B.; Ekim, B.; Fingar, D.C. Regulation and function of ribosomal protein S6 kinase (S6K) within mTOR signalling networks. Biochem. J. 2012, 441, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Tavares, M.R.; Pavan, I.C.B.; Amaral, C.L.; Meneguello, L.; Luchessi, A.D.; Simabuco, F.M. The S6K protein family in health and disease. Life Sci. 2015, 131, 1–10. [Google Scholar] [CrossRef]
- Rivera Rivera, A.; Castillo-Pichardo, L.; Gerena, Y.; Dharmawardhane, S. Anti-Breast Cancer Potential of Quercetin via the Akt/AMPK/Mammalian Target of Rapamycin (mTOR) Signaling Cascade. PLoS ONE 2016, 11, e0157251. [Google Scholar] [CrossRef]
- Tong, X.; Smith, K.A.; Pelling, J.C. Apigenin, a chemopreventive bioflavonoid, induces AMP-activated protein kinase activation in human keratinocytes. Mol. Carcinog. 2012, 51, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Cao, X.; Li, N.; Zhang, Y.; Lan, L.; Zhou, Y.; Pan, X.; Shen, L.; Yin, Z.; Luo, L. Luteolin is effective in the non-small cell lung cancer model with L858R/T790M EGF receptor mutation and erlotinib resistance. Br. J. Pharmacol. 2014, 171, 2842–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caron, A.; Richard, D.; Laplante, M. The Roles of mTOR Complexes in Lipid Metabolism. Annu. Rev. Nutr. 2015, 35, 321–348. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.; Yang, C.; Bazer, F.W.; Song, G. Luteolin Inhibits Proliferation and Induces Apoptosis of Human Placental Choriocarcinoma Cells by Blocking the PI3K/AKT Pathway and Regulating Sterol Regulatory Element Binding Protein Activity. Biol. Reprod. 2016, 95, 82. [Google Scholar] [CrossRef] [PubMed]
- Damiano, F.; Giannotti, L.; Gnoni, G.V.; Siculella, L.; Gnoni, A. Quercetin inhibition of SREBPs and ChREBP expression results in reduced cholesterol and fatty acid synthesis in C6 glioma cells. Int. J. Biochem. Cell Biol. 2019, 117. [Google Scholar] [CrossRef]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.-H. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Revathidevi, S.; Munirajan, A.K. Akt in cancer: Mediator and more. Semin. Cancer Biol. 2019, 59, 80–91. [Google Scholar] [CrossRef]
- Wu, H.; Pan, L.; Gao, C.; Xu, H.; Li, Y.; Zhang, L.; Ma, L.; Meng, L.; Sun, X.; Qin, H. Quercetin Inhibits the Proliferation of Glycolysis-Addicted HCC Cells by Reducing Hexokinase 2 and Akt-mTOR Pathway. Molecules 2019, 24, 1993. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Saini, K.S.; Kumar, A.; Kumar, S.; Ramakrishna, E.; Maurya, R.; Konwar, R.; Chattopadhyay, N. Quercetin-6-C-β-D-glucopyranoside, natural analog of quercetin exhibits anti-prostate cancer activity by inhibiting Akt-mTOR pathway via aryl hydrocarbon receptor. Biochimie 2015, 119, 68–79. [Google Scholar] [CrossRef]
- Cao, L.; Yang, Y.; Ye, Z.; Lin, B.; Zeng, J.; Li, C.; Liang, T.; Zhou, K.; Li, J. Quercetin-3-methyl ether suppresses human breast cancer stem cell formation by inhibiting the Notch1 and PI3K/Akt signaling pathways. Int. J. Mol. Med. 2018, 42, 1625–1636. [Google Scholar] [CrossRef] [Green Version]
- Marfe, G.; Tafani, M.; Indelicato, M.; Sinibaldi-Salimei, P.; Reali, V.; Pucci, B.; Fini, M.; Russo, M.A. Kaempferol induces apoptosis in two different cell lines via Akt inactivation, Bax and SIRT3 activation, and mitochondrial dysfunction. J. Cell. Biochem. 2009, 106, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Su, M.; Qiu, W.; Zhang, M.; Guo, Z.; Su, B.; Liu, J.; Li, X.; Zhou, L. Kaempferol promotes apoptosis in human bladder cancer cells by inducing the tumor suppressor, PTEN. Int. J. Mol. Sci. 2013, 14, 21215–21226. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.-W.; Chen, P.-N.; Wu, H.-C.; Wu, S.-W.; Tsai, P.-Y.; Hsieh, Y.-S.; Chang, H.-R. Kaempferol Inhibits the Invasion and Migration of Renal Cancer Cells through the Downregulation of AKT and FAK Pathways. Int. J. Med. Sci. 2017, 14, 984–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.T.T.; Tran, E.; Ong, C.K.; Lee, S.K.; Do, P.T.; Huynh, T.T.; Nguyen, T.H.; Lee, J.J.; Tan, Y.; Ong, C.S.; et al. Kaempferol-induced growth inhibition and apoptosis in A549 lung cancer cells is mediated by activation of MEK-MAPK. J. Cell. Physiol. 2003, 197, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: A novel therapeutic strategy. Semin. Cancer Biol. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yao, Y.; Shi, Z.; Everaert, N.; Ren, G. Synergistic Effect of Bioactive Anticarcinogens from Soybean on Anti-Proliferative Activity in MDA-MB-231 and MCF-7 Human Breast Cancer Cells In Vitro. Molecules 2018, 23, 1557. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Chen, A.Y.; Rojanasakul, Y.; Ye, X.; Rankin, G.O.; Chen, Y.C. Dietary compounds galangin and myricetin suppress ovarian cancer cell angiogenesis. J. Funct. Foods 2015, 15, 464–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.-J.; Huang, K.-M.; Gui, H.; Wang, J.-J.; Lu, J.-T.; Dai, L.-J.; Zhang, L.; Wang, G. Pluronic-based micelle encapsulation potentiates myricetin-induced cytotoxicity in human glioblastoma cells. Int. J. Nanomed. 2016, 11, 4991–5002. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-J.; Chen, W.-K.; Wang, C.-J.; Lin, W.-L.; Tseng, T.-H. Apigenin inhibits HGF-promoted invasive growth and metastasis involving blocking PI3K/Akt pathway and beta 4 integrin function in MDA-MB-231 breast cancer cells. Toxicol. Appl. Pharmacol. 2008, 226, 178–191. [Google Scholar] [CrossRef]
- Zhao, G.; Han, X.; Cheng, W.; Ni, J.; Zhang, Y.; Lin, J.; Song, Z. Apigenin inhibits proliferation and invasion, and induces apoptosis and cell cycle arrest in human melanoma cells. Oncol. Rep. 2017, 37, 2277–2285. [Google Scholar] [CrossRef] [Green Version]
- Iida, K.; Naiki, T.; Naiki-Ito, A.; Suzuki, S.; Kato, H.; Nozaki, S.; Nagai, T.; Etani, T.; Nagayasu, Y.; Ando, R.; et al. Luteolin suppresses bladder cancer growth via regulation of mechanistic target of rapamycin pathway. Cancer Sci. 2020, 111, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gu, N.; Xue, C.; Li, B.-R. Plant flavonoid taxifolin inhibits the growth, migration and invasion of human osteosarcoma cells. Mol. Med. Rep. 2018, 17, 3239–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, M.W.; Bose, P.; Siddique, M.U.M.; Sunita, P.; Lapenna, A.; Pattanayak, S.P. Taxifolin binds with LXR (α & β) to attenuate DMBA-induced mammary carcinogenesis through mTOR/Maf-1/PTEN pathway. Biomed. Pharmacother. 2018, 105, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Harmon, A.W.; Patel, Y.M. Naringenin inhibits glucose uptake in MCF-7 breast cancer cells: A mechanism for impaired cellular proliferation. Breast Cancer Res. Treat. 2004, 85, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhong, X.; Zhang, X.; Shang, D.; Zhou, Y.I.; Zhang, C. Enhanced anticancer effect of ABT-737 in combination with naringenin on gastric cancer cells. Exp. Ther. Med. 2016, 11, 669–673. [Google Scholar] [CrossRef]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Yasuda, Y.; Kubota, M.; Adachi, S.; Tsurumi, H.; Hara, Y.; Moriwaki, H. (-)-Epigallocatechin gallate inhibits growth and activation of the VEGF/VEGFR axis in human colorectal cancer cells. Chem. Biol. Interact. 2010, 185, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Van Aller, G.S.; Carson, J.D.; Tang, W.; Peng, H.; Zhao, L.; Copeland, R.A.; Tummino, P.J.; Luo, L. Epigallocatechin gallate (EGCG), a major component of green tea, is a dual phosphoinositide-3-kinase/mTOR inhibitor. Biochem. Biophys. Res. Commun. 2011, 406, 194–199. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Z.-S.; Zhang, X.-H.; Yang, S.-N.; Liu, D.; Diao, C.-R.; Wang, H.; Zheng, F.-P. Cyanidin inhibits EMT induced by oxaliplatin via targeting the PDK1-PI3K/Akt signaling pathway. Food Funct. 2019, 10, 592–601. [Google Scholar] [CrossRef]
- Wang, L.; Li, H.; Yang, S.; Ma, W.; Liu, M.; Guo, S.; Zhan, J.; Zhang, H.; Tsang, S.Y.; Zhang, Z.; et al. Cyanidin-3-o-glucoside directly binds to ERα36 and inhibits EGFR-positive triple-negative breast cancer. Oncotarget 2016, 7, 68864–68882. [Google Scholar] [CrossRef] [Green Version]
