
Hepatoprotective Polysaccharides from Geranium wilfordii: Purification, Structural Characterization, and Their Mechanism

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Preparation and Characterization of GCP
2.2. Structural Characterization of GCP, GNP, and GAP
2.2.1. Molecular Weights of GCP, GNP, and GAP
2.2.2. FT-IR Spectral Analysis
2.2.3. Monosaccharide Composition Analysis
2.2.4. Methylation Analysis
2.2.5. NMR Analysis
2D-NMR Analysis of GNP
NMR Analysis of GAP
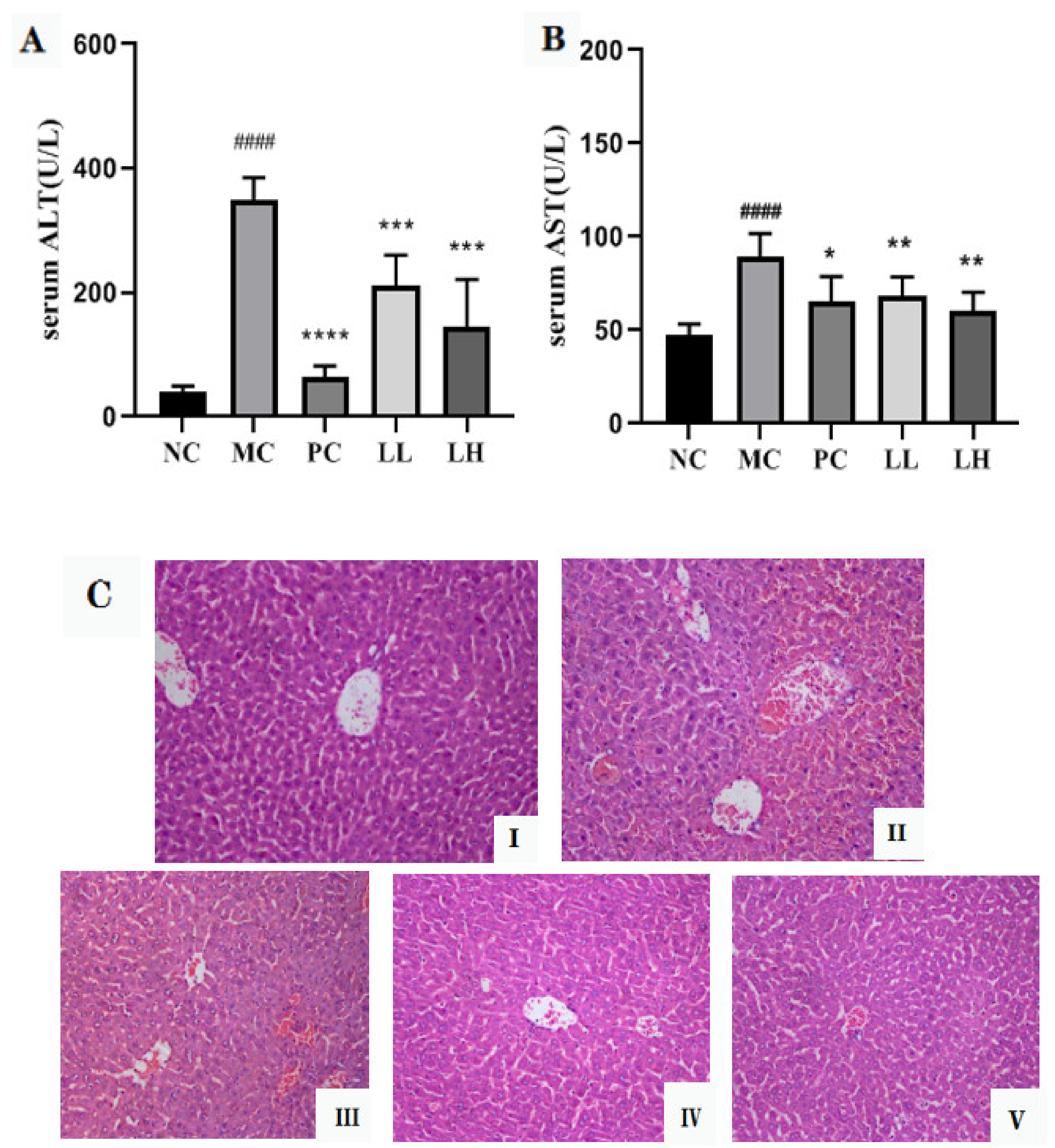
2.3. GCP Ameliorates CCl4-Induced Hepatotoxicity
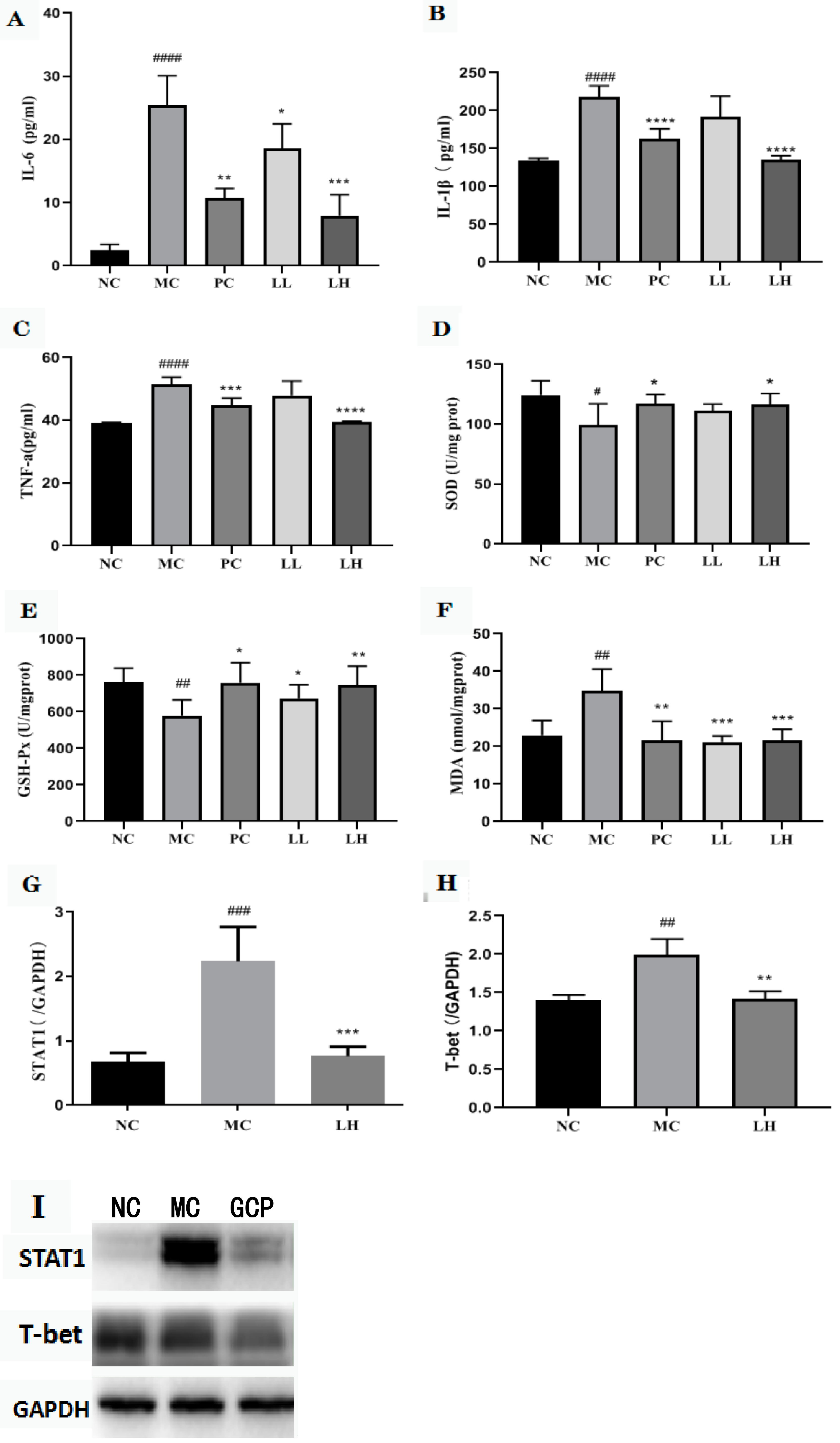
2.4. GCP Reduced the Expressions of Pro-Inflammatory Cytokines
2.5. GCP Inhibited the Oxidative Stress Level
2.6. Effect of GCP on STAT1 and T-Bet in CCl4-Induced Hepatitis
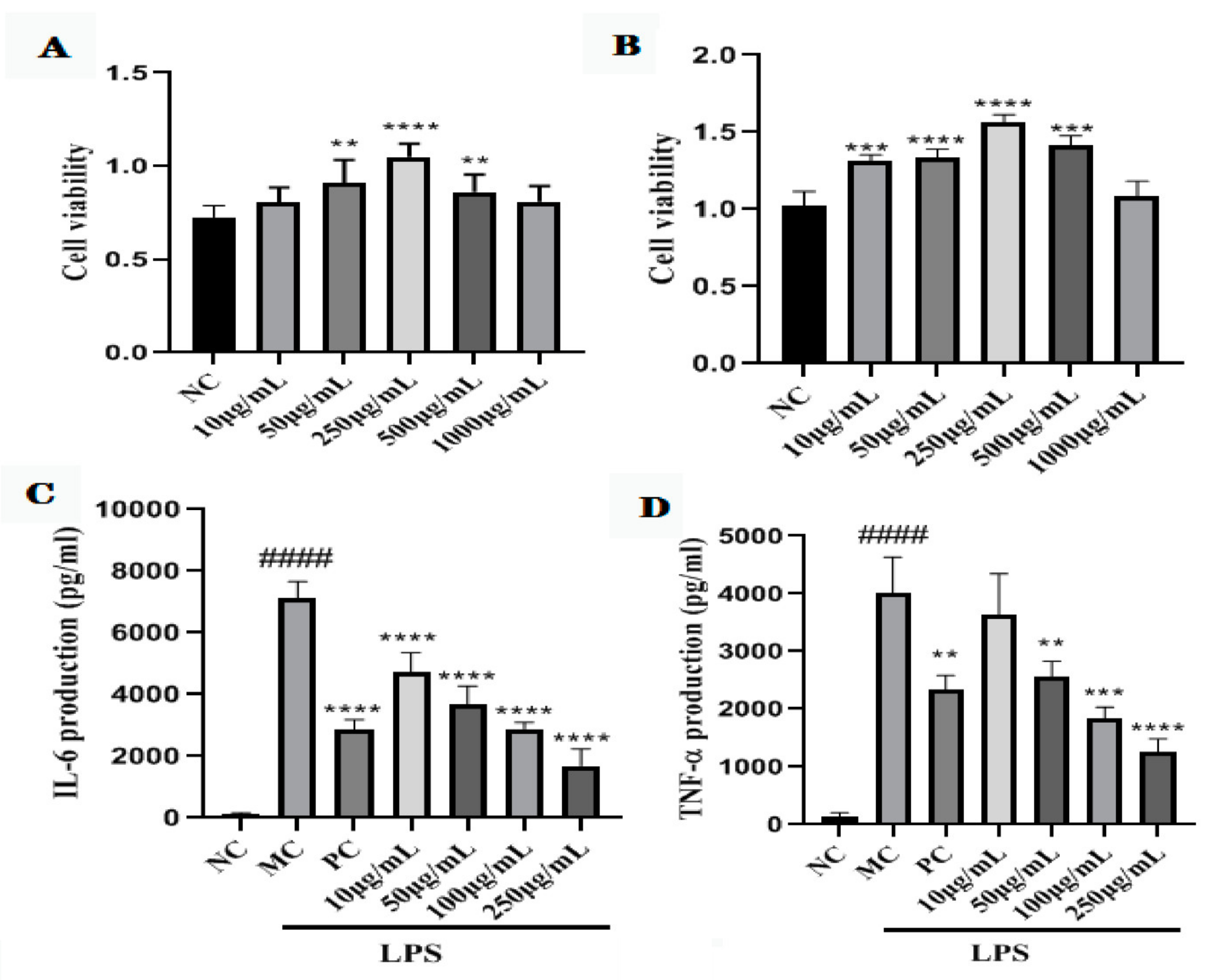
2.7. Cell Viability of GNP and GAP
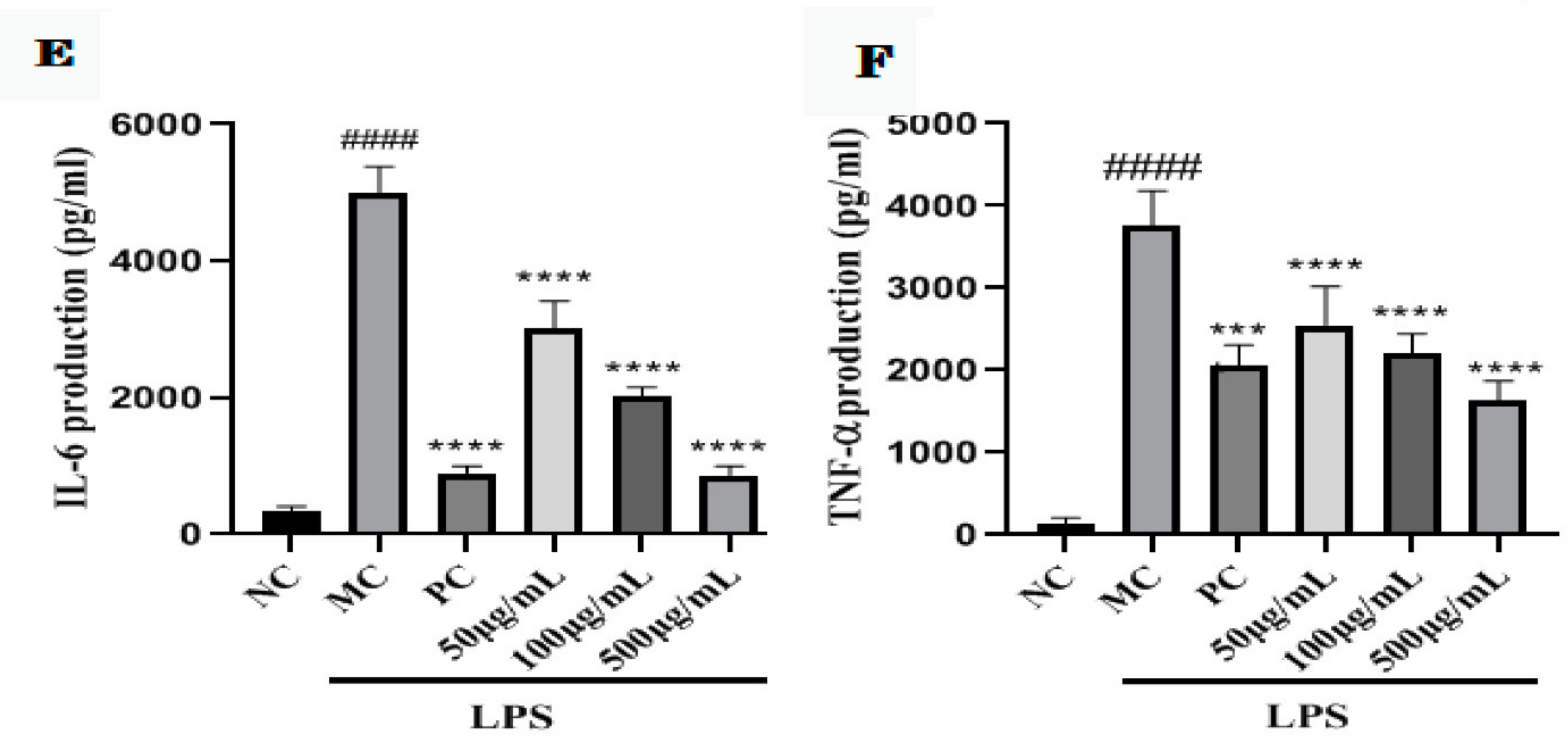
2.8. GNP and GAP Inhibit the LPS-Activated Production of Pro-Inflammatory Cytokines in RAW 264.7 Macrophages
3. Materials and Methods
3.1. Materials and Reagents
3.2. Extraction and Purification of the Plant Polysaccharide
3.3. Structural Characterization of GCP, GNP, and GAP
3.3.1. Determination of Polysaccharide and Protein Content
3.3.2. Molecular Weight Determination
3.3.3. Fourier Transform Infrared Spectroscopy (FT-IR)
3.3.4. Monosaccharide Composition
3.3.5. Methylation Analysis
3.3.6. NMR Analysis
3.4. Animal Experiments
3.4.1. Animal
3.4.2. Animal Grouping and Experimental Design
3.5. Determination of Biochemical Indicators
3.6. Histopathological Studies of Liver Tissues
3.7. Western Blotting
3.8. Cell Experiments In Vitro
3.8.1. Cell Culture
3.8.2. Cell Viability Assay
3.8.3. Determination of Pro-Inflammatory Cytokines Production
3.9. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Sample Availability
References
- Yang, J.Y.; Li, Y.; Wang, F.; Wu, C.F. Hepatoprotective effects of apple polyphenols on CCl4-induced acute liver damage in mice. J. Agric. Food Chem. 2010, 58, 6525–6531. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, J.; Lopez-Pelayo, H.; Michelena, J.; Jones, P.D.; Ortega, L.; Gines, P.; Caballeria, J.; Gual, A.; Bataller, R.; Lligona, A. Alcohol abstinence in patients surviving an episode of alcoholic hepatitis: Prediction and impact on long-term survival. Hepatology 2017, 66, 1842–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatology 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.X.; Zhang, H.M.; Chen, J. Status and progress of diagnosis and treatment of drug-induced liver injury. Mod. Med. Health 2018, 34, 1944–1948. [Google Scholar]
- Yang, X.B.; Yang, S.; Guo, Y.; Jiao, Y.; Zhao, Y. Compositional characterisation of soluble apple polysaccharides, and their antioxidant and hepatoprotective effects on acute CCl4-caused liver damage in mice. Food Chem. 2013, 138, 1256–1264. [Google Scholar] [CrossRef]
- Jaeschke, H.; McGill, M.R.; Ramachandran, A. Oxidant stress, mitochondria, and cell death mechanisms in drug-induced liver injury: Lessons learned from acetaminophen hepatotoxicity. Drug Metab. Rev. 2012, 44, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Bieghs, V.; Trautwein, C. The innate immune response during liver inflammation and metabolic disease. Trends Immunol. 2013, 34, 446–452. [Google Scholar] [CrossRef]
- Zeng, T.; Zhang, C.L.; Zhu, Z.P.; Yu, L.H.; Zhao, X.L.; Xie, K.Q. Diallyl trisulfide (DATS) effectively attenuated oxidative stress-mediated liver injury and hepatic mitochondrial dysfunction in acute ethanol-exposed mice. Toxicology 2008, 252, 86–91. [Google Scholar] [CrossRef]
- Huang, H.L.; Wang, Y.J.; Zhang, Q.Y.; Liu, B.; Wang, F.Y.; Li, J.J.; Run, Z.Z. Hepatoprotective effects of baicalein against CCl(4)-induced acute liver injury in mice. World J. Gastroenterol. 2012, 18, 6605–6613. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; Zhang, M.; Chen, Y.; Zhu, T.; Wang, J. Protective effect of eckol against acute hepatic injury induced by carbon tetrachloride in mice. Mar. Drugs. 2018, 16, 300. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Xiao, X.; Li, D.; Tun, S.; Wang, Y.; Velkov, T.; Tang, S.S. Chloroquine ameliorates carbon tetrachloride-induced acute liver injury in mice via the concomitant inhibition of inflammation and induction of apoptosis. Cell Death Dis. 2018, 9, 1164. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Tang, G.; Liu, P.; Zhao, C.J.; Liu, Q.; Li, H.B. Antioxidant activity and hepatoprotective effect of 10 medicinal herbs on CCl4-induced liver injury in mice. World J. Gastroenterol. 2020, 26, 5629–5645. [Google Scholar] [CrossRef] [PubMed]
