
Co-Application of Statin and Flavonoids as an Effective Strategy to Reduce the Activity of Voltage-Gated Potassium Channels Kv1.3 and Induce Apoptosis in Human Leukemic T Cell Line Jurkat

 , and
, and
Abstract
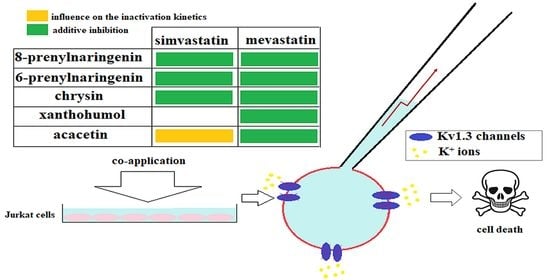
:
1. Introduction
2. Results
2.1. Electrophysiological Studies
2.2. Cell Viability in the Presence of Studied Compounds
2.3. Apoptosis Induction
3. Discussion
4. Material and Methods
4.1. Chemicals
4.2. Cell Culture and Solutions
4.3. Patch-Clamp Recordings
4.4. MTT Assay
4.5. Caspase-3 Activity
4.6. MMP Investigation
4.7. Western Blot Analysis
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| 8-PN | 8-prenylnaringenin. |
| 6-PN | 6–prenylnaringenin. |
| SIM | simvastatin. |
| ACAC | acacetin. |
| Xant | Xantohumol. |
| IC50 | half-maximal inhibitory concentration. |
| MMP | mitochondrial membrane potential. |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide. |
| pNA | chromophore p-nitroaniline. |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis. |
References
- Felipe, A.; Vincente, R.; Villalonga, N.; Roura-Ferrer, M.; Martinez-Marmol, R.; Sole, L.; Ferreres, J.C.; Condom, E. Potassium chanels: New targets in cancer therapy. Cancer Detect. Prev. 2006, 30, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Gutman, G.; Chandy, K.; Grissmer, S.; Lazdunski, M.; McKinnon, D.; Pardo, L.; Robertson, G.A.; Rudy, B.; Sanguinetti, M.C.; Stühmer, W.; et al. International Union of Pharmacology. LIII. Nomenclature and Molecular Relationships of Voltage-gated Potassium channels. Pharm. Rev. 2005, 67, 473–508. [Google Scholar] [CrossRef] [PubMed]
- Gulbins, E.; Sassi, N.; Grassme, H.; Zoratti, M.; Szabo, I. Role of Kv1.3 mitochondrial potassium channels in apoptotic signaling in lymphocytes. Biochim. Biophys. Acta BBA (Bioenerg.) 2010, 1797, 1251–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Verdaguer, M.; Caspera, J.; Serrano-Novillo, C.; Estadella, I.; Sastre, D.; Felipe, A. The voltage-gated potassium channel Kv1.3 is a promising multitherapeutic target against human pathologies. Exp. Opin. Ther. Targets 2016, 20, 577–591. [Google Scholar] [CrossRef]
- Cahalan, M.; Chandy, K. The functional network of ion channels in T lymphocytes. Immunol. Rev. 2009, 231, 59–87. [Google Scholar] [CrossRef] [Green Version]
- Kazama, I.; Tamada, T.; Tachi, M. Usefulness in targeting lymphocyte Kv1.3 channels in the treatment of respiratory diseases. Inflamm. Res. 2015, 64, 753–765. [Google Scholar] [CrossRef]
- Kazama, I. Targeting lymphocyte Kv1.3 channels to suppress cytokine storm in severe COVID-19: Can it be a novel therapeutic strategy? Drug Discov. Ther. 2020, 14, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Comes, N.; Bielanska, J.; Vallejo-Gracia, A.; Serrano-Albarras, A.; Marruecos, I.; Gomez, D.; Soler, C.; Condom, E.; Cajal, S.R.Y.; Hernández-Losa, J.; et al. The voltage-gated K+ channels Kv1.3 and Kv1.5 in human cancer. Front. Physiol. 2013, 4, 283. [Google Scholar] [CrossRef] [Green Version]
