
Pharmacological, Neurochemical, and Behavioral Mechanisms Underlying the Anxiolytic- and Antidepressant-like Effects of Flavonoid Chrysin

 , and
, and
Abstract
:1. Introduction
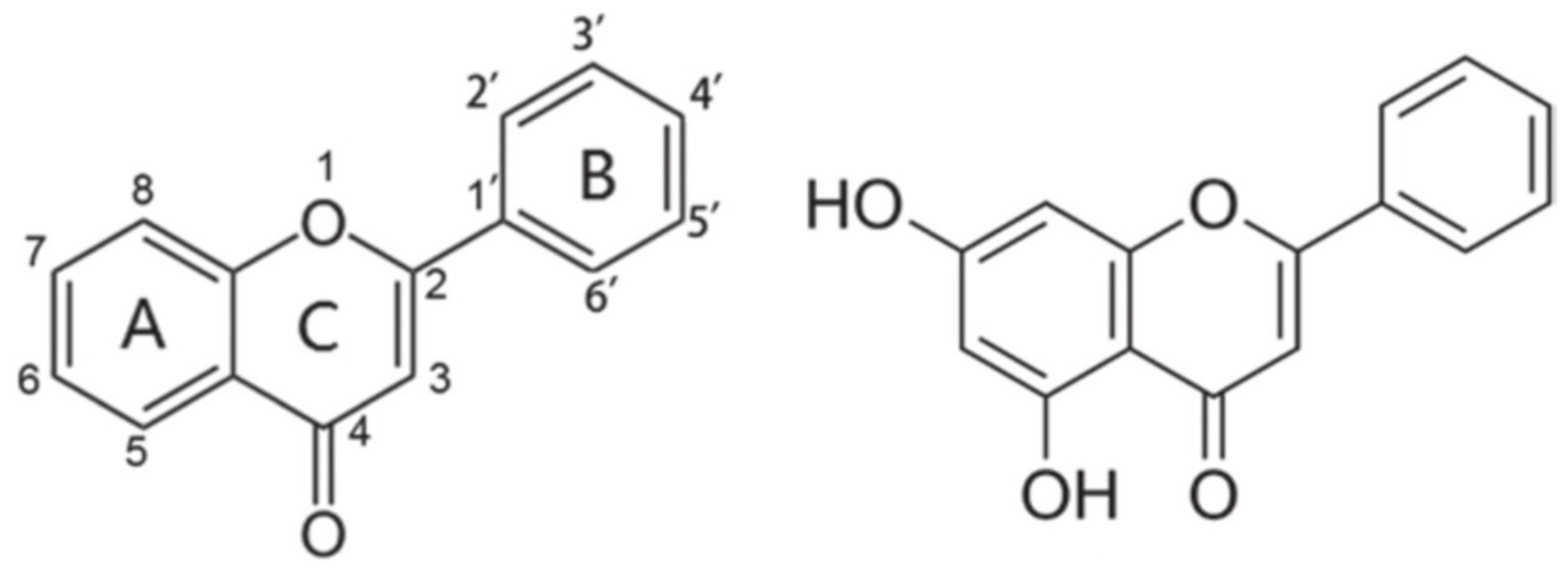
2. Generalities of the Flavonoid Chrysin
3. Biochemical and Pharmacological Activity of Flavonoid Chrysin
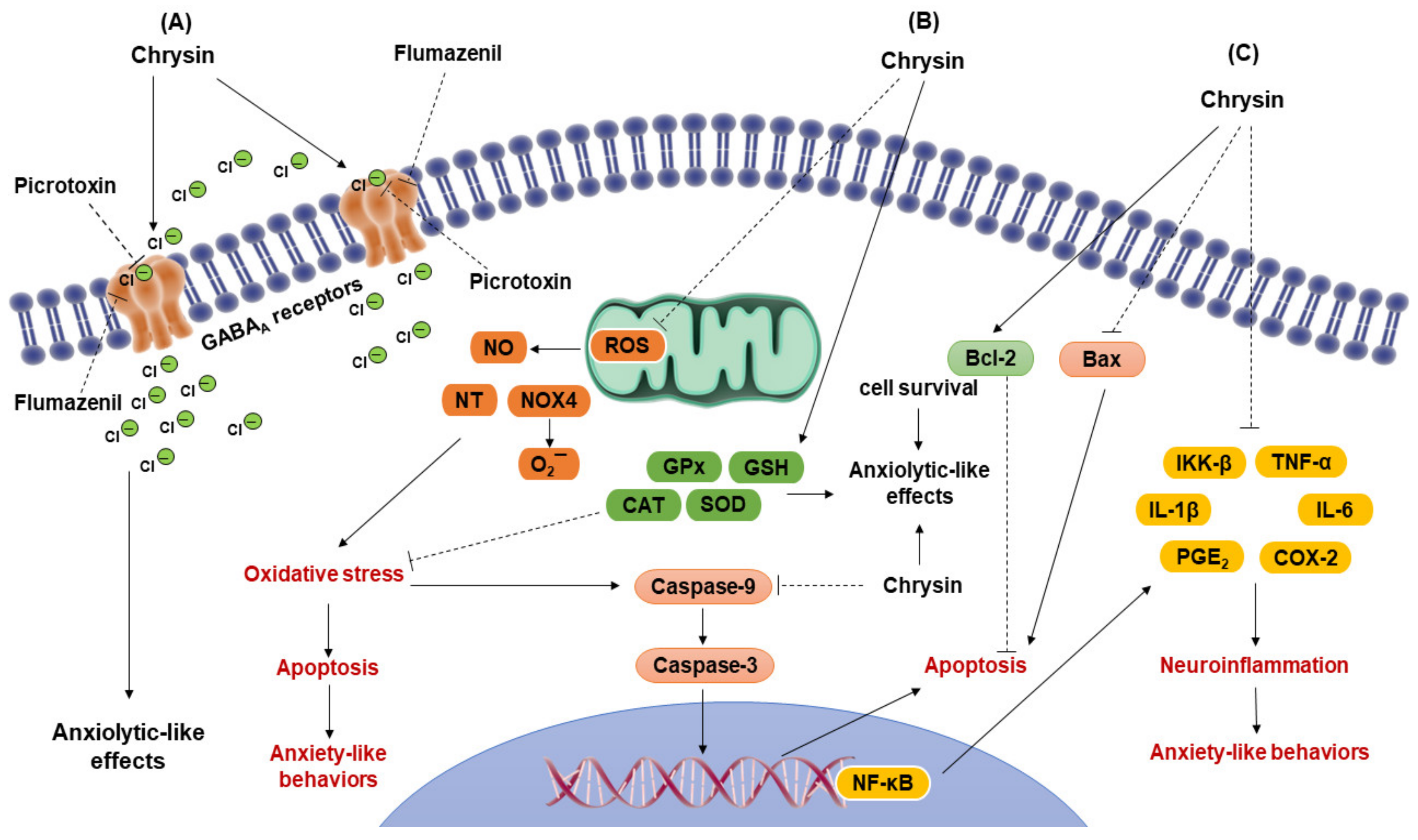
3.1. Action of Chrysin on Neurotransmission Systems
3.2. Antioxidant Activity of Chrysin
3.3. Anti-Inflammatory and Anti-Apoptotic Activity of Chrysin
3.4. Effects of Chrysin on Gut Microbiota
4. Anxiolytic-like Effects of Flavonoid Chrysin
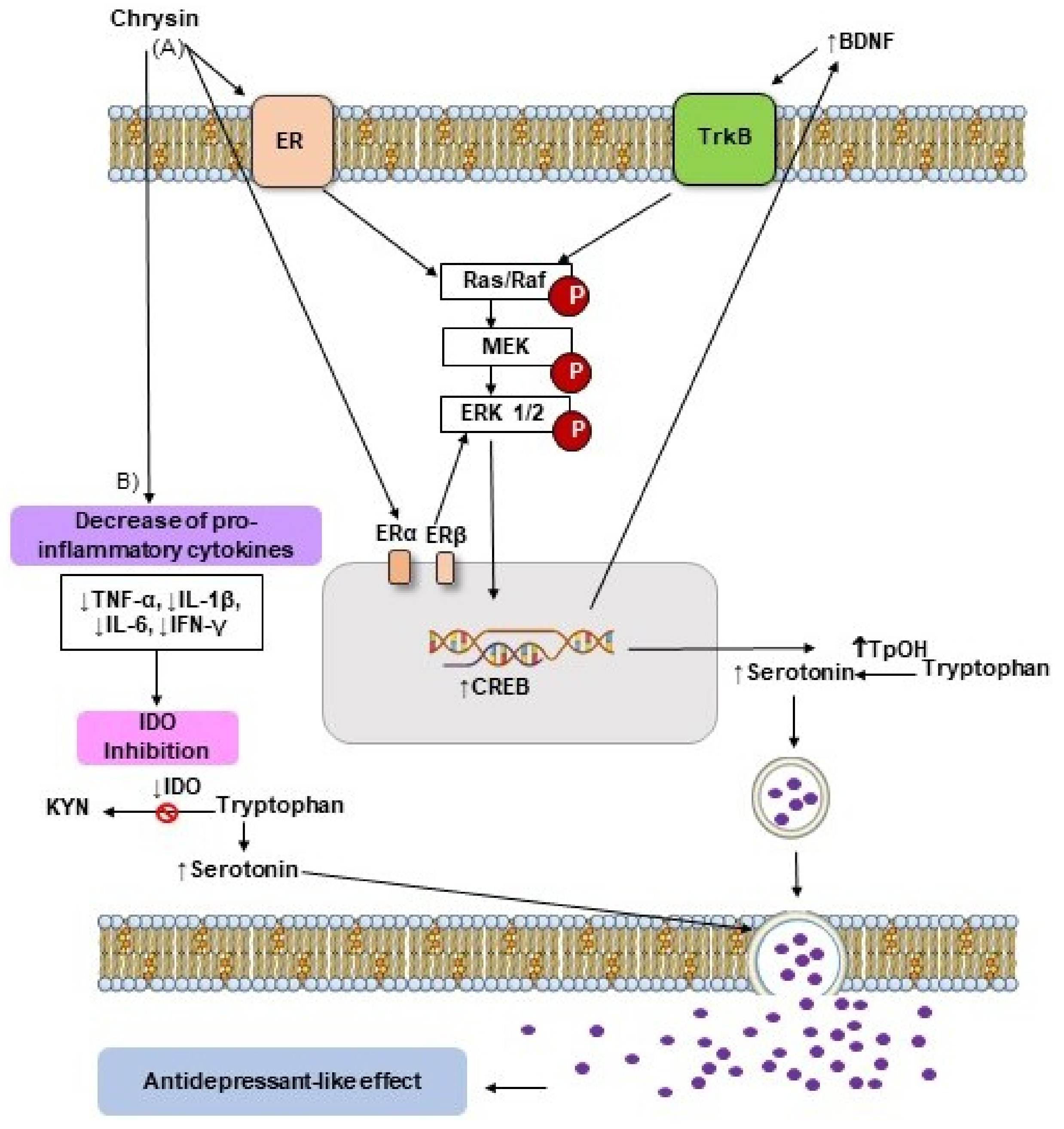
5. Antidepressant-like Effects of Flavonoid Chrysin
6. Future Considerations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferreira, M.J.U. Natural products in drug discovery and human health. Phytochem. Rev. 2021, 20, 1–4. [Google Scholar] [CrossRef]
- Tresina, P.S.; Selvam, M.S.; Rajesh, A.; Doss, A.; Mohan, V.R. Natural products in drug discovery: Approaches and development. J. Pharm. Res. Int. 2021, 33, 93–110. [Google Scholar] [CrossRef]
- Dzobo, K. The role of natural products as sources of therapeutic agents for innovative drug discovery. In Comprehensive Pharmacology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–15. [Google Scholar] [CrossRef]
- Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From theory to practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef] [PubMed]
- German-Ponciano, L.J.; Rosas-Sánchez, G.U.; Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F. Advances in the preclinical study of some flavonoids as potential antidepressant agents. Scientifica 2018, 2018, 2963565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Demeneghi, R.; Rodríguez-Landa, J.F.; Guzmán-Gerónimo, R.I.; Acosta-Mesa, H.G.; Meza-Alvarado, E.; Vargas-Moreno, I.; Herrera-Meza, S. Effect of blackberry juice (Rubus fruticosus L.) on anxiety-like behaviour in Wistar rats. Int. J. Food Sci. Nutr. 2018, 70, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Jayaraman, A.; Chapkin, R.S.; Howard, M.; Mohankumar, K.; Shrestha, R. Flavonoids: Structure-function and mechanisms of action and opportunities for drug development. Toxicol. Res. 2021, 37, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Chaturvedi, S. A comprehensive review on chrysin: Emphasis on molecular targets, pharmacological actions and bio-pharmaceutical aspects. Curr. Drug Targets 2022, 23, 420–436. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Mishra, P.S.; Bandopadhyay, R.; Khurana, N.; Angelopoulou, E.; Paudel, Y.N.; Piperi, C. Neuroprotective potential of chrysin: Mechanistic insights and therapeutic potential for neurological disorders. Molecules 2021, 26, 6456. [Google Scholar] [CrossRef]
- Talebi, M.; Talebi, M.; Farkhondeh, T.; Kopustinskiene, D.M.; Simal-Gandara, J.; Bernatoniene, J.; Samarghandian, S. An updated review on the versatile role of chrysin in neurological diseases: Chemistry, pharmacology, and drug delivery approaches. Biomed. Pharm. 2021, 141, 111906. [Google Scholar] [CrossRef]
- Filho, C.B.; Jesse, C.R.; Donato, F.; Giacomeli, R.; Del Fabbro, L.; da Silva Antunes, M.; de Gomes, M.G.; Goes, A.T.R.; Boeira, S.P.; Prigol, M.; et al. Chronic unpredictable mild stress decreases BDNF and NGF levels and Na+,K+-ATPase activity in the hippocampus and prefrontal cortex of mice: Antidepressant effect of chrysin. Neuroscience 2015, 289, 367–380. [Google Scholar] [CrossRef]
- Bortolotto, V.C.; Pinheiro, F.C.; Araujo, S.M.; Poetini, M.R.; Bertolazi, B.S.; de Paula, M.T.; Meichtry, L.B.; de Almeida, F.P.; de Freitas Couto, S.; Jesse, C.R.; et al. Chrysin reverses the depressive-like behavior induced by hypothyroidism in female mice by regulating hippocampal serotonin and dopamine. Eur. J. Pharmacol. 2018, 822, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Germán-Ponciano, L.J.; Rosas-Sánchez, G.U.; Ortiz-Guerra, S.I.; Soria-Fregozo, C.; Rodríguez-Landa, J.F. Effects of chrysin on mRNA expression of 5-HT1A and 5-HT2A receptors in the raphe nuclei and hippocampus. Rev. Bras. Farmacog. 2021, 31, 352–360. [Google Scholar] [CrossRef]
