
In Vitro Inhibitory Effects of Viburnum opulus Bark and Flower Extracts on Digestion of Potato Starch and Carbohydrate Hydrolases Activity


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Phenolic Profiles of the Bark and Flower Extracts and Fractions
2.2. In Vitro Hydrolysis of Potato Starch in Presence of V. opulus Bark and Flower Extracts
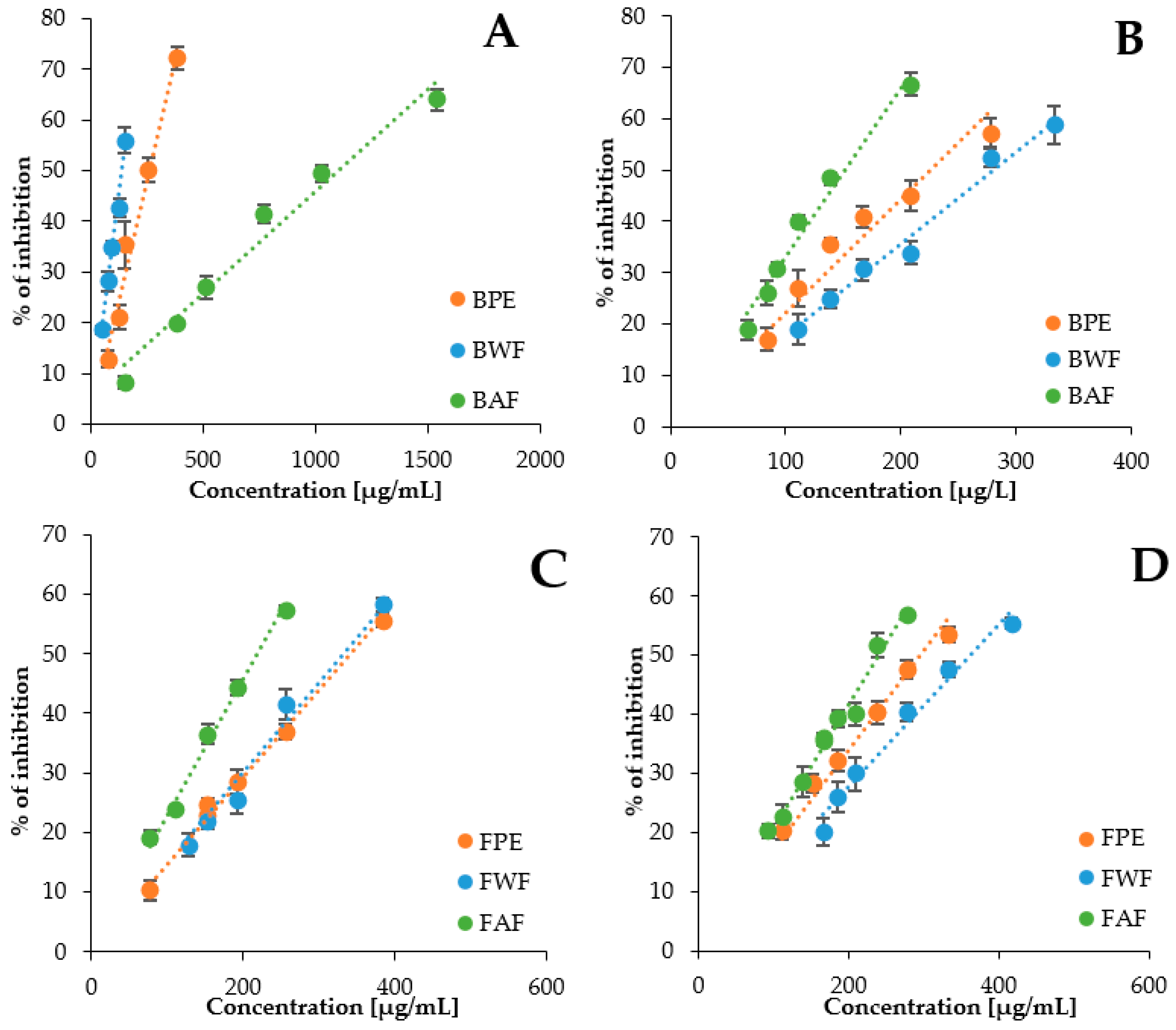
2.3. Effects of V. opulus Phenolic Extracts from Bark and Flower on α-Amylase and α-Glucosidase Activity
2.4. Effects of V. opulus Bark and Flower Phenolic Extracts on α-Amylase and α-Glucosidase Spectra
3. Materials and Methods
3.1. Standards and Reagents
3.2. Plant Material and Phenolic Extracts Preparation
3.3. Identification and Content of Individual Phenolic Compounds
3.4. Total Proanthocyanidins Content
3.5. Simulated In Vitro Digestion of Potato Starch
3.6. α-Amylase Inhibition Assay
3.7. α-Glucosidase Inhibition Assay
3.8. Fluorescence Measurements
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Zhang, Y.; Pan, X.F.; Chen, J.; Xia, L.; Cao, A.; Zhang, Y.; Wang, J.; Li, H.; Yang, K.; Guo, K.; et al. Combined lifestyle factors and risk of incident type 2 diabetes and prognosis among individuals with type 2 diabetes: A systematic review and meta-analysis of prospective cohort studies. Diabetologia 2020, 63, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankatillake, C.; Huynh, T.; Dias, D.A. Understanding glycemic control and current approaches for screening antidiabetic natural products from evidence-based medicinal plants. Plant Methods 2019, 15, 1–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roheem, F.O.; Soad, S.Z.M.; Ahmed, Q.U.; Shah, S.A.A.; Latip, J.; Zakaria, Z.A. Evaluation of the Enzyme Inhibitory and Antioxidant Activities of Entada spiralis Stem Bark and Isolation of the Active Constituents. Molecules 2019, 24, 1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Noh, S.; Lim, S.; Kim, B. Plant extracts for type 2 diabetes: From traditional medicine to modern drug discovery. Antioxidants 2021, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Vonia, S.; Hartati, R.; Insanu, M. In vitro alpha-glucosidase inhibitory activity and the isolation of luteolin from the flower of Gymnanthemum amygdalinum (Delile) Sch. Bip ex Walp. Molecules 2022, 27, 2132. [Google Scholar] [CrossRef]
- Mahomoodally, M.F.; Jugreet, S.; Sinan, K.I.; Zengin, G.; Ak, G.; Ceylan, R.; Jekő, J.; Cziáky, Z.; Angelini, P.; Flores, G.A.; et al. Pharmacological potential and chemical characterization of Bridelia ferruginea Benth.—A native tropical african medicinal plant. Antibiotics 2021, 10, 223. [Google Scholar] [CrossRef]
- Alam, F.; Shafique, Z.; Amjad, S.T.; Bin Asad, M.H.H. Enzymes inhibitors from natural sources with antidiabetic activity: A review. Phytother. Res. 2019, 33, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Quan, N.V.; Xuan, T.D.; Tran, H.-D.; Thuy, N.T.D.; Trang, L.T.; Huong, C.T.; Andriana, Y.; Tuyen, P.T. Antioxidant, α-amylase and α-glucosidase inhibitory activities and potential constituents of Canarium tramdenum bark. Molecules 2019, 24, 605. [Google Scholar] [CrossRef] [Green Version]