- Ya, F.; Li, Q.; Wang, D.; Xie, S.; Song, F.; Gallant, R.C.; Tian, Z.; Wan, J.; Ling, W.; Yang, Y. Cyanidin-3-o-β-Glucoside Induces Megakaryocyte Apoptosis via PI3K/Akt- and MAPKs-Mediated Inhibition of NF-κB Signalling. Thromb. Haemost. 2018, 118, 1215–1229. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yogosawa, S.; Izutani, Y.; Watanabe, H.; Otsuji, E.; Sakai, T. A combination of indol-3-carbinol and genistein synergistically induces apoptosis in human colon cancer HT-29 cells by inhibiting Akt phosphorylation and progression of autophagy. Mol. Cancer 2009, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, K.; Tuzcu, M.; Basak, N.; Caglayan, B.; Kilic, U.; Sahin, F.; Kucuk, O. Sensitization of Cervical Cancer Cells to Cisplatin by Genistein: The Role of NFB and Akt/mTOR Signaling Pathways. J. Oncol. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Cha, H.-J.; Lee, H.; Hwang-Bo, H.; Ji, S.Y.; Kim, M.Y.; Hong, S.H.; Jeong, J.-W.; Han, M.H.; Choi, S.H.; et al. Induction of G2/M Cell Cycle Arrest and Apoptosis by Genistein in Human Bladder Cancer T24 Cells through Inhibition of the ROS-Dependent PI3k/Akt Signal Transduction Pathway. Antioxidants 2019, 8, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanjak, P.; Thiantanawat, A.; Watcharasit, P.; Satayavivad, J. Genistein reduces the activation of AKT and EGFR, and the production of IL6 in cholangiocarcinoma cells involving estrogen and estrogen receptors. Int. J. Oncol. 2018, 53, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Su, H.; Li, Q.; Li, J.; Zhao, Q. Genistein decreases A549 cell viability via inhibition of the PI3K/AKT/HIF-1α/VEGF and NF-κB/COX-2 signaling pathways. Mol. Med. Rep. 2017, 15, 2296–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhu, Y.; Zhang, W.; Peng, X.; Zhou, J.; Li, F.; Han, B.; Liu, X.; Ou, Y.; Yu, X. Delphinidin induced protective autophagy via mTOR pathway suppression and AMPK pathway activation in HER-2 positive breast cancer cells. BMC Cancer 2018, 18, 342. [Google Scholar] [CrossRef]
- Kim, M.-H.; Jeong, Y.-J.; Cho, H.-J.; Hoe, H.-S.; Park, K.-K.; Park, Y.-Y.; Choi, Y.H.; Kim, C.-H.; Chang, H.-W.; Park, Y.-J.; et al. Delphinidin inhibits angiogenesis through the suppression of HIF-1α and VEGF expression in A549 lung cancer cells. Oncol. Rep. 2017, 37, 777–784. [Google Scholar] [CrossRef]
- Lim, W.; Jeong, W.; Song, G. Delphinidin suppresses proliferation and migration of human ovarian clear cell carcinoma cells through blocking AKT and ERK1/2 MAPK signaling pathways. Mol. Cell. Endocrinol. 2016, 422, 172–181. [Google Scholar] [CrossRef]
- Lim, W.; Song, G. Inhibitory effects of delphinidin on the proliferation of ovarian cancer cells via PI3K/AKT and ERK 1/2 MAPK signal transduction. Oncol. Lett. 2017, 14, 810–818. [Google Scholar] [CrossRef] [Green Version]
- Shaw, R.J.; Cantley, L.C. Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature 2006, 441, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Castellano, E.; Downward, J. RAS Interaction with PI3K: More than Just another Effector Pathway. Genes Cancer 2011, 2, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Sjölander, A.; Yamamoto, K.; Huber, B.E.; Lapetina, E.G. Association of p21ras with phosphatidylinositol 3-kinase. Proc. Natl. Acad. Sci. USA 1991, 88, 7908–7912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asati, V.; Mahapatra, D.K.; Bharti, S.K. PI3K/Akt/mTOR and Ras/Raf/MEK/ERK signaling pathways inhibitors as anticancer agents: Structural and pharmacological perspectives. Eur. J. Med. Chem. 2016, 109, 314–341. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Zhou, R.; Kong, Y.; Wang, J.; Xia, W.; Guo, J.; Liu, J.; Sun, H.; Liu, K.; Yang, J.; et al. S-equol, a Secondary Metabolite of Natural Anticancer Isoflavone Daidzein, Inhibits Prostate Cancer Growth In Vitro and In Vivo, Though Activating the Akt/FOXO3a Pathway. Curr. Cancer Drug Targets 2016, 16, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Park, S.; Bazer, F.W.; Lim, W.; Song, G. Myricetin treatment induces apoptosis in canine osteosarcoma cells by inducing DNA fragmentation, disrupting redox homeostasis, and mediating loss of mitochondrial membrane potential. J. Cell. Physiol. 2018, 233, 7457–7466. [Google Scholar] [CrossRef]
- Yang, C.; Lim, W.; Bazer, F.W.; Song, G. Myricetin suppresses invasion and promotes cell death in human placental choriocarcinoma cells through induction of oxidative stress. Cancer Lett. 2017, 399, 10–19. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.-J.; Liu, Y.; Cui, Y.-F. PI3K/AKT/mTOR signaling is involved in (-)-epigallocatechin-3-gallate-induced apoptosis of human pancreatic carcinoma cells. Am. J. Chin. Med. 2013, 41, 629–642. [Google Scholar] [CrossRef]
- Shirakami, Y.; Shimizu, M.; Adachi, S.; Sakai, H.; Nakagawa, T.; Yasuda, Y.; Tsurumi, H.; Hara, Y.; Moriwaki, H. (-)-Epigallocatechin gallate suppresses the growth of human hepatocellular carcinoma cells by inhibiting activation of the vascular endothelial growth factor-vascular endothelial growth factor receptor axis. Cancer Sci. 2009, 100, 1957–1962. [Google Scholar] [CrossRef]
- Park, J.H.; Jin, C.-Y.; Lee, B.K.; Kim, G.-Y.; Choi, Y.H.; Jeong, Y.K. Naringenin induces apoptosis through downregulation of Akt and caspase-3 activation in human leukemia THP-1 cells. Food Chem. Toxicol. 2008, 46, 3684–3690. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Liu, F.; Guo, H.-B.; Li, Y.; Tan, B.-B.; Zhang, W.-X.; Peng, Y.-H. Naringenin inhibits proliferation, migration, and invasion as well as induces apoptosis of gastric cancer SGC7901 cell line by downregulation of AKT pathway. Tumour Biol. 2016, 37, 11365–11374. [Google Scholar] [CrossRef]
- Park, S.; Lim, W.; Bazer, F.W.; Song, G. Naringenin suppresses growth of human placental choriocarcinoma via reactive oxygen species-mediated P38 and JNK MAPK pathways. Phytomedicine 2018, 50, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Mirzoeva, S.; Kim, N.D.; Chiu, K.; Franzen, C.A.; Bergan, R.C.; Pelling, J.C. Inhibition of HIF-1 alpha and VEGF expression by the chemopreventive bioflavonoid apigenin is accompanied by Akt inhibition in human prostate carcinoma PC3-M cells. Mol. Carcinog. 2008, 47, 686–700. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; MacLennan, G.T.; Fu, P.; Gupta, S. Apigenin attenuates insulin-like growth factor-I signaling in an autochthonous mouse prostate cancer model. Pharm. Res. 2012, 29, 1506–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Wang, H.; Jia, Y.; Ding, H.; Zhang, L.; Pan, H. Luteolin reduces migration of human glioblastoma cell lines via inhibition of the p-IGF-1R/PI3K/AKT/mTOR signaling pathway. Oncol. Lett. 2017, 14, 3545–3551. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.T.; Lee, S.H.; Kim, Y.M. Quercetin Regulates Sestrin 2-AMPK-mTOR Signaling Pathway and Induces Apoptosis via Increased Intracellular ROS in HCT116 Colon Cancer Cells. J. Cancer Prev. 2013, 18, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [Green Version]
- Kundu, M.; Lindsten, T.; Yang, C.-Y.; Wu, J.; Zhao, F.; Zhang, J.; Selak, M.A.; Ney, P.A.; Thompson, C.B. Ulk1 plays a critical role in the autophagic clearance of mitochondria and ribosomes during reticulocyte maturation. Blood 2008, 112, 1493–1502. [Google Scholar] [CrossRef] [Green Version]
- Backer, J.M. The regulation and function of Class III PI3Ks: Novel roles for Vps34. Biochem. J. 2008, 410, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.H.; Jackson, S.; Seaman, M.; Brown, K.; Kempkes, B.; Hibshoosh, H.; Levine, B. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature 1999, 402, 672–676. [Google Scholar] [CrossRef]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Dai, Y.; Shi, B.; Yue, F.; Zou, J.; Xu, G.; Jiang, X.; Wang, F.; Zhou, X.; Liu, L. LRPPRC sustains Yap-P27-mediated cell ploidy and P62-HDAC6-mediated autophagy maturation and suppresses genome instability and hepatocellular carcinomas. Oncogene 2020, 39, 3879–3892. [Google Scholar] [CrossRef]
- Takamura, A.; Komatsu, M.; Hara, T.; Sakamoto, A.; Kishi, C.; Waguri, S.; Eishi, Y.; Hino, O.; Tanaka, K.; Mizushima, N. Autophagy-deficient mice develop multiple liver tumors. Genes Dev. 2011, 25, 795–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazova, R.; Camp, R.L.; Klump, V.; Siddiqui, S.F.; Amaravadi, R.K.; Pawelek, J.M. Punctate LC3B expression is a common feature of solid tumors and associated with proliferation, metastasis, and poor outcome. Clin. Cancer Res. 2012, 18, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Ferro, F.; Servais, S.; Besson, P.; Roger, S.; Dumas, J.-F.; Brisson, L. Autophagy and mitophagy in cancer metabolic remodelling. Semin. Cell Dev. Biol. 2020, 98, 129–138. [Google Scholar] [CrossRef]