- Rodríguez-Landa, J.F.; Hernández-López, F.; Martínez-Mota, L.; Scuteri, D.; Bernal-Morales, B.; Rivadeneyra-Domínguez, E. GABAA/benzodiazepine receptor complex in the dorsal hippocampus mediates the effects of chrysin on anxiety-like behaviour in female rats. Front. Behav. Neurosci. 2022, 15, 789557. [Google Scholar] [CrossRef]
- Medina, J.H.; Paladini, A.C.; Wolfman, C.; Levi de Stein, M.; Calvo, D.; Diaz, L.E.; Peña, C. Chrysin (5,7-di-OH-flavone), a naturally-occurring ligand for benzodiazepine receptors, with anticonvulsant properties. Biochem. Pharmacol. 1990, 40, 2227–2231. [Google Scholar] [CrossRef]
- Salgueiro, J.B.; Ardenghi, P.; Dias, M.; Ferreira, M.B.; Izquierdo, I.; Medina, J.H. Anxiolytic natural and synthetic flavonoid ligands of the central benzodiazepine receptor have no effect on memory tasks in rats. Pharmacol. Biochem. Behav. 1997, 58, 887–891. [Google Scholar] [CrossRef]
- Zanoli, P.; Avallone, R.; Baraldi, M. Behavioral characterisation of the flavonoids apigenin and chrysin. Fitoterapia 2000, 71, S117–S123. [Google Scholar] [CrossRef]
- Filho, C.B.; Jesse, C.R.; Donato, F.; Del Fabbro, L.; de Gomes, M.G.; Goes, A.T.R.; Souza, L.C.; Giacomeli, R.; Antunes, M.; Luchese, C.; et al. Neurochemical factors associated with the antidepressant-like effect of flavonoid chrysin in chronically stressed mice. Eur. J. Pharmacol. 2016, 791, 284–296. [Google Scholar] [CrossRef]
- Goes, A.T.; Jesse, C.R.; Antunes, M.S.; Ladd, F.V.L.; Ladd, A.A.L.; Luchese, C.; Paroul, N.; Boeira, S.P. Protective role of chrysin on 6-hydroxydopamine-induced neurodegeneration a mouse model of Parkinson’s disease: Involvement of neuroinflammation and neurotrophins. Chem. Biol. Interact. 2018, 279, 111–120. [Google Scholar] [CrossRef]
- Souza, L.C.; Antunes, M.S.; Filho, C.B.; Del Fabbro, L.; de Gomes, M.G.; Goes, A.T.; Donato, F.; Prigol, M.; Boeira, S.P.; Jesse, C.R. Flavonoid chrysin prevents age-related cognitive decline via attenuation of oxidative stress and modulation of BDNF levels in aged mouse brain. Pharmacol. Biochem. Behav. 2015, 134, 22–30. [Google Scholar] [CrossRef]
- Feiger, J.A.; Snyder, R.L.; Walsh, M.J.; Cissne, M.; Cwiek, A.; Al-Momani, S.I.; Chiou, K.S. The role of neuroinflammation in neuropsychiatric disorders following traumatic brain injury: A systematic review. J. Head Trauma Rehabil. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Sathiavelu, J.; Senapathy, G.J.; Devaraj, R.; Namasivayam, N. Hepatoprotective effect of chrysin on prooxidant-antioxidant status during ethanol-induced toxicity in female albino rats. J. Pharm. Pharmacol. 2009, 61, 809–817. [Google Scholar] [CrossRef] [PubMed]
- German-Ponciano, L.J.; Costa, B.P.D.; Feitosa, L.M.; dos Santos Campos, K.; da Silva Chaves, S.N.; Cueto-Escobedo, J.; Maximino, C. Chrysin, but not flavone backbone, decreases anxiety-like behavior in animal screens. Neurochem. Int. 2020, 140, 104850. [Google Scholar] [CrossRef]
- Wolfman, C.; Viola, H.; Paladini, A.; Dajas, F.; Medina, J.H. Possible anxiolytic effects of chrysin, a central benzodiazepine receptor ligand isolated from Passiflora Coerulea. Pharmacol. Biochem. Behav. 1994, 47, 1–4. [Google Scholar] [CrossRef]
- Filho, C.B.; Jesse, C.R.; Donato, F.; Del Fabbro, L.; Gomes de Gomes, M.; Rossito Goes, A.T.; Souza, L.C.; Boeira, S.P. Chrysin promotes attenuation of depressive-like behavior and hippocampal dysfunction resulting from olfactory bulbectomy in mice. Chem. Biol. Interact. 2016, 260, 154–162. [Google Scholar] [CrossRef]
- Rodríguez-Landa, J.F.; Hernández-López, F.; Cueto-Escobedo, J.; Herrera-Huerta, E.V.; Rivadeneyra-Domínguez, E.; Bernal-Morales, B.; Romero-Avendaño, E. Chrysin (5,7-dihydroxyflavone) exerts anxiolytic-like effects through GABAA receptors in a surgical menopause model in rats. Biomed. Pharmacother. 2019, 109, 2387–2395. [Google Scholar] [CrossRef]
- Mantawy, E.M.; El-Bakly, W.M.; Esmat, A.; Badr, A.M.; El-Demerdash, E. Chrysin alleviates acute doxorubicin cardiotoxicity in rats via suppression of oxidative stress, inflammation and apoptosis. Eur. J. Pharmacol. 2014, 728, 107–118. [Google Scholar] [CrossRef]