- Teisseyre, A.; Gąsiorowska, J.; Michalak, K. Voltage-gated potassium channels Kv1.3-potentially new molecular target in cancer diagnostics and therapy. Adv. Clin. Exp. Med. 2015, 24, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Teisseyre, A.; Palko-Labuz, A.; Środa-Pomianek, K.; Michalak, K. Voltage-gated potassium channel Kv1.3 as a target in therapy of cancer. Front. Oncol. 2019, 9, 933. [Google Scholar] [CrossRef]
- Serrano-Novillo, C.; Capera, J.; Colomer-Molera, M.; Condom, E.; Ferreres, J.; Felipe, A. Implication of voltage-gated potassium channels in neoplastic cell proliferation. Cancers 2019, 11, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teisseyre, A.; Uryga, A.; Michalak, K. Statins as inhibitors of voltage-gated potassium channels Kv1.3 in cancer cells. J. Mol. Struct. 2021, 1230, 129905. [Google Scholar] [CrossRef] [PubMed]
- Gazzerro, P.; Proto, M.; Gangemi, G.; Malfitano, A.; Ciaglia, E.; Pisanti, S.; Santoro, A.; Laezza, C.; Bifulco, M. Pharmacological actions of statins: A critical appraising in the management of cancer. Pharm. Rev. 2012, 64, 102–146. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D. Statins may be a key therapeutic for COVID-19. Med. Hypotheses 2020, 144, 110001. [Google Scholar] [CrossRef] [PubMed]
- Palko-Łabuz, A.; Środa-Pomianek, K.; Wesołowska, O.; Kustrzewa-Susłow, E.; Uryga, A.; Michalak, K. MDR reversal and pro-apoptotic effects of statins and statins combined with flavonoids in colon cancer cells. Biomed. Pharmacother. 2019, 109, 1511–1522. [Google Scholar] [CrossRef]
- Teisseyre, A.; Michalak, K. Inhibition of the activity of human lymphocyte Kv1.3 potassium channels by resveratrol. J. Membr. Biol. 2006, 214, 123–129. [Google Scholar] [CrossRef]
- Wu, X.; Song, M.; Qiu, P.; Rakariyatham, K.; Li, F.; Gao, Z.; Cai, X.; Wang, M.; Xu, F.; Zheng, J.; et al. Synergistic chemopreventive effects of nobiletin and atorvastatin on colon carcinogenesis. Carcinogenesis 2017, 38, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Zheng, J.; Bi, J.; Wu, X.; Lyu, J.; Gao, K. Synergistic inhibition of colon cancer cell growth by a combination of atorvastatin and phloretin. Oncol. Lett. 2018, 15, 1985–1992. [Google Scholar] [CrossRef] [Green Version]
- Attali, B.; Romey, G.; Honore, E.; Schmid-Alliana, A.; Mattei, M.; Lesage, F.; Ricard, P.; Barhanin, J.; Lazdunski, M. Cloning, functional expression, and regulation of two K+ channels in human T lymphocytes. J. Biol. Chem. 1992, 267, 8650–8657. [Google Scholar] [CrossRef]
- Valencia- Cruz, G.; Shabala, L.; Delgado- Enciso, I.; Shabala, S.; Bonales-Alatorre, E.; Pottosin, I.I.; Dobrovinskaya, O.R. Kbg and Kv1.3 channels mediate potassium efflux in the early phase of apoptosis in Jurkat T lymphocytes. Am. J. Physiol. Cell Physiol. 2009, 297, 1544–1553. [Google Scholar] [CrossRef] [Green Version]