- Liu, G.; Pei, F.; Yang, F.; Li, L.; Amin, A.D.; Liu, S.; Buchan, J.R.; Cho, W.C. Role of Autophagy and Apoptosis in Non-Small-Cell Lung Cancer. Int. J. Mol. Sci. 2017, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-D.; Qin, Z.-H. Beclin 1, Bcl-2 and Autophagy. Adv. Exp. Med. Biol. 2019, 1206, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhou, J.-Y.; Zhang, D.; Liu, M.-H.; Chen, Y.-G. Artesunate induces apoptosis and autophagy in HCT116 colon cancer cells, and autophagy inhibition enhances the artesunate-induced apoptosis. Int. J. Mol. Med. 2018, 42, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Kardideh, B.; Samimi, Z.; Norooznezhad, F.; Kiani, S.; Mansouri, K. Autophagy, cancer and angiogenesis: Where is the link? Cell Biosci. 2019, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer 2015, 14, 48. [Google Scholar] [CrossRef] [Green Version]
- Ashrafizadeh, M.; Mohammadinejad, R.; Tavakol, S.; Ahmadi, Z.; Roomiani, S.; Katebi, M. Autophagy, anoikis, ferroptosis, necroptosis, and endoplasmic reticulum stress: Potential applications in melanoma therapy. J. Cell. Physiol. 2019, 234, 19471–19479. [Google Scholar] [CrossRef]
- Dower, C.M.; Wills, C.A.; Frisch, S.M.; Wang, H.-G. Mechanisms and context underlying the role of autophagy in cancer metastasis. Autophagy 2018, 14, 1110–1128. [Google Scholar] [CrossRef] [Green Version]
- Musial, C.; Siedlecka-Kroplewska, K.; Kmiec, Z.; Gorska-Ponikowska, M. Modulation of Autophagy in Cancer Cells by Dietary Polyphenols. Antioxidants 2021, 10, 123. [Google Scholar] [CrossRef]
- Pang, X.; Zhang, X.; Jiang, Y.; Su, Q.; Li, Q.; Li, Z. Autophagy: Mechanisms and Therapeutic Potential of Flavonoids in Cancer. Biomolecules 2021, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Herranz, N.; Gil, J. Mechanisms and functions of cellular senescence. J. Clin. Investig. 2018, 128, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.S.; Vats, S.; Chia, A.Y.-Q.; Tan, T.Z.; Deng, S.; Ong, M.S.; Arfuso, F.; Yap, C.T.; Goh, B.C.; Sethi, G.; et al. Dual role of autophagy in hallmarks of cancer. Oncogene 2018, 37, 1142–1158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-W.; Hu, J.-J.; Fu, R.-Q.; Liu, X.; Zhang, Y.-H.; Li, J.; Liu, L.; Li, Y.-N.; Deng, Q.; Luo, Q.-S.; et al. Flavonoids inhibit cell proliferation and induce apoptosis and autophagy through downregulation of PI3Kγ mediated PI3K/AKT/mTOR/p70S6K/ULK signaling pathway in human breast cancer cells. Sci. Rep. 2018, 8, 11255. [Google Scholar] [CrossRef]
- Tomas-Hernández, S.; Blanco, J.; Rojas, C.; Roca-Martínez, J.; Ojeda-Montes, M.J.; Beltrán-Debón, R.; Garcia-Vallvé, S.; Pujadas, G.; Arola, L.; Mulero, M. Resveratrol Potently Counteracts Quercetin Starvation-Induced Autophagy and Sensitizes HepG2 Cancer Cells to Apoptosis. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef]
- Song, J.; Bai, J.; Wang, S.; Liu, L.; Zhao, Z. Effects of Quercetin on Autophagy and Phosphatidylinositol 3-kinase/Protein Kinase B/Mammalian Target of Rapamycin Signaling Pathway in Human Prostate Cancer PC-3 Cells. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2020, 42, 578–584. [Google Scholar] [CrossRef]
- Han, B.; Yu, Y.-Q.; Yang, Q.-L.; Shen, C.-Y.; Wang, X.-J. Kaempferol induces autophagic cell death of hepatocellular carcinoma cells via activating AMPK signaling. Oncotarget 2017, 8, 86227–86239. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Chang, H.; Deng, S.; Fan, D. Icariin enhances the chemosensitivity of cisplatin-resistant ovarian cancer cells by suppressing autophagy via activation of the AKT/mTOR/ATG5 pathway. Int. J. Oncol. 2019, 54, 1933–1942. [Google Scholar] [CrossRef]
- Zhou, C.; Gu, J.; Zhang, G.; Dong, D.; Yang, Q.; Chen, M.-B.; Xu, D. AMPK-autophagy inhibition sensitizes icaritin-induced anti-colorectal cancer cell activity. Oncotarget 2017, 8, 14736–14747. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Liu, C.-F.; Gao, N.; Zhao, J.; Xu, J. Kaempferol suppresses proliferation but increases apoptosis and autophagy by up-regulating microRNA-340 in human lung cancer cells. Biomed. Pharmacother. 2018, 108, 809–816. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, X.; Chang, H.; Shu, F.; Wu, Y.; Zhang, T.; Fu, Y.; Zhang, Q.; Zhu, J.-D.; Mi, M. Ampelopsin-induced autophagy protects breast cancer cells from apoptosis through Akt-mTOR pathway via endoplasmic reticulum stress. Cancer Sci. 2014, 105, 1279–1287. [Google Scholar] [CrossRef]
- Zhu, M.-L.; Zhang, P.-M.; Jiang, M.; Yu, S.-W.; Wang, L. Myricetin induces apoptosis and autophagy by inhibiting PI3K/Akt/mTOR signalling in human colon cancer cells. BMC Complement. Med. Ther. 2020, 20, 209. [Google Scholar] [CrossRef]
- Zhou, D.-Z.; Sun, H.-Y.; Yue, J.-Q.; Peng, Y.; Chen, Y.-M.; Zhong, Z.-J. Dihydromyricetin induces apoptosis and cytoprotective autophagy through ROS-NF-κB signalling in human melanoma cells. Free Radic. Res. 2017, 51, 517–528. [Google Scholar] [CrossRef]
- Zou, M.; Hu, C.; You, Q.; Zhang, A.; Wang, X.; Guo, Q. Oroxylin A induces autophagy in human malignant glioma cells via the mTOR-STAT3-Notch signaling pathway. Mol. Carcinog. 2015, 54, 1363–1375. [Google Scholar] [CrossRef]
- Zou, M.; Lu, N.; Hu, C.; Liu, W.; Sun, Y.; Wang, X.; You, Q.; Gu, C.; Xi, T.; Guo, Q. Beclin 1-mediated autophagy in hepatocellular carcinoma cells: Implication in anticancer efficiency of oroxylin A via inhibition of mTOR signaling. Cell. Signal. 2012, 24, 1722–1732. [Google Scholar] [CrossRef]
- Zheng, J.; Shao, Y.; Jiang, Y.; Chen, F.; Liu, S.; Yu, N.; Zhang, D.; Liu, X.; Zou, L. Tangeretin inhibits hepatocellular carcinoma proliferation and migration by promoting autophagy-related BECLIN1. Cancer Manag. Res. 2019, 11, 5231–5242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Wei, S.; Wang, J.; Liu, X. Isoorientin induces apoptosis and autophagy simultaneously by reactive oxygen species (ROS)-related p53, PI3K/Akt, JNK, and p38 signaling pathways in HepG2 cancer cells. J. Agric. Food Chem. 2014, 62, 5390–5400. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kwon, Y.H. Regulation of apoptosis and autophagy by luteolin in human hepatocellular cancer Hep3B cells. Biochem. Biophys. Res. Commun. 2019, 517, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Verschooten, L.; Barrette, K.; Van Kelst, S.; Rubio Romero, N.; Proby, C.; De Vos, R.; Agostinis, P.; Garmyn, M. Autophagy inhibitor chloroquine enhanced the cell death inducing effect of the flavonoid luteolin in metastatic squamous cell carcinoma cells. PLoS ONE 2012, 7, e48264. [Google Scholar] [CrossRef] [PubMed]
- Raha, S.; Yumnam, S.; Hong, G.E.; Lee, H.J.; Saralamma, V.V.G.; Park, H.-S.; Heo, J.D.; Lee, S.J.; Kim, E.H.; Kim, J.-A.; et al. Naringin induces autophagy-mediated growth inhibition by downregulating the PI3K/Akt/mTOR cascade via activation of MAPK pathways in AGS cancer cells. Int. J. Oncol. 2015, 47, 1061–1069. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-Y.; Park, Y.-J.; Hwang, S.-C.; Kim, K.-D.; Moon, D.-K.; Kim, D.-H. Cytotoxic effects of delphinidin in human osteosarcoma cells. Acta Orthop. Traumatol. Turc. 2018, 52, 58–64. [Google Scholar] [CrossRef]
- Kang, S.H.; Bak, D.-H.; Chung, B.Y.; Bai, H.-W.; Kang, B.S. Delphinidin enhances radio-therapeutic effects via autophagy induction and JNK/MAPK pathway activation in non-small cell lung cancer. Korean J. Physiol. Pharmacol. 2020, 24, 413–422. [Google Scholar] [CrossRef]
- Chiu, C.-T.; Hsuan, S.-W.; Lin, H.-H.; Hsu, C.-C.; Chou, F.-P.; Chen, J.-H. Hibiscus sabdariffa leaf polyphenolic extract induces human melanoma cell death, apoptosis, and autophagy. J. Food Sci. 2015, 80, H649–H658. [Google Scholar] [CrossRef]
- Enkhbat, T.; Nishi, M.; Yoshikawa, K.; Jun, H.; Tokunaga, T.; Takasu, C.; Kashihara, H.; Ishikawa, D.; Tominaga, M.; Shimada, M. Epigallocatechin-3-gallate Enhances Radiation Sensitivity in Colorectal Cancer Cells Through Nrf2 Activation and Autophagy. Anticancer Res. 2018, 38, 6247–6252. [Google Scholar] [CrossRef]
- Izdebska, M.; Klimaszewska-Wiśniewska, A.; Hałas, M.; Gagat, M.; Grzanka, A. Green tea extract induces protective autophagy in A549 non-small lung cancer cell line. Postepy Hig. Med. Dosw. 2015, 69, 1478–1484. [Google Scholar]