- Rani, N.; Bharti, S.; Bhatia, J.; Tomar, A.; Nag, T.C.; Ray, R.; Arya, D.S. Inhibition of TGF-β by a novel PPAR-γ agonist, chrysin, salvages β-receptor stimulated myocardial injury in rats through MAPKs-dependent mechanism. Nutr. Metab. 2015, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Rani, N.; Bharti, S.; Bhatia, J.; Nag, T.C.; Ray, R.; Arya, D.S. Chrysin, a PPAR-γ agonist improves myocardial injury in diabetic rats through inhibiting AGE-RAGE mediated oxidative stress and inflammation. Chem. Biol. Interact. 2016, 250, 59–67. [Google Scholar] [CrossRef]
- Campos, H.M.; da Costa, M.; da Silva Moreira, L.K.; da Silva Neri, H.F.; Branco da Silva, C.R.; Pruccoli, L.; Dos Santos, F.C.A.; Costa, E.A.; Tarozzi, A.; Ghedini, P.C. Protective effects of chrysin against the neurotoxicity induced by aluminium: In Vitro and In Vivo studies. Toxicology 2022, 465, 153033. [Google Scholar] [CrossRef]
- Harasstani, O.A.; Moin, S.; Tham, C.L.; Liew, C.Y.; Ismail, N.; Rajajendram, R.; Harith, H.H.; Zakaria, Z.A.; Mohamad, A.S.; Sulaiman, M.R.; et al. Flavonoid combinations cause synergistic inhibition of proinflammatory mediator secretion from lipopolysaccharide-induced RAW 264.7 cells. Inflamm. Res. 2010, 59, 711–721. [Google Scholar] [CrossRef]
- Feng, X.; Qin, H.; Shi, Q.; Zhang, Y.; Zhou, F.; Wu, H.; Ding, S.; Niu, Z.; Lu, Y.; Shen, P. Chrysin attenuates inflammation by regulating M1/M2 status via activating PPARγ. Biochem. Pharmacol. 2014, 89, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Harasstani, O.A.; Tham, C.L.; Israf, D.A. Kaempferol and chrysin synergies to improve septic mice survival. Molecules 2017, 22, 92. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hui, K.M.; Chen, Y.; Xu, S.; Wong, J.T.; Xue, H. Structure-activity relationships of flavonoids, isolated from Scutellaria baicalensis, binding to benzodiazepine site of GABAA receptor complex. Planta Med. 2002, 68, 1059–1062. [Google Scholar] [CrossRef] [PubMed]
- Goutman, J.D.; Waxemberg, M.D.; Doñate-Oliver, F.; Pomata, P.E.; Calvo, D.J. Flavonoid modulation of ionic currents mediated by GABAA and GABAc receptors. Eur. J. Pharmacol. 2003, 461, 79–87. [Google Scholar] [CrossRef]
- Rodríguez-Landa, J.F.; Guillén-Ruiz, G.; Hernández-López, F.; Cueto-Escobedo, J.; Rivadeneyra-Domínguez, E.; Bernal-Morales, B.; Herrera-Huerta, E.V. Chrysin reduces anxiety-like behavior through actions on GABAA receptors during metestrus-diestrus in the rat. Behav. Brain Res. 2021, 397, 112952. [Google Scholar] [CrossRef]
- Cueto-Escobedo, J.; Andrade-Soto, J.; Lima-Maximino, M.; Maximino, C.; Hernández-López, F.; Rodríguez-Landa, J.F. Involvement of GABAergic system in the antidepressant-like effects of chrysin (5, 7-dihydroxyflavone) in ovariectomized rats in the forced swim test: Comparison with neurosteroids. Behav. Brain Res. 2020, 386, 112590. [Google Scholar] [CrossRef]
- Haider, M.; Salman, M.; Kaushik, P.; Bharadwaj, N.; Aggarwal, N.B.; Tabassum, H.; Parvez, S. Chrysin ameliorates 3 nitropropinoic acid induced neurotoxicity targeting behavioural, biochemical and histological alterations. Int. J. Neurosci. 2020, 1–9, Online ahead of print. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Cui, W.; Zhou, W.; Zhao, X. 5-HT1A receptor-mediated attenuation of heat hyperalgesia and mechanical allodynia by chrysin in mice with experimental mononeuropathy. Reg. Anesth. Pain Med. 2020, 45, 610–619. [Google Scholar] [CrossRef]
- Krishnamoorthy, A.; Sevanan, M.; Mani, S.; Balu, M.; Balaji, S.; Ramajayan, P. Chrysin restores MPTP induced neuroinflammation, oxidative stress and neurotrophic factors in an acute Parkinson’s disease mouse model. Neurosci. Lett. 2019, 709, 134382. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Samarghandian, S.; Azimin-Nezhad, M.; Samini, F. Effect of chrysin on nociception in formalin test and serum levels of noradrenalin and corticosterone in rats. Int. J. Clin. Exp. Med. 2015, 8, 2465. [Google Scholar]
- Rashno, M.; Ghaderi, S.; Nesari, A.; Khorsandi, L.; Farbood, Y.; Sarkaki, A. Chrysin attenuates traumatic brain injury-induced recognition memory decline, and anxiety/depression-like behaviors in rats: Insights into underlying mechanisms. Psychopharmacology 2020, 237, 1607–1619. [Google Scholar] [CrossRef]