- Teisseyre, A.; Palko-Labuz, A.; Uryga, A.; Michalak, K. The influence of 6-Prenylnaringenin and selected non-prenylated flavonoids on the activity of Kv1.3 channels in human Jurkat T cells. J. Membr. Biol. 2018, 251, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Gąsiorowska, J.; Teisseyre, A.; Uryga, A.; Michalak, K. Inhibition of Kv1.3 channels in human Jurkat T cells by xanthohumol and isoxanthohumol. J. Membr. Biol. 2015, 248, 705–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gąsiorowska, J.; Teisseyre, A.; Uryga, A.; Michalak, K. The influence of 8-prenylnaringenin on the activity of voltage-gated Kv1.3 potassium channels in human Jurkat T cells. Cell Mol. Biol. Lett. 2012, 17, 559–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teisseyre, A.; Mozrzymas, J. Inhibition of the Activity of T lymphocyte Kv1.3 Channels by Extracellular Zinc. Biochem. Pharm. 2002, 64, 595–607. [Google Scholar] [CrossRef]
- Ghosh-Choudhury, N.; Mandal, C.C.; Ghosh-Choudhury, N.; Ghosh-Choudhury, G. Simvastatin induces depression of PTEN expression via NFkappaB to inhibit breast cancer cell growth. Cell Signal. 2010, 22, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Klawitter, L.; Shokati, T.; Moll, V.; Christians, U.; Klawitter, J. Effects of lovastatin on breast cancer cells: A proteo-metabonomic study. Breast Cancer Res. 2010, 12, R16. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.J.; Kim, J.S.; Kim, J.M.; Lee, J.Y.; Jung, H.C.; Song, I.S. Simvastatin induces apoptosis in human colon cancer cells and in tumor xenografts, and attenuates colitis-associated colon cancer in mic. Int. J. Cancer 2008, 123, 951–957. [Google Scholar] [CrossRef]
- Yang, Y.C.; Huang, W.F.; Chuan, L.M.; Xiao, D.W.; Zeng, Y.L.; Zhou, D.A.; Xu, G.Q.; Liu, W.; Huang, B.; Hu, Q. In vitro and in vivo study of cell growth inhibition of simvastatin on chronic myelogenous leukemia cells. Chemotherapy 2008, 54, 438–446. [Google Scholar] [CrossRef]
- Fan, T.J.; Han, L.H.; Cong, R.S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.S.; Zhang, S. Polypharmacology: Drug discovery for the future. Exp. Rev. Clin. Pharmacol. 2013, 6, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Kanno, S.; Tomizawa, A.; Yomogida, S.; Ishikawa, M. Acacetin induces apoptosis in human T cell leukemia Jurkat cells via activation of a caspase cascade. Oncol. Rep. 2012, 27, 204–209. [Google Scholar] [PubMed]
- Xue, C.; Chen, Y.; Hu, D.N.; Lacob, C.; Lu, C.; Huang, Z. Chrysin induces cell apoptosis in human uveal melanoma cells via intrinsic apoptosis. Oncol. Lett. 2016, 12, 4813–4820. [Google Scholar] [CrossRef] [PubMed]
- Salimi, A.; Roudkenar, M.H.; Sadeghi, L.; Mohseni, A.; Seydi, E.; Pirahmad, N.; Pourahmad, J. Selective anticancer activity of acacetin against chronic lymphocytic leukemia using both in vivo and in vitro methods: Key role of oxidative stress and cancerous mitochondria. Nutr. Cancer 2016, 68, 1404–1416. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, S.H.; Nafees, S.; Khan, A.; Rizvi, M.A. Chrysin: A promising anticancer agent its current trends andfuture imedpub journals chrysin: A promising anticancer agent its current trends and future perspectives. Eur. J. Exp. Biol. 2018, 8, 16. [Google Scholar] [CrossRef]
- Kasala, E.R.; Bodduluru, L.N.; Madana, R.M.; Athira, K.V.; Gogoi, R.; Barua, C.C. Chemopreventive and Therapeutic Potential of Chrysin in Cancer: Mechanistic Perspectives. Toxicol. Lett. 2015, 233, 214–225. [Google Scholar] [CrossRef]