- Hu, F.; Wei, F.; Wang, Y.; Wu, B.; Fang, Y.; Xiong, B. EGCG synergizes the therapeutic effect of cisplatin and oxaliplatin through autophagic pathway in human colorectal cancer cells. J. Pharmacol. Sci. 2015, 128, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.-H.; Horng, C.-T.; Lee, C.-F.; Chiang, N.-N.; Tsai, F.-J.; Lu, C.-C.; Chiang, J.-H.; Hsu, Y.-M.; Yang, J.-S.; Chen, F.-A. Epigallocatechin gallate sensitizes cisplatin-resistant oral cancer CAR cell apoptosis and autophagy through stimulating AKT/STAT3 pathway and suppressing multidrug resistance 1 signaling. Environ. Toxicol. 2017, 32, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Prietsch, R.F.; Monte, L.G.; da Silva, F.A.; Beira, F.T.; Del Pino, F.A.B.; Campos, V.F.; Collares, T.; Pinto, L.S.; Spanevello, R.M.; Gamaro, G.D.; et al. Genistein induces apoptosis and autophagy in human breast MCF-7 cells by modulating the expression of proapoptotic factors and oxidative stress enzymes. Mol. Cell. Biochem. 2014, 390, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cook, K.L.; Warri, A.; Cruz, I.M.; Rosim, M.; Riskin, J.; Helferich, W.; Doerge, D.; Clarke, R.; Hilakivi-Clarke, L. Lifetime Genistein Intake Increases the Response of Mammary Tumors to Tamoxifen in Rats. Clin. Cancer Res. 2017, 23, 814–824. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Jin, F.; Lian, X.; Li, M.; Wang, G.; Lan, B.; He, H.; Liu, G.-D.; Wu, Y.; Sun, G.; et al. Genistein promotes ionizing radiation-induced cell death by reducing cytoplasmic Bcl-xL levels in non-small cell lung cancer. Sci. Rep. 2018, 8, 328. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Kang, Y.; Li, X.; Roife, D.; Zhang, R.; Fleming, J.B. Genistein potentiates the antitumor effect of 5-Fluorouracil by inducing apoptosis and autophagy in human pancreatic cancer cells. Anticancer Res. 2014, 34, 4685–4692. [Google Scholar]
- Nazim, U.M.; Park, S.-Y. Genistein enhances TRAIL-induced cancer cell death via inactivation of autophagic flux. Oncol. Rep. 2015, 34, 2692–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.-Y.; Zhu, Y.; Li, X.-F.; Wang, X.-Q.; Tang, L.-P.; Su, Z.-Q.; Li, C.-Y.; Zheng, G.-J.; Feng, B. Scutellarin Increases Cisplatin-Induced Apoptosis and Autophagy to Overcome Cisplatin Resistance in Non-small Cell Lung Cancer via ERK/p53 and c-met/AKT Signaling Pathways. Front. Pharmacol. 2018, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Shanmugam, M.K.; Kumar, A.P.; Yap, C.T.; Sethi, G.; Bishayee, A. Targeting autophagy using natural compounds for cancer prevention and therapy. Cancer 2019, 125, 1228–1246. [Google Scholar] [CrossRef]
- Huang, T.; Song, X.; Yang, Y.; Wan, X.; Alvarez, A.A.; Sastry, N.; Feng, H.; Hu, B.; Cheng, S.-Y. Autophagy and Hallmarks of Cancer. Crit. Rev. Oncog. 2018, 23, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Das, C.K.; Banerjee, I.; Mandal, M. Pro-survival autophagy: An emerging candidate of tumor progression through maintaining hallmarks of cancer. Semin. Cancer Biol. 2020, 66, 59–74. [Google Scholar] [CrossRef]
- Mohammad, R.M.; Muqbil, I.; Lowe, L.; Yedjou, C.; Hsu, H.-Y.; Lin, L.-T.; Siegelin, M.D.; Fimognari, C.; Kumar, N.B.; Dou, Q.P.; et al. Broad targeting of resistance to apoptosis in cancer. Semin. Cancer Biol. 2015, 35, S78–S103. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [Green Version]
- Abotaleb, M.; Samuel, S.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, J.H. Kaempferol Inhibits Pancreatic Cancer Cell Growth and Migration through the Blockade of EGFR-Related Pathway In Vitro. PLoS ONE 2016, 11, e0155264. [Google Scholar] [CrossRef] [Green Version]
- Pan, F.; Liu, Y.; Liu, J.; Wang, E. Stability of blueberry anthocyanin, anthocyanidin and pyranoanthocyanidin pigments and their inhibitory effects and mechanisms in human cervical cancer HeLa cells. RSC Adv. 2019, 9, 10842–10853. [Google Scholar] [CrossRef] [Green Version]
- Shafiee, G.; Saidijam, M.; Tavilani, H.; Ghasemkhani, N.; Khodadadi, I. Genistein Induces Apoptosis and Inhibits Proliferation of HT29 Colon Cancer Cells. Int. J. Mol. Cell. Med. 2016, 5, 178–191. [Google Scholar]
- Huang, L.; Jin, K.; Lan, H. Luteolin inhibits cell cycle progression and induces apoptosis of breast cancer cells through downregulation of human telomerase reverse transcriptase. Oncol. Lett. 2019, 17, 3842–3850. [Google Scholar] [CrossRef] [Green Version]
- Arul, D.; Subramanian, P. Naringenin (citrus flavonone) induces growth inhibition, cell cycle arrest and apoptosis in human hepatocellular carcinoma cells. Pathol. Oncol. Res. 2013, 19, 763–770. [Google Scholar] [CrossRef]
- Kim, M.E.; Ha, T.K.; Yoon, J.H.; Lee, J.S. Myricetin induces cell death of human colon cancer cells via BAX/BCL2-dependent pathway. Anticancer Res. 2014, 34, 701–706. [Google Scholar]
- Shukla, S.; Fu, P.; Gupta, S. Apigenin induces apoptosis by targeting inhibitor of apoptosis proteins and Ku70-Bax interaction in prostate cancer. Apoptosis 2014, 19, 883–894. [Google Scholar] [CrossRef]
- Hsiao, Y.-C.; Peng, S.-F.; Lai, K.-C.; Liao, C.-L.; Huang, Y.-P.; Lin, C.-C.; Lin, M.-L.; Liu, K.-C.; Tsai, C.-C.; Ma, Y.-S.; et al. Genistein induces apoptosis in vitro and has antitumor activity against human leukemia HL-60 cancer cell xenograft growth in vivo. Environ. Toxicol. 2019, 34, 443–456. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, T.; Sun, R.; Yang, L.; An, R.; Xue, Y. Daidzein induces choriocarcinoma cell apoptosis in a dose-dependent manner via the mitochondrial apoptotic pathway. Mol. Med. Rep. 2018, 17, 6093–6099. [Google Scholar] [CrossRef] [PubMed]
- Razak, S.; Afsar, T.; Ullah, A.; Almajwal, A.; Alkholief, M.; Alshamsan, A.; Jahan, S. Taxifolin, a natural flavonoid interacts with cell cycle regulators causes cell cycle arrest and causes tumor regression by activating Wnt/β -catenin signaling pathway. BMC Cancer 2018, 18, 1043. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Zhu, G.; Jia, N.; Yang, W. Epigallocatechin-3-gallate Sensitizes Human 786-O Renal Cell Carcinoma Cells to TRAIL-Induced Apoptosis. Cell Biochem. Biophys. 2015, 72, 157–164. [Google Scholar] [CrossRef]
- Han, B.-J.; Li, W.; Jiang, G.-B.; Lai, S.-H.; Zhang, C.; Zeng, C.-C.; Liu, Y.-J. Effects of daidzein in regards to cytotoxicity in vitro, apoptosis, reactive oxygen species level, cell cycle arrest and the expression of caspase and Bcl-2 family proteins. Oncol. Rep. 2015, 34, 1115–1120. [Google Scholar] [CrossRef]
- Srivastava, S.; Somasagara, R.R.; Hegde, M.; Nishana, M.; Tadi, S.K.; Srivastava, M.; Choudhary, B.; Raghavan, S.C. Quercetin, a Natural Flavonoid Interacts with DNA, Arrests Cell Cycle and Causes Tumor Regression by Activating Mitochondrial Pathway of Apoptosis. Sci. Rep. 2016, 6, 24049. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Polier, G.; Köhler, R.; Giaisi, M.; Krammer, P.H.; Li-Weber, M. Wogonin and related natural flavones overcome tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) protein resistance of tumors by down-regulation of c-FLIP protein and up-regulation of TRAIL receptor 2 expression. J. Biol. Chem. 2012, 287, 641–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, S. Linking the Cell Cycle to Cell Fate Decisions. Trends Cell Biol. 2015, 25, 592–600. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K. Development of cell-cycle checkpoint therapy for solid tumors. Jpn. J. Clin. Oncol. 2015, 45, 1097–1102. [Google Scholar] [CrossRef] [Green Version]
- De Felice, F.; Marchetti, C.; Tombolini, V.; Panici, P.B. Immune check-point in endometrial cancer. Int. J. Clin. Oncol. 2019, 24, 910–916. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, C.-Z.; Du, G.-J.; Qi, L.-W.; Calway, T.; He, T.-C.; Du, W.; Yuan, C.-S. Genistein induces G2/M cell cycle arrest and apoptosis via ATM/p53-dependent pathway in human colon cancer cells. Int. J. Oncol. 2013, 43, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; van Gent, D.C.; Incrocci, L.; van Weerden, W.M.; Nonnekens, J. Role of the DNA damage response in prostate cancer formation, progression and treatment. Prostate Cancer Prostatic Dis. 2020, 23, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.-Y.; Zupkó, I.; Chang, F.-R.; Hunyadi, A.; Wu, C.-C.; Weng, T.-S.; Wang, H.-C. Dietary flavonoid derivatives enhance chemotherapeutic effect by inhibiting the DNA damage response pathway. Toxicol. Appl. Pharmacol. 2016, 311, 99–105. [Google Scholar] [CrossRef]
- Mavrogonatou, E.; Pratsinis, H.; Kletsas, D. The role of senescence in cancer development. Semin. Cancer Biol. 2020, 62, 182–191. [Google Scholar] [CrossRef]
- Campisi, J. Aging, Cellular Senescence, and Cancer. Annu. Rev. Physiol. 2013, 75, 685–705. [Google Scholar] [CrossRef] [Green Version]
- Davalos, A.R.; Coppe, J.-P.; Campisi, J.; Desprez, P.-Y. Senescent cells as a source of inflammatory factors for tumor progression. Cancer Metastasis Rev. 2010, 29, 273–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef]
- Özsoy, S.; Becer, E.; Kabadayı, H.; Vatansever, H.S.; Yücecan, S. Quercetin-Mediated Apoptosis and Cellular Senescence in Human Colon Cancer. Anticancer. Agents Med. Chem. 2020, 20, 1387–1396. [Google Scholar] [CrossRef]