- Aba, P.E.; Asuzu, I.U. Mechanisms of actions of some bioactive anti-diabetic principles from phytochemicals of medicinal plants: A review. Indian J. Nat. Prod. Resour. 2018, 9, 85–96. [Google Scholar]
- Chusak, C.; Henry, C.J.; Chantarasinlapin, P.; Techasukthavorn, V.; Adisakwattana, S. Influence of Clitoria ternatea flower extract on the in vitro enzymatic digestibility of starch and its application in bread. Foods 2018, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Dall’Acqua, S.; Sinan, K.I.; Ferrarese, I.; Sut, S.; Bene, K.; Mahomoodally, M.F.; Sadeer, N.B.; Ak, G.; Zengin, G. Chromatographic separation of Breynia retusa (Dennst.) alston bark, fruit and leaf constituents from bioactive extracts. Molecules 2020, 25, 5537. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Luo, J.; Chai, Y.; Guo, Y.; Tianzhi, Y.; Bao, Y. Hypoglycemic effect of Taraxacum officinale root extract and its synergism with Radix Astragali extract. Food Sci. Nutr. 2021, 9, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Alkhalidy, H.; Liu, D. The Emerging role of polyphenols in the management of type 2 diabetes. Molecules 2021, 26, 703. [Google Scholar] [CrossRef]
- Prpa, E.J.; Bajka, B.H.; Ellis, P.R.; Butterworth, P.J.; Corpe, C.P.; Hall, W.L. A systematic review of in vitro studies evaluating the inhibitory effects of polyphenol-rich fruit extracts on carbohydrate digestive enzymes activity: A focus on culinary fruits consumed in Europe. Crit. Rev. Food Sci. Nutr. 2020, 61, 3783–3803. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.R.; Alves, M.G.; Casal, S.; Oliveira, P.F.; Silva, B.M. Promising potential of dietary (poly)phenolic compounds in the prevention and treatment of diabetes mellitus. Curr. Med. Chem. 2017, 24, 334–354. [Google Scholar] [PubMed]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef] [PubMed]
- Vichayanrat, A.; Ploybutr, S.; Tunlakit, M.; Watanakejorn, P. Efficacy and safety of voglibose in comparison with acarbose in type 2 diabetic patients. Diabetes Res. Clin. Pract. 2002, 55, 99–103. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Pawlik, N.; Polka, D.; Nowak, A.; Koziołkiewicz, M.; Podsędek, A. Viburnum opulus fruit phenolic compounds as cytoprotective agents able to decrease free fatty acids and glucose uptake by Caco-2 cells. Antioxidants 2019, 8, 262. [Google Scholar] [CrossRef] [Green Version]
- Zakłos-Szyda, M.; Kowalska-Baron, A.; Pietrzyk, N.; Drzazga, A.; Podsędek, A. Evaluation of Viburnum opulus L. fruit phenolics cytoprotective potential on insulinoma MIN6 cells relevant for diabetes mellitus and obesity. Antioxidants 2020, 9, 433. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Majewska, I.; Redzynia, M.; Koziołkiewicz, M. Antidiabetic effect of polyphenolic extracts from selected edible plants as α-amylase, α-glucosidase and PTP1B inhibitors, and β-pancreatic cells cytoprotective agents—A comparative study. Curr. Top. Med. Chem. 2015, 15, 2431–2444. [Google Scholar] [CrossRef] [PubMed]