- Ratha, P.; Chitra, L.; Ancy, I.; Kumaradhas, P.; Palvannan, T. New amino acid-Schiff base derived from s-allyl cysteine and methionine alleviates carbon tetrachloride-induced liver dysfunction. Biochimie 2017, 138, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, X.B.; Lu, X.S.; Wang, D.Y.; Zhao, Y. Protective effects of Keemun black tea polysaccharides on acute carbon tetrachloride-caused oxidative hepatotoxicity in mice. Food Chem. Toxicol. 2013, 58, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Wang, W.J.; Dong, C.J.; Huang, L.X.; Wang, H.; Li, C.; Nie, S.P.; Xie, M.Y. Protective effect offlavonoids from Cyclocarya paliurus leaves against carbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. 2018, 119, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Cheng, K.M.; Huang, H.Y.; Chao, P.Y.; Hwang, J.M.; Lee, H.H.; Cheng-You, L.; Yung-Wei, C.; Jer-Yuh, L. Hepatoprotective activity of Chhit-Chan-Than extract powder against carbon tetrachlorideinducedliver injury in rats. Food Drug Anal. 2014, 22, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.M.; Teng, H.; Zha, X.Q.; Pan, L.H.; Luo, J.P. Sulfated Laminaria japonica 24 polysaccharides inhibit macrophage foam cell formation. Int. J. Biol. Macromol. 2018, 111, 857–861. [Google Scholar] [CrossRef]
- Wang, C.; Shi, S.; Chen, Q.; Lin, S.; Wang, R.; Wang, S.; Chen, C. Antitumor and Immunomodulatory Activities of Ganoderma lucidum polysaccharides in Glioma-bearing rats. Integr. Cancer Ther. 2018, 17, 674–683. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Feng, L.; Su, J.; Cui, L.; Dan, L.; Yan, J.; Ding, C.; Tan, X.; Jia, X. Polysaccharides from Epimedium koreanum Nakai with immunomodulatory activity and inhibitory effect on tumor growth in LLC-bearing mice. J. Ethnopharmaco. 2017, 207, 8–18. [Google Scholar] [CrossRef]
- Chen, F.; Huang, G. Extraction and antioxidant activities of cushaw polysaccharide. Int. J. Biol. Macromol. 2018, 120, 1646–1649. [Google Scholar] [CrossRef]