- Udroiu, I.; Marinaccio, J.; Sgura, A. Epigallocatechin-3-gallate induces telomere shortening and clastogenic damage in glio-blastoma cells. Environ. Mol. Mutagen. 2019, 60, 683–692. [Google Scholar] [CrossRef]
- Banerjee, K.; Mandal, M. Oxidative stress triggered by naturally occurring flavone apigenin results in senescence and chemotherapeutic effect in human colorectal cancer cells. Redox Biol. 2015, 5, 153–162. [Google Scholar] [CrossRef] [Green Version]
- León-González, A.J.; Sharif, T.; Auger, C.; Abbas, M.; Fuhrmann, G.; Schini-Kerth, V.B. Anthocyanin-rich bilberry extract induces apoptosis in acute lymphoblastic leukemia cells via redox-sensitive epigenetic modifications. J. Funct. Foods. 2018, 44, 227–234. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, E.H.; Park, S.S.; Lim, J.H.; Kwon, T.K.; Choi, K.S. Quercetin sensitizes human hepatoma cells to TRAIL-induced apoptosis via Sp1-mediated DR5 up-regulation and proteasome-mediated c-FLIPS down-regulation. J. Cell. Biochem. 2008, 105, 1386–1398. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Kim, G.-H. Daidzein causes cell cycle arrest at the G1 and G2/M phases in human breast cancer MCF-7 and MDA-MB-453 cells. Phytomedicine 2008, 15, 683–690. [Google Scholar] [CrossRef] [PubMed]







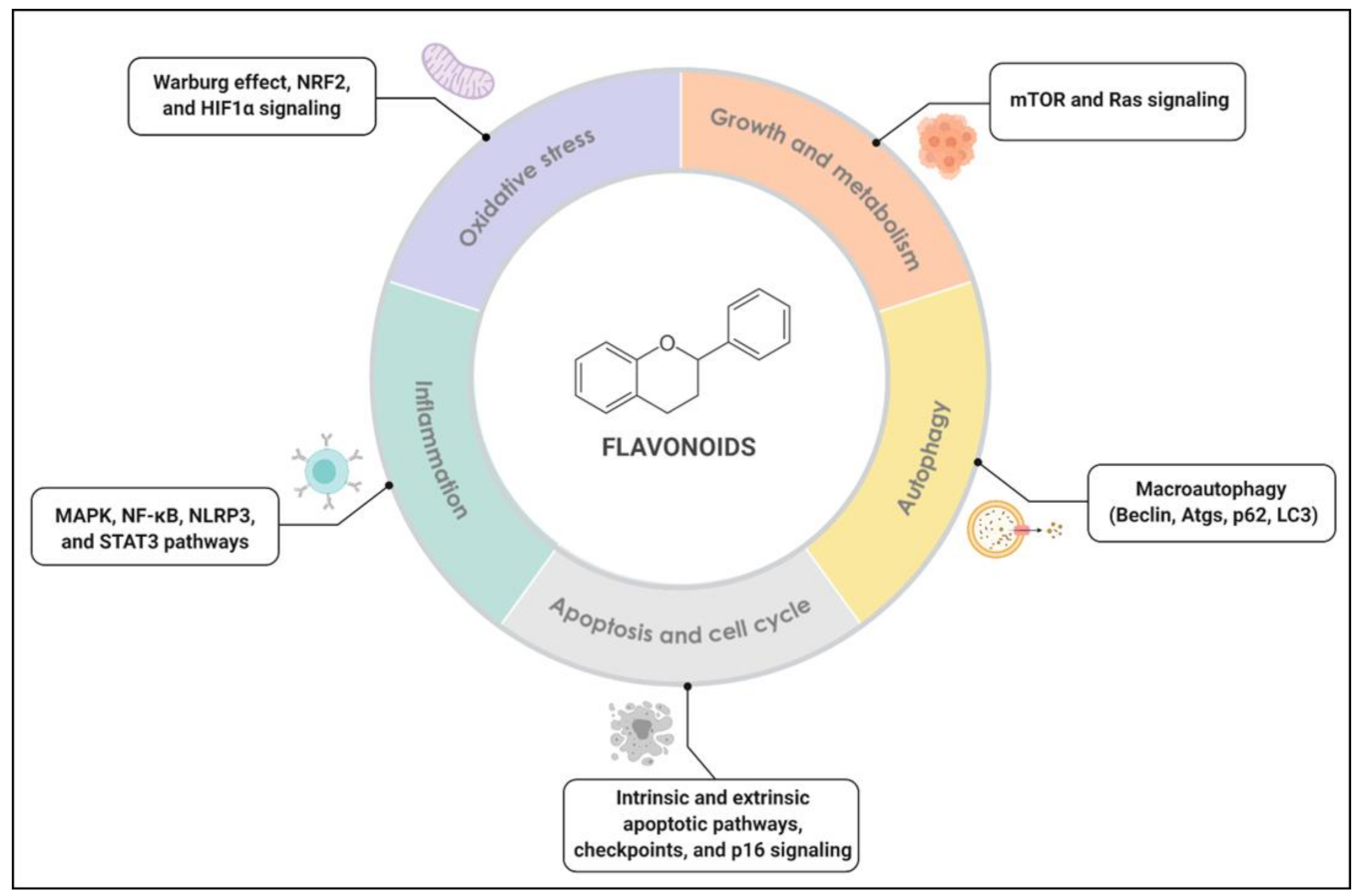{kind=link}
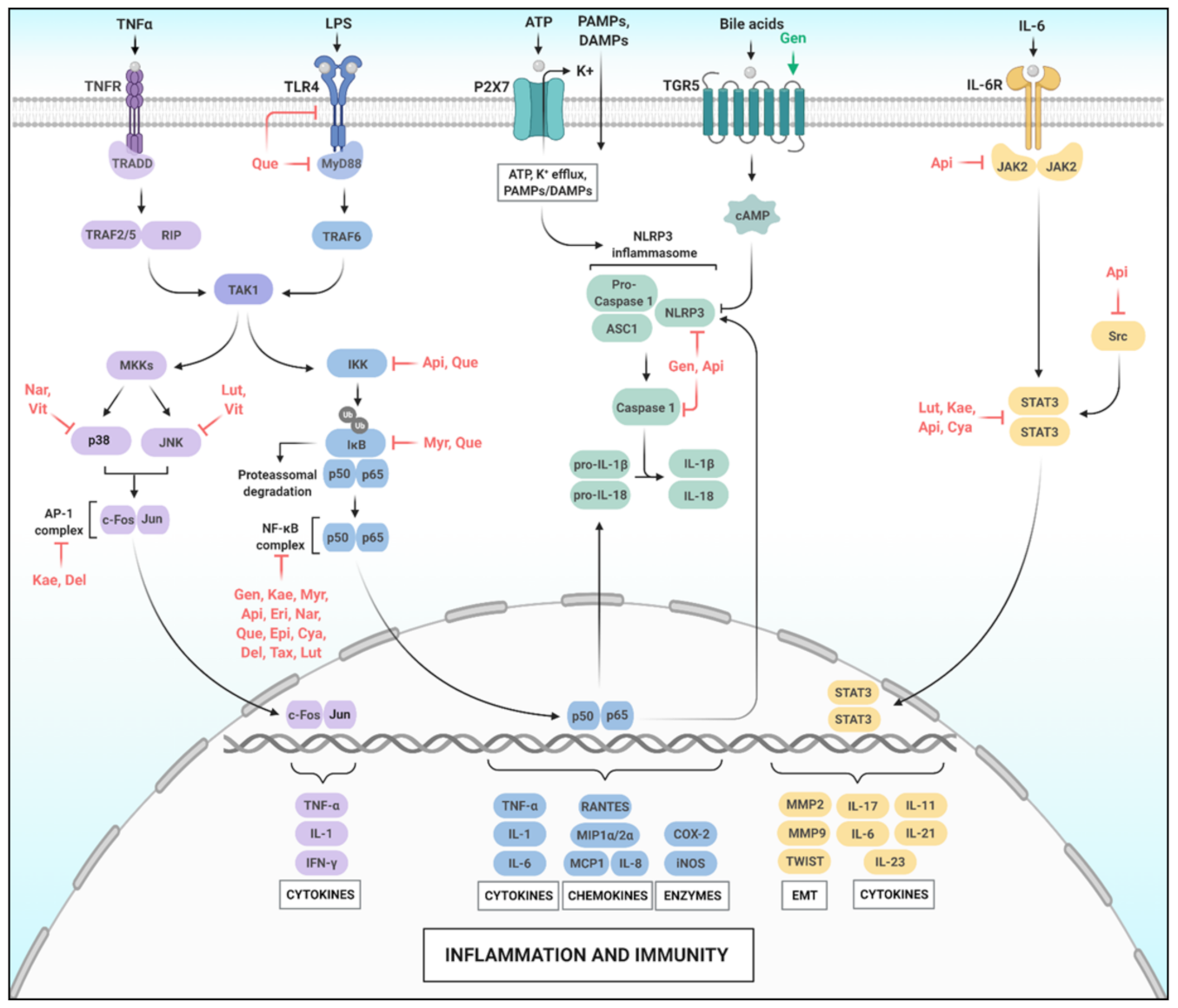{kind=link}
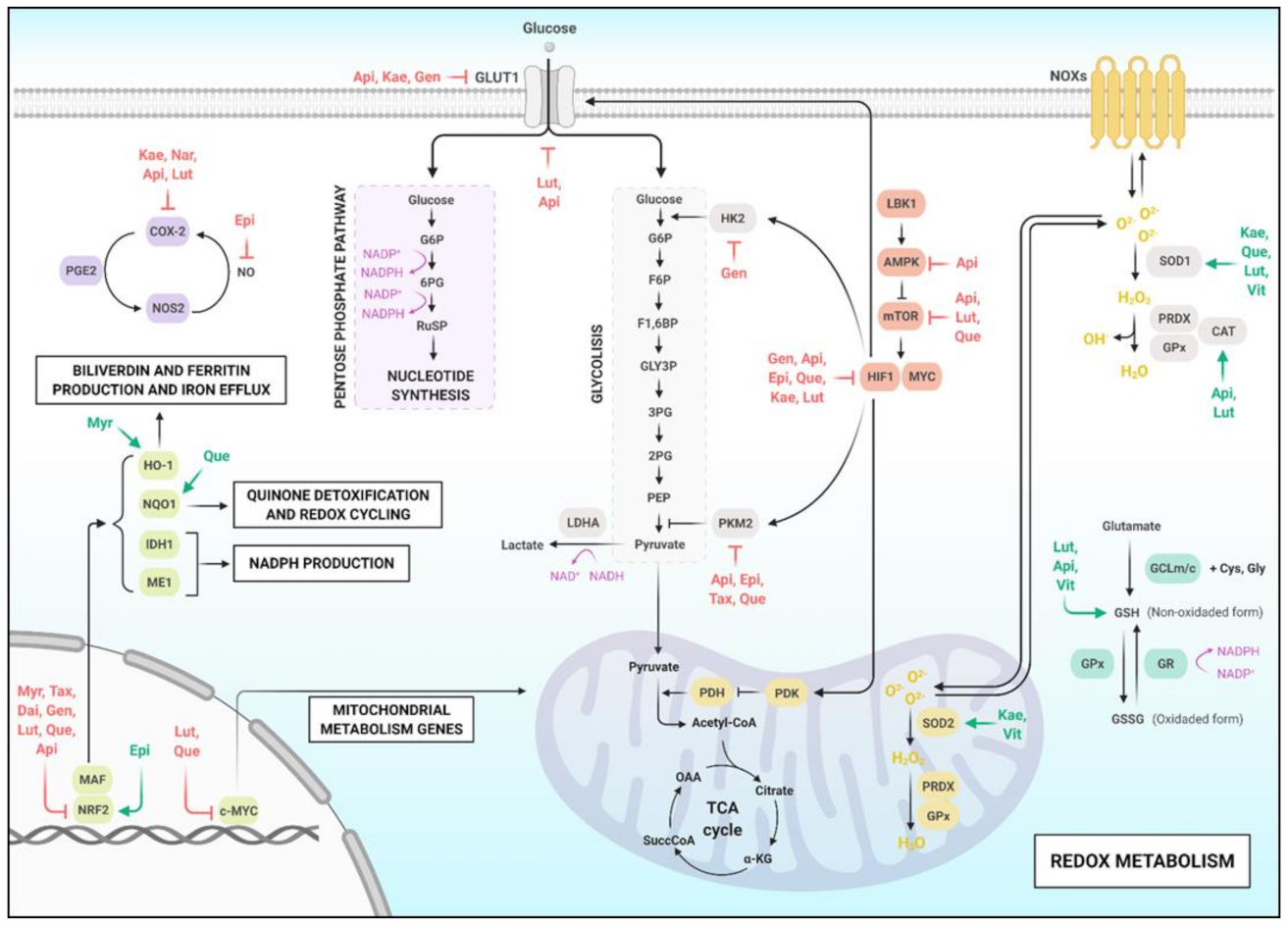{kind=link}
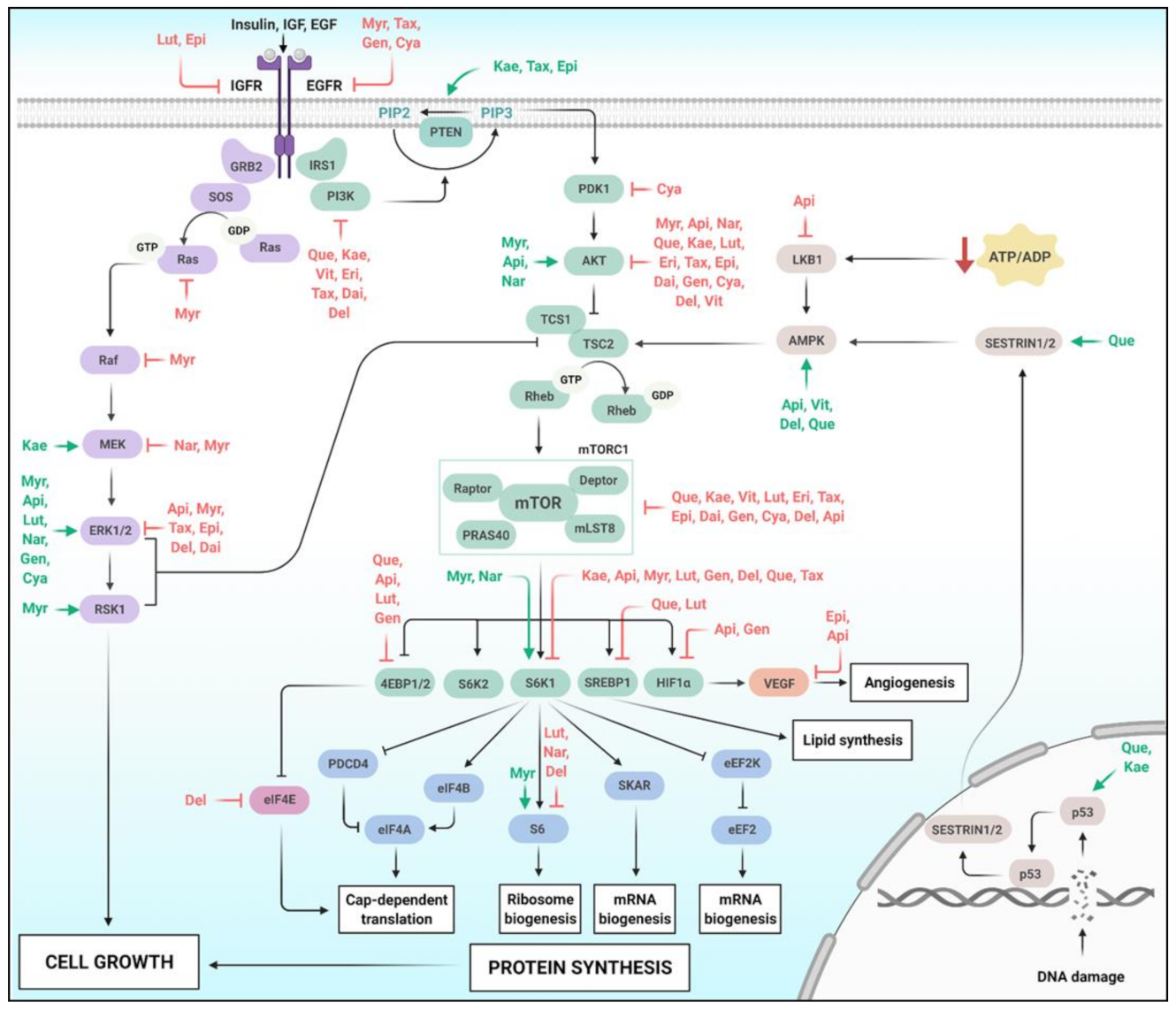{kind=link}
{kind=link}
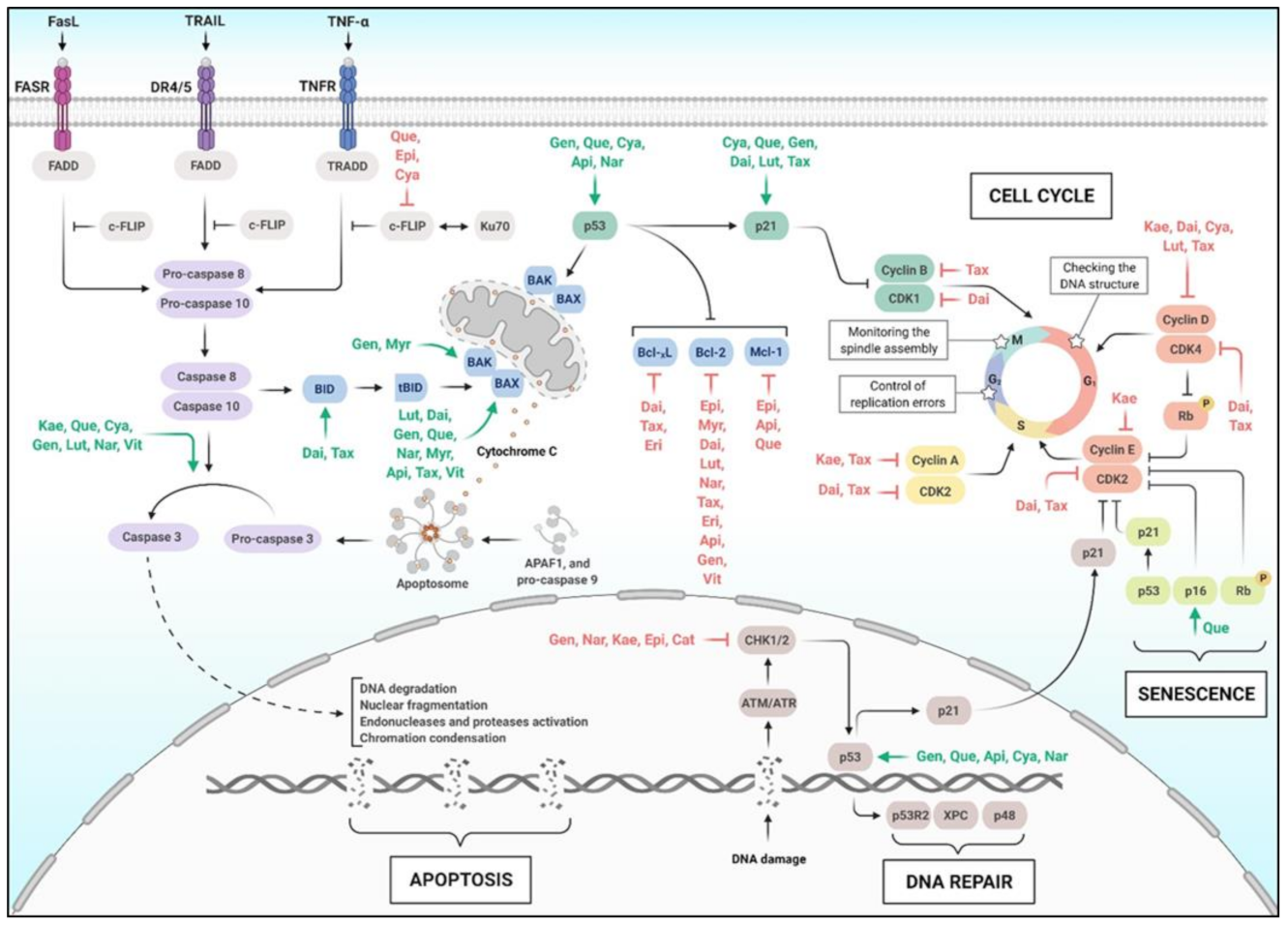{kind=link}
Chemical Structure | Subclass | Examples | Food Source |
|---|---|---|---|
 | Anthocyanidins | Cyanidin (Cya), Delphinidin (Del) | Strawberry, Blackberry, Grape, Red cabbage [8] |
 | Flavanols | Catechin (Cat), Epigallocatechin (Epi) | Tea, Apples, Cocoa [9] |
 | Flavanones | Naringenin (Nar), Taxifolin (Tax), Eriodictyol (Eri) | Citrus fruits [10] |
 | Flavones | Luteolin (Lut), Apigenin (Api), Vitexin (Vit) | Celery, Parsley, Artichokes, Chicory, Tea and Herbs leaves [11] |
 | Flavonols | Quercetin (Que), Kaempferol (Kae), Myricetin (Myr) | Broccoli, Garlic, Onion [12] |
 | Isoflavones | Genistein (Gen), Daidzein (Dai) | Soy [13] |
| Anthocyanidins | ||||||
|---|---|---|---|---|---|---|
| Indication | Number of Participants | FDA Approval Status | Trial Status | National Clinical Trial Number (NCT#) | Ref. | |
| Cyanidin | Breast Cancer | 300 | Not applicable | Completed | NCT02195960 | - |
| Colorectal Adenocarcinoma | 100 | Phase II | Unknown | NCT01948661 | - | |
| Myelodysplastic Syndrome/ Myeloproliferative Neoplasm | 21 | Phase II | Active, not recruiting | NCT03140280 | - | |
| Oral Cancer | 58 | Not Applicable | Not yet recruiting | NCT04372914 | - | |
| Flavanols | ||||||
| Indication | Number of Participants | FDA Approval Status | Trial Status | NCT# | Ref. | |
| Catechins | Prostate Cancer | 50 | Phase I | Completed | NCT00459407 | [19] |
| Breast Cancer | 40 | Phase I | Completed | NCT00516243 | [20] | |
| Breast Cancer | 1075 | Phase II | Completed | NCT00917735 | [21] | |
| Lung Cancer | 53 | Phase II | Completed | NCT00573885 | - | |
| Unspecified Adult Solid Tumor, | Unspecified | Phase I | Completed | NCT00091325 | - | |
| Esophageal Cancer | 55 | Phase I | Completed | NCT00233935 | - | |
| Cervical Cancer | 98 | Phase II | Completed | NCT00303823 | [22] | |
| Bladder | 31 | Phase II | Completed | NCT00666562 | - | |
| Flavanones | ||||||
| Indication | Number of Participants | FDA Approval Status | Trial Status | NCT# | Ref. | |
| Hesperidin | Breast Cancer | 40 | - | Completed | NCT03482401 | [23] |
| Flavones | ||||||
| Indication | Number of Participants | FDA Approval Status | Trial Status | NCT# | Ref. | |
| Apigenin | Colorectal Cancer | 382 | Phase II | Suspended | NCT00609310 | - |
| Luteolin | Tongue Neoplasms | 4 | Early phase I | Unknown | NCT03288298 | - |
| Flavonols | ||||||
| Indication | Number of Participants | FDA Approval Status | Trial Status | NCT# | Ref. | |
| Quercetin | Prostate Cancer | 31 | Phase I | Active, not recruiting | NCT01912820 | - |
| Prostate Cancer | 60 | Not applicable | Recruiting | NCT01538316 | - | |
| Squamous Cell Carcinoma | 55 | Phase II | Recruiting | NCT03476330 | - | |
| Quercetin-3-O-glucoside | Colorectal and Pancreatic Cancer | 64 | Phase II/III | Active, not recruiting | NCT02195232 | - |
| Renal Cell Carcinoma and Kidney Cancer | 104 | Phase I/II | Recruiting | NCT02446795 | - | |
| Isoflavones | ||||||
| Indication | Number of Participants | FDA Approval Status | Trial Status | NCT# | Ref. | |
| Genistein | Bladder Cancer | 60 | Phase II | Active, not recruiting | NCT00118040 | [24] |
| Colon and Rectal Cancer | 13 | Phase I/II | Completed | NCT01985763 | [25] | |
| Refractory Leukemias, Central Nervous System Tumor, Solid Tumor, Refractory Solid Tumor, Leukemia, and Lymphoma | 6 | Phase I/II | Completed | NCT02499861 | - | |
| Non-small Cell Lung Cancer | 21 | Phase I/II | Active, not recruiting | NCT02567799 | [26] | |
| Adenocarcinoma | 44 | Phase I/II | Unknown | NCT01182246 | - | |
| Cancer | 15 | Phase I | Completed | NCT00001696 | - | |
| Prostate Cancer | 24 | Phase II | Temporarily suspended | NCT02766478 | - | |
| Prostatic Neoplasms | 47 | Phase II | Unknown | NCT00546039 | - | |
| Breast Cancer | 126 | Phase II | Completed | NCT00290758 | - | |
| Prostate Cancer | 15 | Phase II | Completed | NCT01325311 | - | |
| Daidzein | Prostate Cancer | 43 | Phase II | Completed | NCT00669656 | - |
| Inflammation and Immunity | ||||||
|---|---|---|---|---|---|---|
| Molecular Mechanisms | Concentration | Incubation | Model | Observations | References | |
| Cyanidin | Inhibits nucleus translocation of NF-κB. Reduces COX-2 and iNOS | 250 and 500 μM | 24 h | SKH-1 hairless mice | Cyanidin was topically administrated, followed by exposition to UV radiation | [58] |
| Reduces p-STAT3 | 100 and 200 μM | 4 h | Peripheral blood mononuclear cells | Stimulated with IL-6 | [115] | |
| Delphinidin | Suppresses MMP9 expression, activation of NF-κB and AP-1 | 60 μM | 24 h | MCF-7 cells | Co-treatment with Phorbol Myristate Acetate (PMA) | [59] |
| Epigallocatechin | Inhibits NF-κB, MMP-2, and MMP-9 activity. Reduces cytokines and chemokines production | 40 μg/mL | 24 h | LnCAP, DU-145, and PC-3 cells | Followed by CpG-ODN or TNFα treatment | [57] |
| Eriodictyol | Inhibits NF-κB pathway | 100 µM | 48 h | U87MG and CHG-5 cells | - | [62] |
| Naringenin | Down-regulates NF-κB pathway and TNF-α, IL-1β, and IL-6 production | 50 mg/kg b.wt | 16 weeks | Swiss albino mice | B[a]P induced lung carcinogenesis in vivo | [65] |
| Down-regulates MMP2, MMP9, and p38 activity | 200 and 300 µM | 24 h | 8901 and 8401 GBM cells | - | [86] | |
| Apigenin | Inhibits IL-6, IL-1β, TNF-α, NF-κB, caspase-1, and NLRP3 assembly | 25 µM | 2 h | THP-1-derived macrophages and HEK293 cells | Followed by IL-1β or TNF-α or LPS treatment | [64] |
| Reduces phosphorylation of src, JAK2, and STAT3 | 20–40 µM | 24 h | HL60 and TF1 cells | - | [112] | |
| Inhibits IKK activation and suppresses NF-κB activation | 20 and 50 μg/mouse/day | 20 weeks | TRAMP mice, oral gavage | - | [54] | |
| Inhibits STAT3 activity, reduces MMP-2, MMP-9, VEGF, and TWIST expression | 40 µM in cells or 150 mg/kg/day in mice | 24 h in cells or 24 days in mice | A375, and G361 cells. Lung metastasis in C57BL/6 mice | - | [113] | |
| Vitexin | Reduces phosphorylation of p38 and JNK | 25, 50, and 100 μg/mL | 2 h | RAW 264.7 cells | Stimulated with LPS | [87] |
| Luteolin | Attenuates TNF-α, IL-8, IL-6, and COX-2 expression. Decreases p-JNK 1/2. Inhibits NF-κB activation and IκB degradation | 50 µM | 1 h | HMC-1 cells | Stimulation with PMA plus A23187 | [63] |
| Deactivates STAT3 Reduces MMP2, MMP7, and MMP9 levels | 80 µM | 24 h | PANC-1 and SW1990 cells | - | [110] | |
| Kaempferol | Inhibits NF-κB activity | 10 µM | 72 h | Jurkat cells | TNF-α treatment | [53] |
| Reduces p-STAT3 | 75 µM | 72 h | LS174 cells | Alone or combined with 5-FU | [111] | |
| Inhibits NF-κB activity and c-Fos expression | 25, 50, or 100 µM | 1 h | RAW264.7 cells | Followed by LPS treatment | [60] | |
| Myricetin | Reduces TNF-α, IL-1β, IL-6, NF-κB, p-NF-κB, and COX-2 | 40 and 100 mg/kg | 4 weeks | AOM/DSS-induced colitis and tumorigenesis in model mice | - | [61] |
| Quercetin | Down-regulates TLR4 and NF-κB pathways. Reduces secretion of IL-1β, IL-6, TNF-α, and IL-10 | 5 μM | 4 h | A549 cells | Followed by nickel exposition | [56] |