- Kajszczak, D.; Kowalska-Baron, A.; Podsędek, A. Glycoside hydrolases and non-enzymatic glycation inhibitory potential of Viburnum opulus L. fruit—In vitro studies. Antioxidants 2021, 10, 989. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Lee, H.; Han, J.; Lee, H.; Kattia, R.O.; Nelson, Z.V.; Choi, S.; Kim, S.-Y.; Park, H.-Y.; Jeong, H.G.; et al. Viburnum stellato-tomentosum extract suppresses obesity and hyperglycemia through regulation of lipid metabolism in high-fat diet-fed mice. Molecules 2021, 26, 1052. [Google Scholar] [CrossRef] [PubMed]
- Mazur, M.; Szperlik, J.; Salejda, A.M.; Krasnowska, G.; Kolniak-Ostek, J.; Bąbelewski, P. Description of the guelder rose fruit in terms of chemical composition, antioxidant capacity and phenolic compounds. Appl. Sci. 2021, 11, 9221. [Google Scholar] [CrossRef]
- Kajszczak, D.; Zakłos-Szyda, M.; Podsędek, A. Viburnum opulus L.—A review of phytochemistry and biological effects. Nutrients 2020, 12, 3398. [Google Scholar] [CrossRef] [PubMed]
- Bubulica, V.M.; Anghel, I.; Grumezescu, A.M.; Saviuc, C.; Anghel, G.A.; Chifriuc, M.C.; Gheorghe, I.; Lazar, V.; Popescu, A. In vitro evaluation of bactericidal and antibiofilm activity of Lonicera tatarica and Viburnum opulus plant extracts on Staphylococcus strains. Farmacia 2012, 60, 80–91. [Google Scholar]
- Sharifi-Rad, J.; Quispe, C.; Vergara, C.V.; Kitic, D.; Kostic, M.; Armstrong, L.; Shinwari, Z.K.; Khalil, A.T.; Brdar-Jokanović, M.; Ljevnaić-Mašić, B.; et al. Genus Viburnum: Therapeutic potentialities and agro-food-pharma applications. Oxid. Med. Cell. Longev. 2021, 2021, 3095514. [Google Scholar] [CrossRef]
- Polka, D.; Podsędek, A.; Koziołkiewicz, M. Comparison of chemical composition and antioxidant capacity of fruit, flower and bark of Viburnum opulus. Plant Food Hum. Nutr. 2019, 74, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Skrypnik, L.N.; Kislyakova, L.A.; Maslennikov, P.V.; Feduraev, P.V. Accumulation of phenolic antioxidants in flowers and fruits of guelder rose (Viburnum opulus L.) depending on site conditions. IOP Conf. Ser. Earth Environ. Sci. 2021, 677, 042042. [Google Scholar] [CrossRef]
- Wójcik-Bojek, U.; Rywaniak, J.; Bernat, P.; Podsędek, A.; Kajszczak, D.; Sadowska, B. An in vitro study of the effect of Viburnum opulus extracts on key processes in the development of Staphylococcal infections. Molecules 2021, 26, 1758. [Google Scholar] [CrossRef]
- Turek, S.; Cisowski, W. Free and chemically bonded phenolic acids in barks of Viburnum opulus L. and Sambucus nigra L. Acta Pol. Pharm.-Drug Res. 2007, 64, 377–383. [Google Scholar]
- Shin, S.; Saravanakumar, K.; Sathiyaseelan, A.; Mariadoss, A.V.A.; Park, S.; Park, S.; Han, K.; Wang, M.-H. Phytochemical profile and antidiabetic effect of the bioactive fraction of Cirsium setidens in streptozotocin-induced type 2 diabetic mice. Process Biochem. 2022, 116, 60–71. [Google Scholar] [CrossRef]