- Jiang, Y.; Zi, W.; Pei, Z.; Liu, S. Characterization of polysaccharides and their antioxidant properties from Plumula nelumbinis. Saudi Pharm. J. 2018, 26, 656–664. [Google Scholar]
- Yüce, A.; Türk, G.; Çeribaşı, S.; Güvenç, M.; Çiftçi, M.; Sönmez, M.; Özer Kaya, Ş.; Çay, M.; Aksakal, M. Effectiveness of cinnamon (Cinnamomum zeylanicum) bark oil in the prevention of carbon tetrachloride-induced damages on the male reproductive system. Andrologia 2014, 46, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Chen, J.; Li, E.; Fan, Q.; Wang, D.; Li, P.; Li, X.; Chen, X.; Qiu, S.; Gao, Z.; et al. The comparison of antioxidative and hepatoprotective activities of Codonopsis pilosula polysaccharide (CP) and sulfated CP. Int. Immunopharmacol. 2015, 24, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.M.; Reisfeld, B.; Mayeno, A.N. Cytochromes P450: A structure-based summary of biotransformations using representative substrates. Drug Metab. Rev. 2008, 40, 1–288. [Google Scholar] [CrossRef] [PubMed]
- Yuetao, W.; Zhexiong, J. Research progress on chemical constituents of tannins in Geranium and their pharmacological activities. Heilongjiang Med. 2008, 46, 67–68. [Google Scholar]
- Behrooz, A.; Mahsa, K.; Babk, P.; Peyman, A. Clinical study on the treatment of chronic hepatitis B with Geranium Oral Liquid. J. Suzhou. First College. 1995, 15, 112. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin-phenol reagents. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Ren, Y.-Y.; Zhu, Z.-Y.; Sun, H.-Q.; Chen, L.J. Chemical Structural characterization and inhibition on α-glucosidase activity of acidic polysaccharide from Annona squamosa. Carbohyd. Polym. 2017, 174, 1–12. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, L.; Yang, Y.; Song, X.; Yu, Z. Optimization of ultrasound-assisted compound enzymatic extraction and characterization of polysaccharides from blackcurrant. Carbohyd. Polym. 2015, 117, 895–902. [Google Scholar]
- Yin, Z.H.; Zhang, W.; Zhang, J.J.; Liu, H.L.; Guo, Q.F.; Chen, L.; Wang, J.M.; Kang, W.Y. Two Novel Polysaccharides in Psoralea corylifolia L and anti-A549 Lung Cancer Cells Activity In Vitro. Molecules 2019, 24, 3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.F.; Sun, H.R.; Zhao, Y.F.; Chen, X.F.; Qi, Z.G. Chemical Constituents from the Defatted Seeds of Linum usitatissimum L. Nat. Prod. Res. Dev. 2009, 20, 60–62. [Google Scholar]