- Bortolotto, V.C.; Araujo, S.M.; Pinheiro, F.C.; Poetini, M.R.; Meichtry, L.B.; Fronza, M.G.; Boeira, S.P.; Savegnago, L.; Prigol, M. Chrysin restores memory deficit in hypothyroidism mice: Behavioral, neurochemical and computational approaches involving the neurotrophinergic system. J. Psychiatr. Res. 2021, 144, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, B.; Bhatia, H.S.; Bouchard, M.; Fiebich, B.L.; Fetoui, H. Inflammatory and oxidative mechanisms potentiate bifenthrin-induced neurological alterations and anxiety-like behavior in adult rats. Toxicol. Lett. 2018, 294, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, S.; Abramov, A.Y. Mechanism of oxidative stress in neurodegeneration. Oxid. Med. Cell Longev. 2012, 2012, 428010. [Google Scholar] [CrossRef] [Green Version]
- Smaga, I.; Niedzielska, E.; Gawlik, M.; Moniczewski, A.; Krzek, J.; Przegaliński, E.; Pera, J.; Filip, M. Oxidative stress as an etiological factor and a potential treatment target of psychiatric disorders. Part 2. Depression, anxiety, schizophrenia and autism. Pharmacol. Rep. 2015, 67, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Nagappa, A.N.; Patil, C.R. Role of oxidative stress in depression. Drug Discov. Today 2020, 25, 1270–1276. [Google Scholar] [CrossRef] [PubMed]
- Di Sarno, R.; Brigida, A.; Caprio, G.G.; Ciardiello, D.; Dallio, M.; Sangineto, M.; Fagoonee, S.; Abenavoli, L.; Luzza, F.; Gravina, A.G.; et al. Critical review on the use and abuse of alcohol. When the dose makes the difference. Minerva Med. 2020, 111, 344–353. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef]
- Hovatta, I.; Juhila, J.; Donner, J. Oxidative stress in anxiety and comorbid disorders. Neurosci. Res. 2010, 68, 261–275. [Google Scholar] [CrossRef]
- Sarwar, H.; Rafiqi, S.I.; Ahmad, S.; Jinna, S.; Khan, S.A.; Karim, T.; Qureshi, O.; Zahid, Z.A.; Elhai, J.D.; Levine, J.C.; et al. Hyperinsulinemia associated depression. Clin. Med. Insights Endocrinol. Diabetes 2022, 15, 11795514221090244. [Google Scholar] [CrossRef]
- Bouayed, J.; Rammal, H.; Younos, C.; Soulimani, R. Positive correlation between peripheral blood granulocyte oxidative status and level of anxiety in mice. Eur. J. Pharmacol. 2007, 564, 146–149. [Google Scholar] [CrossRef]
- Salim, S.; Sarraj, N.; Taneja, M.; Saha, K.; Tejada-Simon, M.V.; Chugh, G. Moderate treadmill exercise prevents oxidative stress-induced anxiety-like behavior in rats. Behav. Brain Res. 2010, 208, 545–552. [Google Scholar] [CrossRef]
- Kaufmann, F.N.; Gazal, M.; Mondin, T.C.; Cardoso, T.A.; Quevedo, L.Á.; Souza, L.D.; Ghisleni, G. Cognitive psychotherapy treatment decreases peripheral oxidative stress parameters associated with major depression disorder. Biol. Psychol. 2015, 110, 175–181. [Google Scholar] [CrossRef]
- Moccia, M.; Capacchione, A.; Lanzillo, R.; Carbone, F.; Micillo, T.; Perna, F.; Brescia Morra, V. Coenzyme Q10 supplementation reduces peripheral oxidative stress and inflammation in interferon-β1a-treated multiple sclerosis. Ther. Adv. Neurol. Disord. 2019, 12, 1756286418819074. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, Y.; To, M.; Saruta, J.; Yamamoto, Y.; Yamamoto, T.; Shimizu, T.; Kamata, Y.; Matsuo, M.; Tsukinoki, K. Effect of social isolation stress on saliva BDNF in rat. J. Oral Sci. 2019, 61, 516–520. [Google Scholar] [CrossRef] [Green Version]
- Khurana, K.; Bansal, N. Lacidipine attenuates caffeine-induced anxiety-like symptoms in mice: Role of calcium-induced oxido-nitrosative stress. Pharmacol. Rep. 2019, 71, 1264–1272. [Google Scholar] [CrossRef]
- Risbrough, V.B.; Vaughn, M.N.; Friend, S.F. Role of inflammation in traumatic brain injury-associated risk for neuropsychiatric disorders: State of the evidence and where do we go from here. Biol. Psychiatry 2022, 91, 438–448. [Google Scholar] [CrossRef]
- Zorzo, C.; Méndez-López, M.; Méndez, M.; Arias, J.L. Adult social isolation leads to anxiety and spatial memory impairment: Brain activity pattern of COx and c-Fos. Behav. Brain Res. 2019, 365, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Park, S.S.; Park, H.S.; Kim, C.J.; Baek, S.S.; Kim, T.W. Exercise attenuates maternal separation-induced mood disorder-like behaviors by enhancing mitochondrial functions and neuroplasticity in the dorsal raphe. Behav. Brain Res. 2019, 372, 112049. [Google Scholar] [CrossRef]