- Mariadoss, A.V.A.; Park, S.; Saravanakumar, K.; Sathiyaseelan, A.; Wang, M.H. Ethyl acetate fraction of Helianthus tuberosus L. induces anti-diabetic, and wound-healing activities in insulin-resistant human liver cancer and mouse fibroblast cells. Antioxidants 2021, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Sathiyaseelan, A.; Park, S.; Saravanakumar, K.; Mariadoss, A.V.A.; Wang, M.H. Evaluation of phytochemicals, antioxidants, and antidiabetic efficacy of various solvent fractions of Gynura procumbens (Lour.) Merr. Process Biochem. 2021, 111, 51–62. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Bielicki, P. Polyphenolic composition, antioxidant activity, and polyphenol oxidase (PPO) activity of quince (Cydonia oblonga Miller) varieties. J. Agric. Food Chem. 2013, 61, 2762–2772. [Google Scholar] [CrossRef]
- Dienaitė, L.; Pukalskienė, M.; Pereira, C.V.; Matias, A.A.; Venskutonis, P.R. Valorization of European cranberry bush (Viburnum opulus L.) berry pomace extracts isolated with pressurized ethanol and water by assessing their phytochemical composition, antioxidant, and antiproliferative activities. Foods 2020, 9, 1413. [Google Scholar] [CrossRef]
- Senica, M.; Stampar, F.; Veberic, R.; Mikulic-Petkovsek, M. Fruit seeds of the Rosaceae family: A waste, new life, or a danger to human health? J. Agric. Food Chem. 2017, 65, 10621–10629. [Google Scholar] [CrossRef]
- Bubulica, V.M.; Chirigiu, L.; Grumezescu, A.M.; Popescu, A.; Simionescu, A. Screening of antioxidant potential of Lonicera tatarica, Viburnum opulus and Sambucus ebulus L. by multiple in vitro assays. J. Med. Plant. Res. 2012, 6, 544–552. [Google Scholar]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. HPLC–MSn identification and quantification of flavonol glycosides in 28 wild and cultivated berry species. Food Chem. 2012, 135, 2138–2146. [Google Scholar] [CrossRef]
- Nowicka, P.; Wojdyło, A. Anti-hyperglycemic and anticholinergic effects of natural antioxidant contents in edible flowers. Antioxidants 2019, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Perova, I.B.; Zhogova, A.A.; Cherkashin, A.V.; Éller, K.I.; Ramenskaya, G.V.; Samylina, I.A. Biologically active substances from European guelder berry fruits. Pharm. Chem. J. 2014, 48, 332–339. [Google Scholar] [CrossRef]
- Bujor, O.C.; Le Bourvellec, C.; Volf, I.; Popa, V.I.; Dufour, C. Seasonal variations of the phenolic constituents in bilberry (Vaccinium myrtillus L.) leaves, stems and fruits, and their antioxidant activity. Food Chem. 2016, 213, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Saucier, C.; Mirabel, M.; Daviaud, F.; Longieras, A.; Glories, Y. Rapid fractionation of grape seed proanthocyanidins. J. Agric. Food Chem. 2001, 49, 5732–5735. [Google Scholar] [CrossRef]
- Kato, E. Bioactive compounds in plant materials for the prevention of diabetes and obesity. Biosci. Biotechnol. Biochem. 2019, 83, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Xue, Y.; Lu, X.; Shao, Q.; Cao, Y.; Wu, Z.; Chen, G. The effects of different degrees of procyanidin polymerization on the nutrient absorption and digestive enzyme activity in mice. Molecules 2018, 23, 2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghani, U. Re-exploring promising α-glucosidase inhibitors for potential development into oral anti-diabetic drugs: Finding needle in the haystack. Eur. J. Med. Chem. 2015, 103, 133–162. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Statistics. Available online: https://www.fao.org/faostat/en/#home (accessed on 6 May 2020).