- Peng, D.; Wen, Y.; Bi, S.X.; Huang, C.H.; Yang, J.N.; Guo, Z.Y.; Huang, W.J.; Zhu, J.H.; Yu, R.M.; Song, L.Y. A new GlcNAc-containing polysaccharide from Morchella importuna fruiting bodies: Structural characterization and immunomodulatory activities in vitro and in vivo. Int. J. Biol. Macromol. 2021, 192, 1134–1149. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T. Isolation, Structural Identification and Biological Activity of Polysaccharides from Angelica Sinensis. Ph.D. Thesis, Huazhong Universtiy of Science and Technology, Wuhan, China, 2018. [Google Scholar]
- Wan, X.L.; Xin, J.; Wu, X.; Yang, X.; Lin, X.; Li, D.; Fu, Y.; Liu, Y.; Liu, X.; Lv, J.; et al. Structural characterization and antitumor activity against non-small cell lung cancer of polysaccharides from Sanghuangporus vaninii. Carbohyd. Polym. 2022, 276, 118798. [Google Scholar] [CrossRef] [PubMed]
- Kuang, M.T.; Li, J.Y.; Yang, X.B.; Yang, L.; Xu, J.Y.; Yan, S.; Lv, Y.F.; Ren, F.C.; Hu, J.M.; Zhou, J. Structural characterization and hypoglycemic effect via stimulating glucagon-like peptide-1 secretion of two polysaccharides from Dendrobium officinale. Carbohyd. Polym. 2022, 241, 116326. [Google Scholar] [CrossRef]
- Zou, Y.F.; Chen, X.F.; Malterud, K.E.; Rise, F.; Barsett, H.; Inngjerdingen, K.T.; Michaelsen, T.E.; Paulsen, B.S. Structural features and complement fixing activity of polysaccharides from Codonopsis pilosula Nannf. var. modesta L.T.Shen roots. Carbohyd. Polym. 2014, 113, 420–429. [Google Scholar] [CrossRef]
- Staub, A.M. Removal of protein-Sevag method. Carbohyd. Chem. 1965, 5, 5–6. [Google Scholar]
- Zhao, H.; Wang, Q.H.; Sun, Y.P.; Bingyou, Y.; Zhibin, W.; Guifang, C.; Yongzhou, G.; Weiguo, Z.; Zunpeng, S.; Xia, L.; et al. Purification, characterization and immunodulatory effects of Plantago depressa polysaccharides. Carbohyd. Polym. 2014, 112, 63–72. [Google Scholar] [CrossRef]
- Hall, M.B. Efficacy of reducing sugar and phenol-sulfuricacid assays for analysis of soluble carbohydrates in feed stuffs. Anim. Feed. Sci. Tech. 2013, 185, 94–100. [Google Scholar] [CrossRef]