- Zhang, L.X.; Levine, S.; Dent, G.; Zhan, Y.; Xing, G.; Okimoto, D.; Kathleen Gordon, M.; Post, R.M.; Smith, M.A. Maternal deprivation increases cell death in the infant rat brain. Brain Res. Dev. Brain Res. 2002, 133, 1–11. [Google Scholar] [CrossRef]
- Wang, Q.; Dong, X.; Wang, Y.; Liu, M.; Sun, A.; Li, N.; Lin, Y.; Geng, Z.; Jin, Y.; Li, X. Adolescent escitalopram prevents the effects of maternal separation on depression- and anxiety-like behaviours and regulates the levels of inflammatory cytokines in adult male mice. Int. J. Dev. Neurosci. 2017, 62, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rivera, N.M.; Fernández-Guasti, A.; Ramírez-Rodríguez, G.; Estrada-Camarena, E. Acute stress further decreases the effect of ovariectomy on immobility behavior and hippocampal cell survival in rats. Psychoneuroendocrinology 2013, 38, 1407–1417. [Google Scholar] [CrossRef]
- Wang, Z.; Gu, J.; Wang, X.; Xie, K.; Luan, Q.; Wan, N.; Zhang, Q.; Jiang, H.; Liu, D. Antidepressant-like activity of resveratrol treatment in the forced swim test and tail suspension test in mice: The HPA axis, BDNF expression and phosphorylation of ERK. Pharmacol. Biochem. Behav. 2013, 112, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Abd Al Haleem, E.N.; Ahmed, H.I.; El-Naga, R.N. Lycopene and chrysin through mitigation of neuroinflammation and oxidative stress exerted antidepressant effects in clonidine-induced depression-like behavior in rats. J. Diet Suppl. 2021, 1–20, Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Bear, T.L.; Dalziel, J.E.; Coad, J.; Roy, N.C.; Butts, C.A.; Gopal, P.K. The role of the gut microbiota in dietary interventions for depression and anxiety. Adv. Nutr. 2020, 11, 890–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonali, S.; Ray, B.; Ahmed Tousif, H.; Rathipriya, A.G.; Sunanda, T.; Mahalakshmi, A.M.; Rungratanawanich, W.; Essa, M.M.; Qoronfleh, M.W.; Chidambaram, S.B.; et al. Mechanistic insights into the link between gut dysbiosis and major depression: An extensive review. Cells 2022, 11, 1362. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Borre, Y.; O’Brien, C.; Patterson, E.; El Aidy, S.; Deane, J.; Dinan, T.G. Transferring the blues: Depression-associated gut microbiota induces neurobehavioural changes in the rat. J. Psychiatr. Res. 2016, 82, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.; Qiao, Y.; Xie, R.; Lin, L.; Jiang, J.; Wang, C.; Li, G. Fecal microbiota transplantation ameliorates stress-induced depression-like behaviors associated with the inhibition of glial and NLRP3 inflammasome in rat brain. J. Psychiatr. Res. 2021, 137, 147–157. [Google Scholar] [CrossRef]
- Hervert-Hernández, D.; Goñi, I. Dietary polyphenols and human gut microbiota: A review. Food Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Hong, M.; Zhang, R.; Liu, Y.; Wu, Z.; Weng, P. The interaction effect between tea polyphenols and intestinal microbiota: Role in ameliorating neurological diseases. J. Food Biochem. 2022, 46, e13870. [Google Scholar] [CrossRef]
- Zhou, N.; Gu, X.; Zhuang, T.; Xu, Y.; Yang, L.; Zhou, M. Gut microbiota: A pivotal hub for polyphenols as antidepressants. J. Agric. Food Chem. 2020, 68, 6007–6020. [Google Scholar] [CrossRef]
- Cheng, N.; Chen, S.; Liu, X.; Zhao, H.; Cao, W. Impact of schisandra chinensis bee pollen on nonalcoholic fatty liver disease and gut microbiota in high fat diet induced obese mice. Nutrients 2019, 11, 346. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.T.; Wang, Z.H. Effects of chrysin on intestinal inflammation and gut microbiota in lipopolysaccharides-induced mice. FASEB J. 2019, 33, 764–768. [Google Scholar] [CrossRef]
- Wen, X.; Walle, T. Methylated flavonoids have greatly improved intestinal absorption and metabolic stability. Drug Metab. Dispos. 2006, 34, 1786–1792. [Google Scholar] [CrossRef] [Green Version]