- Zhu, F. Interactions between starch and phenolic compound. Trends Food Sci. Technol. 2015, 43, 129–143. [Google Scholar] [CrossRef]
- Fu, X.; Liu, Q.; Xu, E.; Yang, Y.; Zhu, K.; Jin, Z.; Jiao, A. The effect of Vaccinium bracteatum Thunb. leaves addition on antioxidant capacity, physicochemical properties, and in vitro digestibility of rice extrudates. J. Food Sci. 2021, 86, 4730–4740. [Google Scholar] [CrossRef]
- Yazdankhah, S.; Hojjati, M.; Azizi, M.H. The antidiabetic potential of black mulberry extract-enriched pasta through inhibition of enzymes and glycemic index. Plant Food. Hum. Nutr. 2019, 74, 149–155. [Google Scholar] [CrossRef]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef]
- Kam, A.; Li, K.M.; Razmovski-Naumovski, V.; Nammi, S.; Shi, J.; Chan, K.; Li, G.Q. A comparative study on the inhibitory effects of different parts and chemical constituents of pomegranate on α-amylase and α-glucosidase. Phytother. Res. 2013, 27, 1614–1620. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P. Berry fruits for cancer prevention: Current status and future prospects. J. Agric. Food Chem. 2008, 56, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Narita, Y.; Inouye, K. Inhibitory effects of chlorogenic acids from green coffee beans and cinnamate derivatives on the activity of porcine pancreas α-amylase isozyme I. Food Chem. 2011, 127, 1532–1539. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Li, K.; Liu, Z. Research on the influence of five food-borne polyphenols on in vitro slow starch digestion and the mechanism of action. J. Agric. Food Chem. 2019, 67, 8617–8625. [Google Scholar] [CrossRef]
- Wang, H.; Du, Y.J.; Song, H.C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Abdelli, I.; Benariba, N.; Adjdir, S.; Fekhikher, Z.; Daoud, I.; Terki, M.; Benramdane, H.; Ghalem, S. In silico evaluation of phenolic compounds as inhibitors of A-amylase and A-glucosidase. J. Biomol. Struct. Dyn. 2021, 39, 816–822. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, W.; Sun, W.; Chen, S.; Liu, D.; Kong, X.; Tian, J.; Ye, X. Inhibition of porcine pancreatic α-amylase activity by chlorogenic acid. J. Funct. Foods 2020, 64, 103587. [Google Scholar] [CrossRef]
- Rösch, D.; Bergmann, M.; Knorr, D.; Kroh, L.W. Structure antioxidant efficiency relationships of phenolic compounds and their contribution to the antioxidant activity of sea buckthorn juice. J. Agric. Food Chem. 2003, 51, 4233–4239. [Google Scholar] [CrossRef]
- Bellesia, A.; Verzelloni, E.; Tagliazucchi, D. Pomegranate ellagitannins inhibit α-glucosidase activity in vitro and reduce starch digestibility under simulated gastro-intestinal conditions. Int. J. Food Sci. Nutr. 2015, 66, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.F.; Jayaprakasha, G.K.; Patil, B.S. In vitro bile acid binding capacities of red leaf lettuce and cruciferous vegetables. J. Agric. Food Chem. 2017, 65, 8054–8062. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt (min) | λmax (nm) | [M − H]− (m/z) | MS/MS (m/z) | Phenolic Compound | Extract | Acetate Fraction | Water Fraction |