- Silvério, S.; Moreira, S.; Milagres, A.M.; Macedo, E.A.; Teixeira, J.; Mussatto, S.I. Interference of some aqueous twophase system phase-forming components in protein determination by the Bradford method. Anal. Biochem. 2012, 421, 719–724. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Huang, Q.L.; Ling, C.Q. Water-soluble yeastβ’glucan fractions with different molecular weights: Extraction and separation by acidolysis assisted-size exclusion chromatography and their association with proliferative activity. Int. J. Biol. Macromol. 2018, 123, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, B.; Xiao, J.; Huang, Q.; Li, C.; Fu, X. Physicochemical, functional, and biological properties of water-soluble polysaccharides from Rosa roxburghii Tratt fruit. Food Chem. 2018, 249, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Salvador, L.D.; Suganuma, T.; Kitahara, K.; Tanoue, H.; Ichiki, M. Monosaccharide composition of sweetpotato fiber and cell wall polysaccharides from sweetpotato, cassava, and potato analyzed by the high-performance anion exchange chromatography with pulsed amperometric detection method. J. Agric. Food Chem. 2000, 48, 3448–3454. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.B.; Liang, H.P.; Li, H.M.; Yuan, R.N.; Sun, J.; Zhang, L.L.; Han, M.H.; Wu, Y. Isolation, purification, characterization and antioxidant activity of polysaccharides from the stem barks of Acanthopanax leucorrhizus. Carbohyd. Polym. 2018, 196, 359–367. [Google Scholar] [CrossRef]
- Kumagal, K.; Tabu, K.; Sasakl, F.; Takami, Y.; Morianga, Y.; Mawatari, S.; Hashimoto, S.; Tanoue, S.; Hanmura, S.; Tamai, T.; et al. Glycoprotein nonmetastatic melanoma B (Gpnmb)-positive macrophages contribute to the balance betweenfibrosis andfibrolysis during the repair of acute liver injury in mice. PLoS ONE 2015, 23, 1–19. [Google Scholar]
- Liu, F.C.; Hwang, T.L.; Lau, Y.T.; Yu, H.P. Mechanism of salutary effects of Astringinin on rodent hepatic injury following trauma-hemorrhage: Akt-dependent heme oxygenase-1 signaling pathways. PLoS ONE 2011, 11, 1–7. [Google Scholar] [CrossRef]