| Inhibits NF-κB nuclear translocation | 50 and 100 μM | 24 h | ACC-2 and ACC-M cells | - | [67] | |
| Down-regulates NF-κB, p-IκB-α, p-IKK-β, and cyclin D1 | 80 μM | 24 h | HeLa cells | - | [55] | |
| Genistein | Inhibits NLRP3 inflammasome via TGR5-cAMP | 20 µM | 2 h | THP-1 and U937 cells | Followed by LPS and ATP treatment | [100] |
| Inhibits NLRP3 and NF-κB pathway | 1, 5, and 10 μM | 1 h | RAW264.7 cells | Followed by LPS treatment | [101] | |
| Inhibits NF-κB activity | 10 μM | 30–120 min | HCT116 cells | Combined LPS treatment | [52] | |
| Taxifolin | Down-regulates NF-κB, TNF-α, COX-2, and cyclin D1 | 4 μg/kg | 15 weeks | Swiss albino mice | Pre-treated with 1,2-Dimethylhydrazine (DMH) | [66] |
| Redox Metabolism | ||||||
|---|---|---|---|---|---|---|
| Molecular Mechanisms | Concentration | Incubation | Model | Observations | References | |
| Epigallocatechin | Acts as a chemosensitizer in breast cancer cells by activating NRF2 signaling | 25 µM | 24 h | HeLa cells | Observed in Cisplatin triple-negative cells | [167] |
| Inhibits cell proliferation under hypoxia via the downregulation of HIF-1α and its downstream target gene VEGF levels | 80 μg/mL | 48 h | SGC7901 cells | - | [182] | |
| Naringenin | Suppresses the early stage of colon cancer by attenuating iNOS and COX-2 levels | 200 mg/kg | 10 weeks | Sprague-Dawley rats injected with a carcinogen | - | [188] |
| Apigenin | Decreases PKM2 expression | 10–60 µM | 24 h | HCT116, HT29, and DLD1 cells | - | [148] |
| Reduces cell viability in a dose- and time-dependent manner through CAT and GSH activity | 100 μmol/L | 24–48 h | HepG2 cells | Observed in doxorubicin-resistant hepatocarcinoma | [171] | |
| Down-regulates of HIF1α and GLUT-1 mRNA expression. Represses hypoxia-mediated induction of GLUT-1 expression. Significant reduces of the HIF1 protein level | 50 µM | 24 h | S2–013 and CD18 cells | - | [186] | |
| Translocates FOXO1 and reduces G6Pc-mRNA levels | 20–100 µM | 24 h | U-2 OS cells | - | [192] | |
| Luteolin | Inhibits colon carcinogenesis through iNOS and COX-2 | 1.2 mg/body kg | 3 weeks | Balb/c mice | - | [190] |
| Regulates HIF1α -VEGF/MMP9signaling pathway through suppression of HIF1α activation | 20 µM | 24 h | RAW264.7 cells | - | [184] | |
| Induces cell apoptosis through antioxidant enzymes activity like SODs e CAT | 50 µM | 6 to 24 h | CH27 cells | - | [201] | |
| Translocates FOXO1 and reduces G6Pc-mRNA levels | 10–100 µM | 24 h | U-2 OS cells | - | [192] | |
| Increases NRF2, increases GSH content, decreases the level of GSSG, and prevents tumorigenesis | 1–20 µM | 8–24 h | Caco-2, HT-29, HepG2, and HEK-293 cells | - | [160] | |
| Kaempferol | Blocks ROS generation causing cell cycle arrest at G1 and G2/M arrest involving p53 and p38 | 50 and 100 μM | 24, 48, and 72 h | HCT116 cells | - | [195] |
| Reduces the COX-2, p-AKT, and p-ERK levels, decreasing tumor growth | 60 or 120 µM | 48 h | MKN28 and SGC7901 cells | - | [197] | |
| Causes HIF1α mislocalization into the cytoplasm due to p44/42 MAPK inactivation, resulting in the suppression of HIF1 activity | 5 to 50 µM | 4 h | Huh7 cells | - | [185] | |
| Triggers ROS generation and apoptosis through reduction of the thioredoxin and SOD concentrations | 50 µM | 72 and 96 h | LN229, U87MG, and T98G cells | - | [194] | |
| Myricetin | Activates NRF2 by modifying the KEAP1 protein, decreasing NRF2 ubiquitination, and increasing HO-1 levels | 20 µM | 24 h | HepG2 cells | - | [174] |
| Quercetin | Interacts directly with NRF2 increasing its half-life | 0–40 µM | 6 to 12 h | HepG2 cells | - | [169] |
| Inhibits metastasis of cancer cells by blocking AKT/mTOR/c-Myc signaling pathway | 40 µM | 24 h | A431-III cells | - | [202] | |
| Suppresses the mobility of breast cancer by suppressing glycolysis through AKT-mTOR-PKM2 | 50 mg/kg twice daily | 1 month | BALB/c nude mice | - | [149] | |
| Modulates the balance between HIF1α translation and degradation, inhibiting HIF1α protein synthesis and accumulation | 100 µM | 8 h | LNCaP, SkBr3, and CX-1 cells | - | [179] | |
| Daidzein | Increases expression of QR mRNA and its activity, as well as increased NRF2/ARE binding capacity | 1 and 5 µM | 24 and 48 h | Hepa-1c1c7 cells | - | [191] |
| Genistein | Increases expression of QR mRNA and its activity, as well as increased NRF2/ARE binding capacity | 1, 5, and 25 µM | 24 and 48 h | Hepa-1c1c7 cells | - | [191] |
| Reduces the level of methylation in the KEAP1 promoter region, leading to an increased mRNA expression, thus effectively inhibited the transcription of NRF2 to the nucleus | 10 µM | 48 h | A549 cells | - | [177] | |
| Sensitizes aerobic glycolytic cells to apoptosis by directly downregulating HIF1α, inactivating GLUT1 and HK2 to suppress aerobic glycolysis | 20–80 mg/kg | 21 days | Athymic BALB/C nu/nu mice | - | [187] | |
| Vitexin | Suppresses melanoma cell growth through DNA damage by increasing ROS levels | 40 or 80 mg/kg | 2–3 weeks | c BALB/c female nude mice (nu/nu) | - | [178] |
| 5–20 µM | 24 h | A375, Sk-Mel-5, and Sk-Mel-28 cells | - | |||
| Decreases ROS levels, increases GSH and SOD levels | 10 and 100 µM | 24 h | PC12 cells. | - | [200] | |
| Taxifolin | Inhibits NF-κB-mediated Wnt/β-catenin signaling, via up-regulation of NRF2 pathway | 4 μg/kg | 15 to 30 weeks | Albino Swiss mice | - | [66] |
| Cell Growth Signaling | |||||||
|---|---|---|---|---|---|---|---|
| Molecular Mechanisms | Concentration | Incubation | Model | Observations | References | ||
| Cyanidin | Targets PDK1 to suppress PI3K/AKT signaling, decreasing p-AKT, p-mTOR, and PDK1 activity | 40 µM | 24 h | HepG2, Hep3B, HepG2/DDP, and Hep3B/DDP cells. HepG2 xenograft BALB/c mice | Reverses oxaliplatin resistance | [252] | |
| Cyanidin-3-o-glucoside | Inhibits the growth of MDA-MB-231 cells in vivo. Inhibits EGFR/AKT signaling, and promotes EGFR degradation | 150 μM and 420 mg/kg | 24 h and 6 weeks | MCF-7, SK-BR-3, MDA-MB-436, BT-20, and MDA-MB-231 cells and MDA-MB-231-luc xenograft mice | - | [253] | |
| Increases p-ERK1/2 and p-p38 MAPK, and decreases p-AKT | 5, 50 and 100 µM | 48 h | Meg-01 cells | - | [254] | ||
| Delphinidin | Inhibits proliferation and decreases p-AKT, p-mTOR, p-S6K1, and p-eIF4E | 20, 40, and 80 μM | 48 h | MDA-MB-453 and BT474 cells | Increases p-AMPK at 80 μM | [260] | |
| Prevents HIF1α expression and ERK1/2, AKT, mTOR, and S6K1 phosphorylation | 10, 20, and 40 μM | 1h | A549 cells | Induced by cobalt chloride (200 μM) and EGF (20 ng/mL) treatments | [261] | ||
| Decreases p-PI3K, p-AKT (Ser473), and p-ERK1/2 | 20, 40, and 60 μM in cell culture. 1 or 2mg in xenograft BALB/c mice | 48 h | NCI-H441 and SK-MES-1 cells and NCI-H441 xenograft mice | Decreased tumor volume in the NCI-H441 xenograft model | [212] | ||
| Decreased p-AKT, p-S6K1, and p-ERK | 0.1, 1 and 10 μM | 24 h | ES2 cells | Delphinidin exhibits the same effects as cisplatin and paclitaxel | [262] | ||
| Decreased cell proliferation in a dose-dependent manner and decreased p-ERK1/2, p-S6K1, p-S6, and p-AKT | 0.1, 1 and 10 μM | 30 min | SKOV3 cells | Sensitizes to paclitaxel treatment | [263] | ||
| Epigallocatechin gallate (EGCG) | Decreases p-mTOR and p-AKT and increased PTEN | 40 µg/ml | 48 h | PANC-1 and BxPC-3 cells | - | [215] | |
| Decreases p-IGF1R, p-ERK1/2, and p-AKT in SW837 cells. | 25 μg/mL in cell culture and tap water containing 0.1% or 1% EGCG in xenograft mice model. | 3,6,12 and 24 h in cell culture and 35 days in xenograft mice model. | SW837 cell line and xenograft SW837 BALB/c nude mice. | Decreases tumor volume while decreases p-AKT and p-ERK1/2 at 0.1 and 1% of EGCG in xenograft BALB/c mice | [250] | ||
| Inhibits the proliferation in a dose-dependent manner. Decreases p-AKT and p-mTOR | 10, 20, 40, and 80 μM | 24, 48, and 72 h | PANC-1 cells | - | [272] | ||
| Prevents AKT phosphorylation (Ser473) induced by IGF-1 treatment in A549, MDA-MB-231, and LnCAP cells, but not in PC-3 | 25 μM | 1 h | MDA-MB-231, PC-3, LnCAP, and A549 cells | Decreases MDA-MB-231 and A549 cell proliferation in a dose-dependent manner after 72 h | [251] | ||
| Decreases HuH7 cell proliferation in a dose-dependent manner. Decreases VEGF secretion. Decreases tumor volume and p-VEGF, p-ERK1/2 and p-AKT | 0–100 µg/mL in cell culture/tap water containing 0.01 or 0.1% EGCG in mice xenograft model | 24 or 48 h/5 weeks | HuH7 cells and mice BALB/c xenograft model | - | [273] | ||
| Eriodictyol | Decreases p-PI3K, p-mTOR, and p-AKT | 25, 50 and 100 μM | 48 h | A549 and FR2 cells | - | [209] | |