|---|---|---|---|---|---|---|---|---|
| mg/g of Extract or Fraction | ||||||||
| Flavanols | ||||||||
| 1 | 4.61 | 281 | 577 | 125,161,255 | Procyanidin B1 | 36.56 ± 0.02 c | 22.16 ± 0.06 a | 29.66 ± 1.18 b |
| 2 | 5.11 | 281 | 865 | 407,289,125 | Procyanidin trimer I a,1 | 28.50 ± 0.02 c | 22.86 ± 0.12 b | 14.22 ± 0.02 a |
| 3 | 5.26 | 278 | 289 | 109,159,173 | (+)-Catechin | 64.90 ± 0.03 b | 179.29 ± 0.14 c | 6.70 ± 0.16 a |
| 4 | 5.45 | 279 | 865 | 407,289,125 | Procyanidin trimer II a,1 | 12.75 ± 0.07 c | 10.37 ± 0.02 b | 6.12 ± 0.20 a |
| 5 | 6.12 | 279 | 577 | 125,161,255 | Procyanidin B2 | 10.58 ± 0.06 c | 8.09 ± 0.11 a | 9.50 ± 0.10 b |
| 6 | 6.7 | 279 | 1153 | 287,407,125 | Procyanidin tetramer I a,1 | 4.12 ± 0.02 b | 8.10 ± 0.03 c | 1.92 ± 0.09 a |
| 7 | 6.91 | 279 | 289 | 109,159,173 | (−)-Epicatechin | 7.66 ± 0.01 a | 18.20 ± 0.01 b | - |
| 8 | 7.29 | 279 | 865 | 407,243,289 | Procyanidin C1 | 5.41 ± 0.09 b | 6.85 ± 0.16 c | 2.50 ± 0.27 a |
| 9 | 7.61 | 279 | 577 | 125,161,255 | Procyanidin dimer b,1 | 9.68 ± 0.05 a | 29.16 ± 0.05 b | - |
| Total flavanols | 180.16 ± 0.37 b | 305.08 ± 0.70 c | 70.62 ± 2.02 a | |||||
| Flavalignans | ||||||||
| 10 | 7.39 | 279 | 739 | 177,289,161 | Cinchonain IIx c,2 | 4.28 ± 0.09 b | 8.63 ± 0.01 c | 1.10 ± 0.04 a |
| 11 | 9.23 | 281 | 451 | 176,191,269 | Cinchonain Ix c,2 | 2.28 ± 0.06 a | 3.10 ± 0.01 b | - |
| 12 | 10.88 | 281 | 451 | 189,161 | Cinchonain Ix c,2 | 1.24 ± 0.01 a | 6.78 ± 0.03 b | - |
| Total flavalignans | 7.80 ± 0.16 b | 18.51 ± 0.05 c | 1.10 ± 0.04 a | |||||
| Hydroxycinnamic Acids | ||||||||
| 13 | 4.05 | 324 | 353 | 134,135, 191 | Neochlorogenic acid | 1.16 ± 0.00 b | 0.26 ± 0.00 a | - |
| 14 | 5.71 | 326 | 353 | 191,133 | Chlorogenic acid | 17.15 ± 0.00 a | 22.74 ± 0.01 c | 17.26 ± 0.01 b |
| 15 | 5.9 | 324 | 353 | 191,133 | Cryptochlorogenic acid | 1.07 ± 0.00 b | 0.59 ± 0.00 a | 1.18 ± 0.01 c |
| 16 | 10.79 | 325 | 515 | 191,135 | 3,5-Dicaffeoylquinic acid | 2.06 ± 0.00 c | 0.45 ± 0.00 b | 0.19 ± 0.00 a |
| 17 | 11.63 | 325 | 515 | 191,135 | Dicaffeoylquinic acid d,3 | 3.56 ± 0.01 a | 19.95 ± 0.01 b | - |
| Total hydroxycinnamic acids | 25.00 ± 0.01 b | 43.99 ± 0.02 c | 18.63 ± 0.02 a | |||||
| TOTAL PHENOLICS | 212.96 ± 0.54 b | 367.58 ± 0.77 c | 90.35 ± 2.08 a | |||||
| Peak | Rt (min) | λmax (nm) | [M − H]− (m/z) | MS/MS (m/z) | Phenolic Compound | Extract | Acetate Fraction | Water Fraction |
|---|---|---|---|---|---|---|---|---|
| mg/g of Extract or Fraction | ||||||||
| Flavanon | ||||||||
| 1 | 9.37 | 281 | 449 | 135,151 | Eriodictyol hexoside a,1 | 0.59 ± 0.00 a | 0.71 ± 0.00 b | - |
| Hydroxycinnamic Acids | ||||||||