- Zeng, B.Y.; Su, M.H.; Chen, Q.X.; Chang, Q.; Wang, W.; Li, H.H. Antioxidant and hepatoprotective activities of polysaccharides from Anoectochilus roxburghii. Carbohydr. Polym. 2016, 153, 391–398. [Google Scholar] [CrossRef]
- Yang, L.; Liu, R.H.; Fang, Y.W.; He, J.W. Anti-inflammatory effect of phenylpropanoids from Dendropanax dentiger in TNF-α-induced MH7A cells via inhibition of NF-κB, Akt and JNK signaling pathways. Int. Immunopharmacol. 2021, 94, 107463. [Google Scholar] [CrossRef]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef]
- Yuan, Y.; Che, L.H.; Qi, C.; Meng, Z.L. Protective effects of polysaccharides on hepatic injury: A review. Int. J. Biol. Macromol. 2019, 141, 822–830. [Google Scholar] [CrossRef]
- Xianfei, X.; Guolin, Z.; Chenghai, L. Antitumor and immunomodulatory activities of a water-soluble polysaccharide from Chaenomeles speciosa. Carbohyd. Polym. 2015, 132, 323–329. [Google Scholar]
- Xing, X.; Cui, S.W.; Nie, S.; Phillips, G.O.; Goff, H.D.; Wang, Q. Study on Dendrobium officinale O -acetyl-glucomannan (Dendronan ) Part II. Fine structures of O -acetylated residues. Carbohyd. Polym. 2015, 117, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Ning, W.; Yu, X.; Tan, H.Y.; Li, S.; Feng, Y.B. Molecular Mechanisms Involved in Oxidative Stress-Associated Liver Injury Induced by Chinese Herbal Medicine: An Experimental Evidence-Based Literature Review and Network Pharmacology Study. Int. J. Biol. Macromol. 2018, 19(9), 2745. [Google Scholar]
- Hai-Xia, W.; Man, L.; Shun-Yan, W.; Jing-Jing, L.; Hong-Lin, H.; Wen, G.; Yun-Sheng, Y.; Jin, G. Immune mechanisms of Concanavalin A model of autoimmune hepatitis. World J. Gastroenterol. 2012, 18(2), 119. [Google Scholar]
- Afkarian, M.; Sedy, J.R.; Yang, J.; Jacobson, N.G.; Cereb, N.; Yang, S.Y.; Murphy, T.L.; Murphy, M.K. T-bet is a STAT1-induced regulator of IL-12R expression in naive CD4 T cells. Nat. Immunol. 2002, 3(6), 549–557. [Google Scholar] [CrossRef] [PubMed]
- Siebler, J.; Wirtz, S.; Klein, S.; Martina, P.; Manfred, B.; Peter, R.G.; Markus, F.N. A key pathogenic role for the STAT1/T-bet signaling pathway in T-cell–mediated liver inflammation. Hepatology 2003, 38, 1573–1580. [Google Scholar]