- Paladini, A.C.; Marder, M.; Viola, H.; Wolfman, C.; Wasowski, C.; Medina, J.H. Flavonoids and the central nervous system: From forgotten factors to potent anxiolytic compounds. J. Pharm. Pharmacol. 1999, 51, 519–526. [Google Scholar] [CrossRef]
- Brown, E.; Hurd, N.S.; McCall, S.; Ceremuga, T.E. Evaluation of the anxiolytic effects of chrysin, a Passiflora incarnata extract, in the laboratory rat. AANA J. 2007, 75, 333–337. [Google Scholar]
- Germán-Ponciano, L.J.; Puga-Olguín, A.; Rovirosa-Hernández, M.J.; Caba, M.; Meza, E.; Rodríguez-Landa, J.F. Differential effects of acute and chronic treatment with the flavonoid chrysin on anxiety-like behavior and Fos immunoreactivity in the lateral septal nucleus in rats. Acta Pharm. 2020, 70, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Ognibene, E.; Bovicelli, P.; Adriani, W.; Saso, L.; Laviola, G. Behavioral effects of 6-bromoflavanone and 5-methoxy-6,8-dibromoflavanone as anxiolytic compounds. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 128–134. [Google Scholar] [CrossRef]
- Albert, K.; Pruessner, J.; Newhouse, P. Estradiol levels modulate brain activity and negative responses to psychosocial stress across the menstrual cycle. Psychoneuroendocrinology 2015, 59, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Rocca, W.A.; Grossardt, B.R.; Geda, Y.E.; Gostout, B.S.; Bower, J.H.; Maraganore, D.M.; de Andrade, M.; Melton, L.J. 3rd Long-term risk of depressive and anxiety symptoms after early bilateral oophorectomy. Menopause 2018, 25, 1275–1285. [Google Scholar] [CrossRef]
- Giannini, A.; Caretto, M.; Genazzani, A.R.; Simoncini, T. Neuroendocrine changes during menopausal transition. Endocrines 2021, 2, 405–416. [Google Scholar] [CrossRef]
- Pinna, G. Allopregnanolone, the neuromodulator turned therapeutic agent: Thank you, next? Front. Endocrinol. 2020, 11, 236. [Google Scholar] [CrossRef]
- Puga-Olguín, A.; Rodríguez-Landa, J.F.; Rovirosa-Hernández, M.J.; Germán-Ponciano, L.J.; Caba, M.; Meza, E.; Guillén-Ruiz, G.; Olmos-Vázquez, O.J. Long-term ovariectomy increases anxiety- and despair-like behaviors associated with lower Fos immunoreactivity in the lateral septal nucleus in rats. Behav. Brain Res. 2019, 360, 185–195. [Google Scholar] [CrossRef]
- Lovick, T.A. GABA in the female brain—Oestrous cycle-related changes in GABAergic function in the periaqueductal grey matter. Pharmacol. Biochem. Behav. 2008, 90, 43–50. [Google Scholar] [CrossRef]
- Rodríguez-Landa, J.F. Considerations of timing post-ovariectomy in mice and rats in studying anxiety- and depression-like behaviors associated with surgical menopause in women. Front. Behav. Neurosci. 2022, 16, 829274. [Google Scholar] [CrossRef]
- Paul, S.M.; Pinna, G.; Guidotti, A. Allopregnanolone: From molecular pathophysiology to therapeutics. A historical perspective. Neurobiol. Stress 2020, 12, 100215. [Google Scholar] [CrossRef]
- Georgieva, I.; Lepping, P.; Bozev, V.; Lickiewicz, J.; Pekara, J.; Wikman, S.; Lantta, T. Prevalence, new incidence, course, and risk factors of PTSD, depression, anxiety, and panic disorder during the COVID-19 pandemic in 11 countries. Healthcare 2021, 9, 664. [Google Scholar] [CrossRef]
- Zhukov, D.A.; Vinogradova, E.P. Neurosteroids and depression. Neurochem. J. 2021, 15, 240–246. [Google Scholar] [CrossRef]
- Perez-Caballero, L.; Torres-Sanchez, S.; Romero-López-Alberca, C.; González-Saiz, F.; Mico, J.A.; Berrocoso, E. Monoaminergic system and depression. Cell Tissue Res. 2019, 377, 107–113. [Google Scholar] [CrossRef]
- Rana, T.; Behl, T.; Sehgal, A.; Srivastava, P.; Bungau, S. Unfolding the role of BDNF as a biomarker for treatment of depression. J. Mol. Neurosci. 2021, 71, 2008–2021. [Google Scholar] [CrossRef]
- Zhou, C.; Zhong, J.; Zou, B.; Fang, L.; Chen, J.; Deng, X.; Zhang, L.; Zhao, X.; Qu, Z.; Lei, Y.; et al. Meta-analyses of comparative efficacy of antidepressant medications on peripheral BDNF concentration in patients with depression. PLoS ONE 2017, 12, e0172270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagud, M.; Nikolac Perkovic, M.; Vuksan-Cusa, B.; Maravic, A.; Svob Strac, D.; Mihaljevic Peles, A.; Zivkovic, M.; Kusevic, Z.; Pivac, N. A prospective, longitudinal study of platelet serotonin and plasma brain-derived neurotrophic factor concentrations in major depression: Effects of vortioxetine treatment. Psychopharmacol 2016, 233, 3259–3267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannestad, J.; Dellagioia, N.; Bloch, M. The effect of antidepressant medication treatment on serum levels of inflammatory cytokines: A meta-analysis. Neuropsychopharmacol 2011, 36, 2452. [Google Scholar] [CrossRef] [PubMed]