| 2 | 5.67 | 326 | 353 | 191,133 | Chlorogenic acid | 110.69 ± 0.15 a | 178.47 ± 0.08 c | 124.70 ± 0.06 b |
| 3 | 5.89 | 326 | 353 | 191,133 | Cryptochlorogenic acid | 1.29 ± 0.00 b | 1.33 ± 0.02 c | 0.90 ± 0.00 a |
| 4 | 6.07 | 324 | 179 | 108,134,191 | Caffeic acid | 1.03 ± 0.00 a | 1.13 ± 0.00 b | - |
| 5 | 7.38 | 326 | 335 | 135 | Caffeoylshikimic acid b,2 | 4.89 ± 0.18 a | 28.22 ± 0.01 b | - |
| 6 | 8.29 | 305 | 337 | 191,117,127 | p-Coumaroylquinic acid b,1 | 1.48 ± 0.11 a | 2.80 ± 0.00 b | - |
| 7 | 10.75 | 327 | 515 | 191,135 | 3,5-Dicaffeoylquinic acid | 4.44 ± 0.09 a | 5.60 ± 0.00 b | - |
| Total hydroxycinnamic acids | 123.82 ± 0.53 a | 217.55 ± 0.11 c | 125.60 ± 0.06 b | |||||
| Flavonols | ||||||||
| 8 | 6.34 | 352 | 625 | 299,271 | Quercetin dihexoside c,3 | 6.48 ± 0.01 a | - | 10.27 ± 0.01 b |
| 9 | 6.64 | 338 | 771 | 285 | Quercetin dihexoside c,4 | 1.05 ± 0.03 a | - | 1.87 ± 0.00 b |
| 10 | 7.15 | 322 | 609 | 283,255 | Kaempferol 3-sophoroside d,3 | 69.12 ± 0.40 b | 7.64 ± 0.00 a | 90.67 ± 1.05 c |
| 11 | 7.32 | 345 | 755 | 285 | Quercetin dihexoside c,4 | 6.41 ± 0.01 a | - | 8.77 ± 0.03 b |
| 12 | 7.58 | 352 | 639 | 331,300,270 | Laricitin 3-rutinoside c,3 | 4.06 ± 0.00 b | 0.32 ± 0.00 a | 6.00 ± 0.00 c |
| 13 | 8.08 | 343 | 651 | 283,255 | Quercetin 3-(acetyl)-rutinoside c,4 | 1.71 ± 0.00 a | - | 1.73 ± 0.00 b |
| 14 | 8.19 | 338 | 593 | 283,255,161 | Kaempferol hexoside d,3 | 0.78 ± 0.05 a | - | 1.10 ± 0.00 b |
| 15 | 9.11 | 352 | 595 | 271,255,300 | Quercetin 3-sambubioside c,5 | 1.01 ± 0.00 a | - | 1.51 ± 0.00 b |
| 16 | 9.71 | 352 | 609 | 271,255,300 | Quercetin 3-rutinoside | 7.49 ± 0.01 b | 1.19 ± 0.01 a | 9.53 ± 0.05 c |
| 17 | 9.99 | 352 | 463 | 271,255,243 | Quercetin 3-glucoside | 17.62 ± 0.01 b | 33.67 ± 0.02 c | 8.10 ± 0.00 a |
| 18 | 10.23 | 347 | 579 | 255,227,285 | Quercetin pentosyldeoxyhexoside c,6 | 13.98 ± 0.01 b | 3.59 ± 0.09 a | 19.73 ± 0.00 c |
| 19 | 10.62 | 354 | 505 | 271,255,243 | Quercetin 3-(acetyl)-galactoside c,4 | 5.55 ± 0.01 b | 6.87 ± 0.00 c | 3.32 ± 0.01 a |
| 20 | 10.88 | 347 | 593 | 255,227,285 | Kaempferol 3-rutinoside d,4 | 13.34 ± 0.18 b | 9.45 ± 0.00 a | 15.31 ± 0.00 c |
| 21 | 11.16 | 360 | 447 | 227,255,183 | Kaempferol 3-glucoside | 81.46 ± 0.06 b | 239.18 ± 0.03 c | 42.21 ± 0.05 a |
| 22 | 11.59 | 352 | 477 | 243,271,199 | Isorhamnetin 3-glucoside | 11.65 ± 0.24 b | 23.97 ± 0.00 c | 3.98 ± 0.00 a |
| 23 | 12.05 | 348 | 489 | 227,255 | Kaempferol 3-(acetyl)-glucoside d,4 | 30.74 ± 0.14 b | 64.75 ± 0.04 c | 7.95 ± 0.00 a |
| 24 | 12.28 | 345 | 489 | 227,255 | Kaempferol 3-(acetyl)-glucoside d,4 | 1.28 ± 0.00 a | 2.81 ± 0.00 b | - |
| 25 | 12.53 | 354 | 519 | 243,271,285 | Isorhamnetin 3-(acetyl)-glucoside e,3 | 2.56 ± 0.01 b | 4.60 ± 0.00 c | 0.91 ± 0.00 a |
| 26 | 13.14 | 366 | 285 | 182,117,227 | Kaempferol | 5.29 ± 0.03 | - | - |