- Liu, S.; Yang, T.; Ming, T.W.; Gaun, T.K.W.; Zhou, T.; Wang, S.; Ye, B.G. Isosteroid alkaloids with different chemical structures from Fritillariae cirrhosae bulbus alleviate LPS-induced inflammatory response in RAW 264.7 cells by MAPK signaling pathway. Int. Immunopharmacol. 2020, 78, 106047. [Google Scholar] [CrossRef]
- Shuping, S.; Yunyan, D.; Chuanliu, Y.; Xiaoguo, S.; Rui, W.; Rongping, X.; Xiaoping, Z. Water-separated part of Chloranthus serratus alleviates lipopolysaccharide- induced RAW264.7 cell injury mainly by regulating the MAPK and Nrf2/HO-1 inflammatory pathways. BMC Complement. Altern. Med. 2019, 19, 1–13. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mouse Liver Index% ± SD | Mouse Spleen Index% ± SD |
|---|---|---|
| Normal group (NC) | 4.33 ± 0.27 | 0.43 ± 0.08 |
| Model group (MC) | 5.06 ± 0.14 #### | 0.68 ± 0.9 ### |
| Positive control group (PC) | 4.57 ± 0.34 | 0.51 ± 0.08 ** |
| Low-dose group LL | 4.37 ± 0.39 *** | 0.63 ± 0.07 * |
| High-dose group LH | 3.34 ± 0.27 **** | 0.49 ± 0.04 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.-Y.; Xie, Y.-Q.; Zhang, P.; Zhou, Q.; Khan, A.; Zhou, Z.-H.; Xia, X.-S.; Liu, L. Hepatoprotective Polysaccharides from Geranium wilfordii: Purification, Structural Characterization, and Their Mechanism. Molecules 2022, 27, 3602. https://doi.org/10.3390/molecules27113602
Feng J-Y, Xie Y-Q, Zhang P, Zhou Q, Khan A, Zhou Z-H, Xia X-S, Liu L. Hepatoprotective Polysaccharides from Geranium wilfordii: Purification, Structural Characterization, and Their Mechanism. Molecules. 2022; 27(11):3602. https://doi.org/10.3390/molecules27113602
Chicago/Turabian StyleFeng, Jia-Yi, Yan-Qing Xie, Peng Zhang, Qian Zhou, Afsar Khan, Zhi-Hong Zhou, Xian-Song Xia, and Lu Liu. 2022. "Hepatoprotective Polysaccharides from Geranium wilfordii: Purification, Structural Characterization, and Their Mechanism" Molecules 27, no. 11: 3602. https://doi.org/10.3390/molecules27113602
APA StyleFeng, J. -Y., Xie, Y. -Q., Zhang, P., Zhou, Q., Khan, A., Zhou, Z. -H., Xia, X. -S., & Liu, L. (2022). Hepatoprotective Polysaccharides from Geranium wilfordii: Purification, Structural Characterization, and Their Mechanism. Molecules, 27(11), 3602. https://doi.org/10.3390/molecules27113602