- Köhler, C.A.; Freitas, T.H.; Stubbs, B.; Maes, M.; Solmi, M.; Veronese, N.; de Andrade, N.Q.; Morris, G.; Fernandes, B.S.; Brunoni, A.R.; et al. Peripheral alterations in cytokine and chemokine levels after antidepressant drug treatment for major depressive disorder: Systematic review and meta-analysis. Mol. Neurobiol. 2018, 55, 4195–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Więdłocha, M.; Marcinowicz, P.; Krupa, R.; Janoska-Jaździk, M.; Janus, M.; Dębowska, W.; Mosiołek, A.; Waszkiewicz, N.; Szulc, A. Effect of antidepressant treatment on peripheral inflammation markers-A meta-analysis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 80, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Patatanian, E.; Nguyen, D.R. Brexanolone: A novel drug for the treatment of postpartum depression. J. Pharm. Pract. 2020, 0897190020979627, Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.; Krohn, H.; Richardson, E.; Kimmel, M.; Meltzer-Brody, S.A. Brexanolone treatment program at an academic medical center: Patient selection, 90-day posttreatment outcomes, and lessons learned. J. Acad. Consult. Liaison Psychiatry 2022, 63, 14–22. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Activity | Chrysin Treatment | Effects | Reference |
|---|---|---|---|
| Antioxidant | 20 mg/kg/30 days, p.o. | ↓ TBARS, lipid hydroperoxides, conjugated dienes tissue, circulatory levels ↑ SOD, CAT, GPx, Gsr activity, GSH, GSTs, vitamin C and vitamin E levels in ethanol-induced toxicity in rats | [22] |
| 50 mg/kg/12 days, i.p. | ↑ GHS levels and CAT and SOD activity in heart homogenate in male rats | [27] | |
| 30 and 60 mg/kg/28 days, p.o. | ↑ NO and GHS levels, GSHPx, CAT, and SOD activity in rat heart homogenate | [28] | |
| 60 mg/kg/28 days, p.o | ↑ 8-OHdG, TBARS levels ↓ GSH, CAT, NO levels | [29] | |
| 1 and 10 mg/kg/60 days, p.o. | ↑ SOD, CAT and GPx activity in PFC and HP of aged mice | [20] | |
| 1.25, 2.5, and 5 µM/30 min exposure | ↓ ROS formation in neuronal SH-SY5Y and microglial THP-1 cells in vitro | [30] | |
| 10, 30, and 100 mg/kg/44 days, p.o. | Protects against aluminum-induced oxidative stress by restored LPO levels and SOD and CAT activity in cortex and HP of male Swiss mice | [30] | |
| Anti-inflammatory | 7.50, 4.75, and 120.90 µM, 18 h exposure | ↓ NO, PGE2 and TNF-α biosynthesis in CLP-induced RAW 264.7 cells | [31] |
| 30 mg/kg/2 weeks, i.p. | ↓ ALT and AST activity ↓ TNF-α and IL-1β levels ↑ IL-10 and adiponectin in high-fat feeding mice | [32] | |
| 25 and 50 mg/kg/12 days, i.p. | ↓ NF-κB, iNOS, COX-2, and TNF-α expression in heart homogenate of DOX-induced cardiotoxicity mice | [27] | |
| 30 and 60 mg/kg/28 days, p.o. | ↑ PPAR-γ and TGF-β expression ↓ NF-κBp65 and IKK-β expression and TNF-α level in heart homogenate of isoproterenol-induced myocardial injury rats | [28,29] | |
| 5 and 20 mg/kg/28 days, p.o. | ↓ TNF-α, IL-1β and IL-6 levels in PFC and HP of chronically stressed mice | [18] | |
| 5 mg/kg/1 h before LP, i.p. | ↓ AST and TNF-α serum levels in septic mice survival | [33] | |
| 5 µM/24 h exposure | ↓ iNOS, IL-1β, and TNF-α expression in microglial THP-1 cells exposed to LPS | [30] | |
| GABAergic/BZD | 3 µM, 60 min exposure | Acts as competitive ligand for central BZD site in bovine cerebral cortical membranes in vitro | [15] |
| 13 µM, 60 min exposure | Acts as competitive ligand for peripheral BZD binding site in rat kidneys membranes in vitro | [15] | |
| 1 mg/kg, i.p. | Activates the GABAA/BZD receptor complex in male CF1 mice | [24] | |
| 1 mg/kg, i.p. | Activates the GABAA/BZD receptor complex in male Sprague Dawley rats | [17] | |
| 0.62 µM, 2 h exposure | Acts as competitive ligand for central BZD site in synaptosomal fractions of rat brain in vitro | [34] | |