| Total flavonols | 281.58 ± 1.20 b | 398.04 ± 0.19 c | 232.97 ± 1.20 a | |||||
| Total phenolics | 405.99 ± 1.73 b | 616.30 ± 0.30 c | 358.57 ± 1.26 a | |||||
| Scheme | Bark | Flower |
|---|---|---|
| Phenolic extract | 71.85 ± 3.50 a | 25.33 ± 1.23 a |
| Acetate fraction | 73.04 ± 3.83 a | 23.27 ± 2.15 a |
| Water fraction | 127.29 ± 1.72 b | 42.53 ± 2.19 b |
| Sample | α-Amylase | α-Glucosidase | |
|---|---|---|---|
| Bark | Phenolic extract | 260.75 ± 2.51 d | 217.03 ± 11.17 c |
| Acetate fraction | 985.80 ± 19.00 f | 164.85 ± 2.75 b | |
| Water fraction | 140.86 ± 4.98 b | 267.05 ± 9.70 e | |
| Flower | Phenolic extract | 351.87 ± 3.02 e | 300.29 ± 12.65 f |
| Acetate fraction | 224.70 ± 3.78 c | 243.08 ± 3.90 d | |
| Water fraction | 337.15 ± 7.86 e | 346.14 ± 4.44 g | |
| Acarbose | 13.33 ± 0.17 a | 0.051 ± 0.001 a | |
| Oral Digestion; Incubation Conditions: 37 °C, 2 min |
| 0.05–20 mg of bark or flower phenolic-rich extract |
| 1 mL of water |
| 1 mL of gelatinized potato starch (25 g/L) |
| 2.5 mL saliva solution (prepared according to [57]) |
| 0.5 mL α-amylase solution (0.1 mg/mL) |
| Gastric Digestion; Incubation Conditions: 37 °C, 2 h |
| 4.5 mL gastric solution (2 g NaCl in 0.7% HCl in water, pH 1.2) |
| 0.5 mL pepsin solution (3.2 mg/mL) |
| pH correction to a value of 2.0 with 2 M NaOH |
| Intestinal Digestion; Incubation Conditions: 37 °C, 2 h |
| 5 mL of water |
| pH correction to a value of 6.0 with 2 M NaOH followed to 7.5 with 1 M NHCO3 |
| The volume of the sample was adjusted to 16.4 mL with water |
| 1 mL of bile salts (100 mg/mL) |
| 2 mL of α-glucosidase solution |
| 0.6 mL of pancreatin solution (0.04 mg/mL) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kajszczak, D.; Kowalska-Baron, A.; Sosnowska, D.; Podsędek, A. In Vitro Inhibitory Effects of Viburnum opulus Bark and Flower Extracts on Digestion of Potato Starch and Carbohydrate Hydrolases Activity. Molecules 2022, 27, 3118. https://doi.org/10.3390/molecules27103118
Kajszczak D, Kowalska-Baron A, Sosnowska D, Podsędek A. In Vitro Inhibitory Effects of Viburnum opulus Bark and Flower Extracts on Digestion of Potato Starch and Carbohydrate Hydrolases Activity. Molecules. 2022; 27(10):3118. https://doi.org/10.3390/molecules27103118
Chicago/Turabian StyleKajszczak, Dominika, Agnieszka Kowalska-Baron, Dorota Sosnowska, and Anna Podsędek. 2022. "In Vitro Inhibitory Effects of Viburnum opulus Bark and Flower Extracts on Digestion of Potato Starch and Carbohydrate Hydrolases Activity" Molecules 27, no. 10: 3118. https://doi.org/10.3390/molecules27103118
APA StyleKajszczak, D., Kowalska-Baron, A., Sosnowska, D., & Podsędek, A. (2022). In Vitro Inhibitory Effects of Viburnum opulus Bark and Flower Extracts on Digestion of Potato Starch and Carbohydrate Hydrolases Activity. Molecules, 27(10), 3118. https://doi.org/10.3390/molecules27103118