- Grissmer, S.; Nguyen, A.; Cahalan, M. Calcium-activated potassium channels in resting and activated human T lymphocytes. J. Gen. Physiol. 1993, 102, 601–630. [Google Scholar] [CrossRef]
- Grissmer, S.; Lewis, R.; Cahalan, M. Ca2+-activated K+ Channels in Human Leukemic T Cells. J. Gen. Physiol. 1992, 99, 63–84. [Google Scholar] [CrossRef] [Green Version]
- Hamill, O.P.; Marty, A.; Neher, E.; Sakmann, B.; Sigworth, F.J. Improved patch-clamp techniques for high- resolution current recording from cells and cell-free membrane patches. Pfluegers Arch. 1981, 39, 85–100. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SIM Co-Applied with: | 8-PR | 6-PR | Xant | ACAC | Chrysin |
|---|---|---|---|---|---|
| Theoretical values | 0.195 | 0.395 | 0.36 | 0.32 | 0.285 |
| Experimental values | 0.12 | 0.26 | 0.50 | 0.53 | 0.25 |
| Difference | Significant (+) | Significant (+) | Significant (−) | Significant (−) | Not significant |
| MEV co-applied with: | 8-PR | 6-PR | Xant | ACAC | Chrysin |
| Theoretical values | 0.16 | 0.32 | 0.31 | 0.26 | 0.23 |
| Experimental values | 0.09 | 0.29 | 0.30 | 0.30 | 0.26 |
| Difference | Significant (+) | Not significant | Not significant | Not significant | Not significant |
| No Statin | Simvastatin | Mevastatin | |
|---|---|---|---|
| 8-prenylnaringenin | n.a. | 26.9 µM | 7.1 µM |
| 6-prenylnaringenin | n.a. [19] | 38.9 µM | 34.8 µM |
| chrysin | 26.2 µM [19] | 10.8 µM | 8.3 µM |
| xanthohumol | 32.5 µM | 30.8 µM | 3.8 µM |
| acacetin | n.a. [19] | n.a. | n.a. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teisseyre, A.; Chmielarz, M.; Uryga, A.; Środa-Pomianek, K.; Palko-Łabuz, A. Co-Application of Statin and Flavonoids as an Effective Strategy to Reduce the Activity of Voltage-Gated Potassium Channels Kv1.3 and Induce Apoptosis in Human Leukemic T Cell Line Jurkat. Molecules 2022, 27, 3227. https://doi.org/10.3390/molecules27103227
Teisseyre A, Chmielarz M, Uryga A, Środa-Pomianek K, Palko-Łabuz A. Co-Application of Statin and Flavonoids as an Effective Strategy to Reduce the Activity of Voltage-Gated Potassium Channels Kv1.3 and Induce Apoptosis in Human Leukemic T Cell Line Jurkat. Molecules. 2022; 27(10):3227. https://doi.org/10.3390/molecules27103227
Chicago/Turabian StyleTeisseyre, Andrzej, Mateusz Chmielarz, Anna Uryga, Kamila Środa-Pomianek, and Anna Palko-Łabuz. 2022. "Co-Application of Statin and Flavonoids as an Effective Strategy to Reduce the Activity of Voltage-Gated Potassium Channels Kv1.3 and Induce Apoptosis in Human Leukemic T Cell Line Jurkat" Molecules 27, no. 10: 3227. https://doi.org/10.3390/molecules27103227
APA StyleTeisseyre, A., Chmielarz, M., Uryga, A., Środa-Pomianek, K., & Palko-Łabuz, A. (2022). Co-Application of Statin and Flavonoids as an Effective Strategy to Reduce the Activity of Voltage-Gated Potassium Channels Kv1.3 and Induce Apoptosis in Human Leukemic T Cell Line Jurkat. Molecules, 27(10), 3227. https://doi.org/10.3390/molecules27103227