| Decreases p-PI3K and p-AKT | 25, 50 and 100 μM | 48 h | U87-MG and CHG-5 cells | - | [62] | ||
| Naringenin | Decreases p-AKT (Ser473), p-MEK1/2, and p-MAPK after insulin stimuli. Inhibits glucose uptake | 100 μM | 15 min | T47D and MCF-7 cells | - | [248] | |
| Decreases p-AKT | 200 μM | 24 h | THP-1 cells | - | [274] | ||
| Decreases p-AKT | 20, 40, or 80 μM | 24, 48 and 72 h | SGC7901 cells | - | [275] | ||
| Decreases p-AKT | 40 µM | 48 h | SGC7901 cells | Combined with 5 µM ABT-737 | [249] | ||
| In JAR cells, increases p-ERK1/2, p-AKT, and p-S6K1, but decreases p-S6. In JEG cells, increases p-ERK1/2, p-S6K1, but decreases p-S6 and p-AKT | 12.5–100 μM | 48 h | JAR and JEG-3 cells | - | [276] | ||
| Apigenin | Prevents AKT phophorylation | 40 μM | 2 h | MDA-MB-231 cells | Induced by Hepatocyte growth factor (40 ng/mL) | [243] | |
| Prevents p-AKT (Ser473) and p-GSK3b (Ser9) during hypoxia. Inhibits HIF1α activity and decreases VEGF mRNA levels during hypoxia | 25, 50 and 100 μM | 1 h | PC3–M cells | - | [277] | ||
| Increases p-AKT (Ser473), p-ERK1, and p-ERK2 in dorsolateral prostate of TRAMP mice decreases IGF-I and increases IGFBP-3 in the serum and the dorsolateral prostate | 20 μg and 50 μg/day | 20 weeks | C57BL/TGN TRAMP mice | Apigenin also inhibits tumor growth and metastasis | [278] | ||
| Increases p-AMPK (Thr172) and decreases p-AKT (Ser473), LKB1, p-S6K1 (Thr389), p-4EBP1 and 4EBP1 | 20 μM | 24 h | HaCaT cells | - | [225] | ||
| Decreases p-AKT, p-mTOR and p-ERK1/2 | 100 μM | 24 h | A375 and C8161cells | - | [244] | ||
| Luteolin | Decreases p-AKT (Ser473), p-S6K1, and p-S6. | 10, 25, and 50 μM | 48 h | T24 and 5637 cells | Decreases tumor growth in the xenograft model | [245] | |
| Decreases p-AKT, p-mTOR, p-S6K1 and increases p-ERK1/2. Decreases expression of SREBP1, SREBP2, and SREBP cleavage-activating protein (SCAP) mRNAs and protein expression | 5, 10 and 20 μM | 48 h | JAR and JEG-3 cells | Exhibits synergistic effects with etoposide, cisplatin, and paclitaxel | [228] | ||
| Decreases p-AKT, p-S6, and p-4EBP1 in NCI–H1975 cells | 10, 30, 50, 80 and 100 μM | 24 h | A549, HCC827, and NCl-H1975 cells | - | [226] | ||
| Decreases p-IGFR, p-AKT, and p-mTOR | 5, 10 and 20 μM | 24 h | U251MG and U87MG cells | - | [279] | ||
| Vitexin | Decreases p-AKT, p-mTOR and p-PI3K | 20 μM | 48 h | A549 cells | - | [207] | |
| Increases p-AMPK and decreases p-PI3K, p-AKT, and p-mTOR | 20 and 40 μM | 24 h | ACHN and OS-RC-2cells | - | [206] | ||
| Myricetin | Decreases p-AKT (Ser473) and p-S6K1 | 5–20 μM | 24 h | A2780/CP70 and OVCAR-3 cells | - | [241] | |
| Increases p-AKT, p-S6K1, p-S6, and p-ERK1/2 | 20, 50 and 100 μM | D-17 canine osteosarcoma cells | - | [270] | |||
| Increases p-AKT, p-ERK1/2 and p-p90RSK | 5, 10 and 20 μM | JAR and JEG-3 cells | Exhibits synergistic antiproliferative effects with cisplatin and etoposide | [271] | |||
| Decreases AKT, p-AKT (Ser473), and EGFR expression. 200 uM decreases K-ras and Raf-1, ERK and p-ERK | 25–200 μM | 24 h | DBTRG cells | - | [242] | ||
| Decreases p-ERK and ERK, AP-1 and MEK1 activity | 5–20 μM | 1 h | JB6 P+ and H-Ras-transformed JB6 P+ cells | Treatment combined with 12-O-tetradecanoylphorbol-13-acetate (TPA) 20 ng/mL) | [135] | ||
| Kaempferol | Increases PTEN while decreases p-PI3K, p-AKT, p-mTOR and p-S6K1 | 50 μM | 24 h | HepG2 cells | - | [213] | |
| Increases p53 and PTEN while decreases PI3K and AKT mRNA expression | 25, 50 and 100 μM | 24, 48, and 72 h | HeLa cells | - | [214] | ||
| Increases PTEN and decreases p-AKT | 40 μM | 48h | EJ cells | - | [236] | ||
| Decreases p-AKT (Ser473 and Thr308). | 50 μM | 48 and 72 h | K562 and U937 cells | - | [235] | ||
| Decreases p-AKT (Ser473) | 25–100 μM | 24 h | RCC 786-O cells | - | [237] | ||
| Inhibits p-AKT (Ser473) and increases p-MEK2 and p-MAPK | 35–70 μM | 48 h | A549 cells | - | [238] | ||
| Quercetin | Decreases the expression of SREBPs and nuclear chSREBP | 25 mM | 4 h | C6 cells | - | [229] | |
| Decreases p-AKT and p-mTOR | 25 and 50 μM | 24 h | SMMG-7721 and BEL-7402 cells | - | [232] | ||
| Suppresses cell invasion and migration of breast cancer while decreases p-mTOR, p-AKT, and p-S6K1. Decreases tumor growth in xenograft model though decreases of VEGF+ cells, p-AKT, and PKM2 | 30 µM in cell culture. 50 mg/kg in xenograft model | 24 h/4 weeks | MCF7 and MDA-MB-231 cells and MCF7 xenograft BALB/C mice | - | [149] | ||
| Decreases tumor growth | 15 and 45mg/kg | 13 weeks | GFP-MDA-MB-231 xenograft SCID mice | - | [224] | ||
| Decreases p-4EBP1, p-S6K1, p-AKT | 15 μM | 15 min | MDA-MB-231and MDA-MB-435 cells | - | [224] | ||
| Increases p-AMPK, Sestrin2, p53 and decreases p-mTOR | 25 and 50 μM | 6 h | HCT116 cells | - | [280] | ||
| Quercetin-6-C-β-d-glucopyranoside | Suppresses proliferation through decreases of p-AKT and p-mTOR | 60 μM (PC-3) and 100 μM (DU-145) | 24 and 48 h | PC-3 and DU-145 cells | - | [233] | |
| Daidzein | Decreases p-PI3K, p-AKT, and p-mTOR | 10 µM CC and 50 µM Dadizein | 48 h | MCF-7 and MDA MB-231 cells | Treatments separately or together | [210] | |
| Decreases p-PI3K, p-AKT, and p-GSK3β | 10 and 50 µM | 24 h | SKOV-3, A2780CP, and OVCAR-3 cells | - | [211] | ||
| Anti-proliferative activity against MCF-7 cells and in MDA-MB-231. Inhibits proliferation and invasion | MCF-7 cells at 66.98 ± 4.87 μM and MDA-MB-231 at 93.75 ± 5.15 μM | MCF-7 and MDA-MB-231 cells | - | [240] | |||
| Genistein | Decreases p-AKT and p-mTOR | 40 µM | 12 h | human colon cancer HT-29 | Treatment combined with indol-3-carbinol | [255] | |
| Enhances the antitumor activity of cisplatin and reduces p-mTOR, p-p70S6K1, p-4E-BP1, and p-AKT | 25 μM | 24 h | HeLa cells. | Sensitizes cells to cisplatin treatment. | [256] | ||
| Decreases cell viability, p-PI3K, and p-AKT | 160 μM | 12 and 48 h | Human Bladder Cancer T24 Cells | - | [257] | ||
| Decreases p-EGFR and p-AKT (Ser473) and increases p-ERK1/2 | 50-200 µM | 24 h | The HuCCA-1 and RMCCA-1 human intrahepatic CCA cell lines | - | [258] | ||
| Decreases cell viability, p-AKT, and p-HIF1α | 50 µM | 48 h | A549 human lung carcinoma cells | - | [259] | ||
| S-Equol (a metabolite of Daidzein) | Decreases p-AKT, p-ERK, and p-FOXO3a in vitro. Inhibits the growth of PC-3 xenograft tumors in BALB/c nude mice | 100, 150, and 200 µM | 24 h | LnCaP, DU-145, and PC-3 cells | - | [269] | |
| Taxifolin | Decreases p-PI3K, cell viability and tumor growth | 25, 50 and 100 μM in cells/1mg/kg in mice | 24 h/25 days | A549 and H1975 cells and A549 Xenograft BALB/c nude nude | - | [208] | |
| Taxifolin binds to EGFR and PI3K and decreases its activity. Decreases p-AKT (Thr308 and Ser473), p-S6K1, and p-ERK1/2 | 20, 40, and 80 μM | 24 h | JB6 P+ cells | - | [216] | ||
| Decreases p-AKT (Ser473) in cell lines and tumor growth in a xenograft model | 25 and 50 μM | 48 h | U2OS and Saos-2 cells and U2OS xenograft BALB/c nude mice | - | [246] | ||
| Autophagy | ||||||
|---|---|---|---|---|---|---|
| Molecular Mechanisms | Concentration | Incubation | Model | Observations | References | |
| Delphinidin | Increases LC3-II, Atg5/12, and p-ULK | 80 µM | 24 h | HER-2 (breast) | - | [260] |
| Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 100 µM | 24 h | U20S cells | - | [320] | |
| Increases LC3-II | 20 µM | 48 h | A549 cells | - | [321] | |
| Epigallocatechin | Increases Beclin1, LC3-II, p62, and Atg5/12/16 | 250 µg/mL | 24 h | A375 cells | - | [322] |
| Increases LC3-II | 12.5 µM | 24 h | HTC-116 cells | - | [323] | |
| Increases LC3-II | 150 µM | 24 h | A549 cells | Sensibilization to γ-ray irradiation | [324] | |
| Increases LC3-II | 100 µM | 48 h | HT-29 and DLD-1 cells | - | [325] | |
| Increases LC3-II, Beclin1, and Atg5/7/12 | 100 µM | 24 h | CAR cells | - | [326] | |
| Naringin | Increases Beclin1 and LC3-II | 2 mM | 24 h | AGS cells | - | [319] |
| Apigenin | Increases Beclin1 and LC3-II | 25, 50, 100, and 200 µM | 36 h | U251, U118, and U87 cells | - | [312] |
| Increases Beclin1 and LC3-II | 80 µM | 12 h | HepG2 cells | - | [313] | |
| Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 90 µM | 24 h | HepG2 cells | - | [314] | |
| Stimulates the formation of autophagosomes (decreases in p62 and increase in LC3-II), increase in Beclin1, and Atg5 | 10, 20, and 40 µM | 24 h | HepG2 | - | [315] | |
| Luteolin | Increases Beclin1 and LC3-II | 80 µM | 24 h | HepG2 cells | - | [316] |
| Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 100 µM | 48 h | Hep 3B cells | - | [317] | |
| Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 20 µM | 24 h | MET4 cells | - | [318] | |
| Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 100 µM | 24 h | MET4 cells | - | [311] | |
| Kaempferol | Increases Beclin1, LC3-II, and in Atg5, 7, 12 | 75 µM | 24 h | HepG2 cells | - | [305] |
| Inhibits autophagosome formation (accumulation of LC3-I and p62). Reduces Beclin1 | 20 µM | 48 h | SKVCR cells | - | [306] | |
| Increases Beclin1. Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II). Increases Atg5 | 25 µM | 24 h | HT-29 cells | - | [307] | |
| Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II). Increases Atg5, 7 | 100 µM | 24 h | HepG2 e Hh7 cells | - | [308] | |
| Myricetin | Increases Beclin1. Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 60 µM | 24 h | MCF-7/MDA-MB-231cells | - | [309] |
| Increases Beclin1 and LC3-II | 100 µM | 24 h | HCT116 and SW620 cells | - | [310] | |
| Increases Beclin1. Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 100 µM | 24 h | SK-MEL-28 cells | - | [311] | |
| Quercetin | Increases Beclin1. Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 40 µM | 24 h | HepG2 cells | - | [303] |
| Increases Beclin1 and LC3-II | 100 µM | 24 h | PC-3 cells | - | [304] | |
| Increases Beclin1 and LC3-II | 30 µM | 24 h | MCF-7 and MDA-MB 231 cells | - | [149] | |
| Genistein | Increases LC3 | 100 µM | 72 h | MCF-7 cells | - | [327] |
| Increases Beclin1 | 500 ppm | 55 days | Xenografts of breast tumors | - | [328] | |
| Increases Beclin 1. Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 60 µM | 24 h | A549 cells | - | [329] | |
| Increases LC3-II and Beclin1 | 100 µM | 24 h | MIA-PaCa 2 cells | - | [330] | |
| Increases Beclin 1 and p62 | 40 µM | 24 h | A549 cells | - | [331] | |
| Quercetin | Increases Atg5. Stimulates the formation of autophagosomes (decreases p62 and increases LC3-II) | 10, 20, and 40 µM | 24 h | MDA-MB-231 cells | - | [302] |
| Apigenin (Apigenin-7-methyl ether) | Stimulates the formation of autophagosomes (decreases in p62 and increase in LC3-II), increases in Beclin1, and Atg5 | 20, 40, and 80 µM | 24 h | MDA-MB-231 cells | - | |
| Apoptosis | ||||||
|---|---|---|---|---|---|---|
| Molecular Mechanisms | Concentration | Incubation | Model | Observations | References | |
| Cyanidin | Up-regulates cleaved Caspase 3 | 50–100 µg/mL | 24 h | Jurkat cells | - | [368] |
| Up-regulates p53 and caspase 3 | 50–300 µg/mL | 24 h | HeLa cells | - | [342] | |
| Up-regulates p53 and caspase 3 | 100–400 µg/mL | 24 h | HeLa cells | - | ||
| Epigallocatechin | Down-regulates Mcl-1, c-FLIP, and Bcl-2 | 50 µg/mL | 48 h | Renal Cell Carcinoma (786-O) | The treatment was combined with 200 ng/mL of TRAIL | [351] |
| Eriodictyol | Down-regulates Bcl-XL | 25–100 µM | 48 h | U87MG and CHG-5 cells | - | [62] |
| Up-regulates BAX/Bcl-2 ratio | 50–100 µM | 24/48 h | A549 cells | - | [209] | |
| Naringenin | Down-regulates Bcl-2 | 150–200 µM | 24 h | HepG2 cells | - | [345] |
| Up-regulates p53, Caspase 3 and BAX | 150–200 µM | 24 h | HepG2 cells | - | ||
| Apigenin | Down-regulates Mcl-1 and c-FLIP | 50 and 100 µM | 8 h | Human leukemic T cell line Jurkat HTLV-1-associated ATL cells | - | [354] |
| Up-regulates p53 | 50 and 100 µM | 16 h | ATL cells | - | ||
| Down-regulates Bcl-2 | 20 µM | 24 h | PC-3 cells | - | [347] | |
| Up-regulates BAX | 20 µM | 24 h | PC-3 cells | - | ||
| Up-regulates PARP cleaved | 50 µg/day | 8 weeks | Athymic nude mice | - | ||
| Luteolin | Up-regulates BAX and Caspase 3 | 10–30 µM | 48 h | MDA-MB-231 cells | - | [344] |
| Down-regulates Bcl-2 | 10–30 µM | 48 h | MDA-MB-231 cells | - | ||
| Kaempferol | Increases cleaved caspase 3 | 10 µM | 72 h | Miapaca-2 cells | Tre treatment was compared with Gefetinib action | [341] |
| Increases cleaved caspase 3 | 100 µM | 72 h | Panc-1 cells | Tre treatment was compared with Gefetinib action | ||
| Myricetin | Down-regulates Bcl-2 | 50 and 100 µM | 24 h | HCT-15 cells | - | [346] |
| Up-regulates BAX | 50 and 100 µM | 24 h | HCT-15 cells | - | ||
| Vitexin | Up-regulates cleaved Caspase-3 and Caspase-9 | 10–40 µM | 24 h | ACHN and OS-RC-2 cells | The treatment was compared with Metformin and Rapamycin action | [206] |
| Down-regulates Bcl-2 and up-regulates BAX and cleaved Caspase-3 and Caspase-9 | 10–40 µM | 48 h | A549 cells | - | [207] | |
| Quercetin | Down-regulates c-FLIP | 200 µM | 12 h | HepG2 cells | - | [369] |
| Down-regulates c-FLIP | 50, 100 and 200 µM | 12 h | SNU-475 cells | - | ||
| Up-regulates cleaved PARP and p53 | 20 µM | 24 h | Leukemia cell (Nalm6) | - | [353] | |
| Down-regulates Mcl-1 | 20 µM | 24 h | Leukemia cell (Nalm6) | - | ||
| Up-regulates BAX, Caspase 3 and p53 | 80 µM | 24 h | HeLa cells | - | [55] | |
| Daidzein | Down-regulates Bcl-2 | 90 µM | 24 h | BEL-7402 cells | - | [352] |
| Up-regulates cleaved PARP | 25–100 µM | 48 h | Choriocarcinoma cell line (JAR) | - | [349] | |
| Down-regulates Bcl-2/BAX ratio | 25–100 µM | 48 h | Choriocarcinoma cell line (JEG-3) | - | ||
| Genistein | Up-regulates Caspase 3 | 30–70 µM | 48 h | HT29 cells | - | [343] |
| Up-regulates BAX | 20–50 µM | 48 h | HL-60 cells | - | [348] | |
| Down-regulates Bcl-2 | 40–50 µM | 48 h | HL-60 cells | - | ||
| Up-regulates BAX and BAK | 0.2–0.4 mg/kg | 23 days | Athymic nude mice | - | ||
| Taxifolin | Up-regulates BAK | 40–60 µM | 24 h | HCT116 cells | - | [350] |
| Down-regulates Bcl-2, Bcl-xL, and Bid | 40–60 µM | 24 h | HCT116 cells | - | ||
| Cell Cycle | ||||||
| Molecular mechanisms | Concentration | Incubation | Model | Observations | References | |
| Anthocyanidins | Up-regulates p21 | 10–100 µg/mL | 24 h | Jurkat cells | - | [368] |
| Down-Downregulates Cyclin D1 | 400 µg/mL | 24 h | HeLa cells | - | [342] | |
| Down-regulates Cyclin D1 | 50–300 µg/mL | 24 h | HeLa cells | - | ||
| Luteolin | Down-regulates Cyclin D1 | 10–30 µM | 24 h | MDA-MB-231 cells | - | [344] |
| Up-regulates p21 | 10–30 µM | 24 h | MDA-MB-231 cells | - | ||
| Kaempferol | Down-regulates Cyclin D1, Cyclin A, and Cyclin E | 60 µmol/L | 6 h | HT-29 cells | - | [196] |
| Quercetin | Up-regulates p21 | 80 µM | 24 h | HeLa cells | - | [55] |
| Daidzein | Down-regulates cyclin D, CDK2, CDK1, and CDK4 | 100 µM | 72 h | MDA-MB-453 and MCF-7 cells | - | [370] |
| Genistein | Up-regulates p53 and p21 | 50–100 µM | 72 h | HCT116 cells | - | [358] |
| Taxifolin | Up-regulates p21 | 40–60 µM | 24 h | HCT116 cells | - | [350] |
| Down-regulates Cdk-2, Cdk-4, Cdk-6, Cyclin D, Cyclin A, Cyclin B | 40–60 µM | 24 h | HCT116 cells | - | ||
| DNA Repair | ||||||
| Molecular mechanisms | Concentration | Incubation | Model | Observations | References | |
| Catechin | Down-regulates CHK1 | 5–10 µM | 30 min | MDA-MB-231 cells | - | [360] |
| Epigallocatechin | Down-regulates CHK1/2 | 10–50 µM | ||||
| Naringenin | Down-regulates CHK1/2 | 50 µM | ||||
| Kaempferol | Down-regulates CHK1/2 | 50 µM | ||||
| Genistein | Down-regulates CHK1/2 | 5 µM | ||||
| Senescence | ||||||
| Molecular mechanisms | Concentration | Incubation | Model | Observations | References | |
| Quercetin | Increases p16 | 25 μg/mL | 48 h | Colo-320 and Colo-741 cells | - | [365] |
| Epigallocatechin | Telomerase shortening and increase of β-galactosidase | 10 μg/mL | 98 days | U251 cells | - | [366] |
| Apigenin | Increasesβ-galactosidase | Above 25 μM | 6 days | HCT-15 cells | - | [367] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponte, L.G.S.; Pavan, I.C.B.; Mancini, M.C.S.; da Silva, L.G.S.; Morelli, A.P.; Severino, M.B.; Bezerra, R.M.N.; Simabuco, F.M. The Hallmarks of Flavonoids in Cancer. Molecules 2021, 26, 2029. https://doi.org/10.3390/molecules26072029
Ponte LGS, Pavan ICB, Mancini MCS, da Silva LGS, Morelli AP, Severino MB, Bezerra RMN, Simabuco FM. The Hallmarks of Flavonoids in Cancer. Molecules. 2021; 26(7):2029. https://doi.org/10.3390/molecules26072029
Chicago/Turabian StylePonte, Luis Gustavo Saboia, Isadora Carolina Betim Pavan, Mariana Camargo Silva Mancini, Luiz Guilherme Salvino da Silva, Ana Paula Morelli, Matheus Brandemarte Severino, Rosangela Maria Neves Bezerra, and Fernando Moreira Simabuco. 2021. "The Hallmarks of Flavonoids in Cancer" Molecules 26, no. 7: 2029. https://doi.org/10.3390/molecules26072029
APA StylePonte, L. G. S., Pavan, I. C. B., Mancini, M. C. S., da Silva, L. G. S., Morelli, A. P., Severino, M. B., Bezerra, R. M. N., & Simabuco, F. M. (2021). The Hallmarks of Flavonoids in Cancer. Molecules, 26(7), 2029. https://doi.org/10.3390/molecules26072029