| 10 and 30 µM, 30 s exposure | Modulates the activity of Cl− ion channel in the GABAA receptor expressed in Xenopus oocytes in vitro | [35] | |
| 2 mg/kg, i.p. | ↓ Anxiety-like behavior by modulating Cl− ion channel in the GABAA receptor of cycling female rats | [36] | |
| 2 mg/kg, i.p. | ↓ Depression-like behavior by modulating GABA-binding site in the GABAA receptor of ovariectomized female rats | [37] | |
| 0.5 µg/rat, i.h. | ↓ Anxiety-like behavior by modulating GABAA/BZD receptor complex in the dorsal hippocampus of cycling female rats | [14] | |
| Serotonergic | 5 and 20 mg/kg, p.o. | ↑ 5-HT levels and 5-HIAA/5-HT ratio in HP of chronic stressed mice | [18] |
| 20 mg/kg/28 days, p.o. | ↑ 5-HT levels in PFC and HP in female mice with hypothyroidism | [12] | |
| 50 mg/kg twice a day per 4 days, p.o. | ↑ 5-HT levels in the striatum of the rat brain | [38] | |
| 10 and 30 mg/kg/2 weeks, p.o. | ↑ 5-HT spinal levels ↓ 5-HIAA/5-HT ratio in male mice with experimental neuropathy | [39] | |
| 5 mg/kg/28 days, i.p. | ↓ 5-HT1A receptor expression in the dorsal raphe ↑ 5-HT1A and 5-HT2A in the hippocampus of male rats | [13] | |
| Dopaminergic | 10 mg/kg/28 days, p.o | ↑ DA striatal levels in mice | [19] |
| 50, 100 and 200 mg/kg/5 days, p.o. | ↑ DA levels in striatum of mice treated with 1-methyl-1,2,3,6-tetrahidropidine | [40] | |
| 20 mg/kg/28 days, p.o. | ↑ DA levels in PFC and HP in a hypothyroidism model in female mice | [12] | |
| Noradrenergic | 50, 100 and 150 mg/kg, i.p. | ↓ NE serum levels in rats with pain induced by formalin | [41] |
| 20 mg/kg/28 days, p.o. | No effects | [12] | |
| Anti-apoptotic | 25 and 50 mg/kg/12 days, i.p. | ↓ Bax, caspase-3, and cytochrome c activity ↑ Bcl-2 expression in rat heart tissue extract | [27] |
| 30 and 60 mg/kg/28 days, p.o. | ↑ Bcl-2 expression ↓ Bax and caspase-3 activity | [29] | |
| 5 and 20 mg/kg/28 days, p.o. | ↓ Caspase-3 and caspase-9 activity in HP and PFC of chronically stressed mice | [18] | |
| 25, 50 and 100 mg/kg/3 days, p.o. | ↓ Apoptotic index in cerebral cortex and HP of rats with traumatic brain injury | [42] | |
| Neuroendocrine | 5 and 20 mg/kg/28 days, p.o. | ↓ Corticosterone plasma levels in chronically stressed mice | [11] |
| 5 and 20 mg/kg/28 days, p.o. | ↓ CRH and ACTH in chronically stressed mice | [18] | |
| 50, 100 and 150 mg/kg, i.p | ↓ Corticosterone serum levels in rats with pain induced by formalin | [41] | |
| Neurotrophic | 5 and 20 mg/kg/28 days, p.o. | ↑ BDNF and NGF levels in PFC and HP in chronically stressed mice | [11] |
| 1 and 10 mg/kg/60 days, p.o. | ↑ BDNF levels in HP and PFC in aged mice | [20] | |
| 10 mg/kg/28 days, p.o. | ↑ BDNF and NGF levels in striatum in a Parkinson’s disease model in mice | [19] | |
| 20 mg/kg/28 days, p.o. | ↑ BDNF and NGF in HP and PFC in mice subjected to a hypothyroidism model | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Landa, J.F.; German-Ponciano, L.J.; Puga-Olguín, A.; Olmos-Vázquez, O.J. Pharmacological, Neurochemical, and Behavioral Mechanisms Underlying the Anxiolytic- and Antidepressant-like Effects of Flavonoid Chrysin. Molecules 2022, 27, 3551. https://doi.org/10.3390/molecules27113551
Rodríguez-Landa JF, German-Ponciano LJ, Puga-Olguín A, Olmos-Vázquez OJ. Pharmacological, Neurochemical, and Behavioral Mechanisms Underlying the Anxiolytic- and Antidepressant-like Effects of Flavonoid Chrysin. Molecules. 2022; 27(11):3551. https://doi.org/10.3390/molecules27113551
Chicago/Turabian StyleRodríguez-Landa, Juan Francisco, León Jesús German-Ponciano, Abraham Puga-Olguín, and Oscar Jerónimo Olmos-Vázquez. 2022. "Pharmacological, Neurochemical, and Behavioral Mechanisms Underlying the Anxiolytic- and Antidepressant-like Effects of Flavonoid Chrysin" Molecules 27, no. 11: 3551. https://doi.org/10.3390/molecules27113551
APA StyleRodríguez-Landa, J. F., German-Ponciano, L. J., Puga-Olguín, A., & Olmos-Vázquez, O. J. (2022). Pharmacological, Neurochemical, and Behavioral Mechanisms Underlying the Anxiolytic- and Antidepressant-like Effects of Flavonoid Chrysin. Molecules, 27(11), 3551. https://doi.org/10.3390/molecules27113551