
Antifungal Volatilomes Mediated Defense Mechanism against Fusarium oxysporum f. sp. lycopersici, the Incitant of Tomato Wilt

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Volatilomes Formulation
2.2. Efficacy of Volatilomes Immobilized Vermiculite Balls against Pathogens
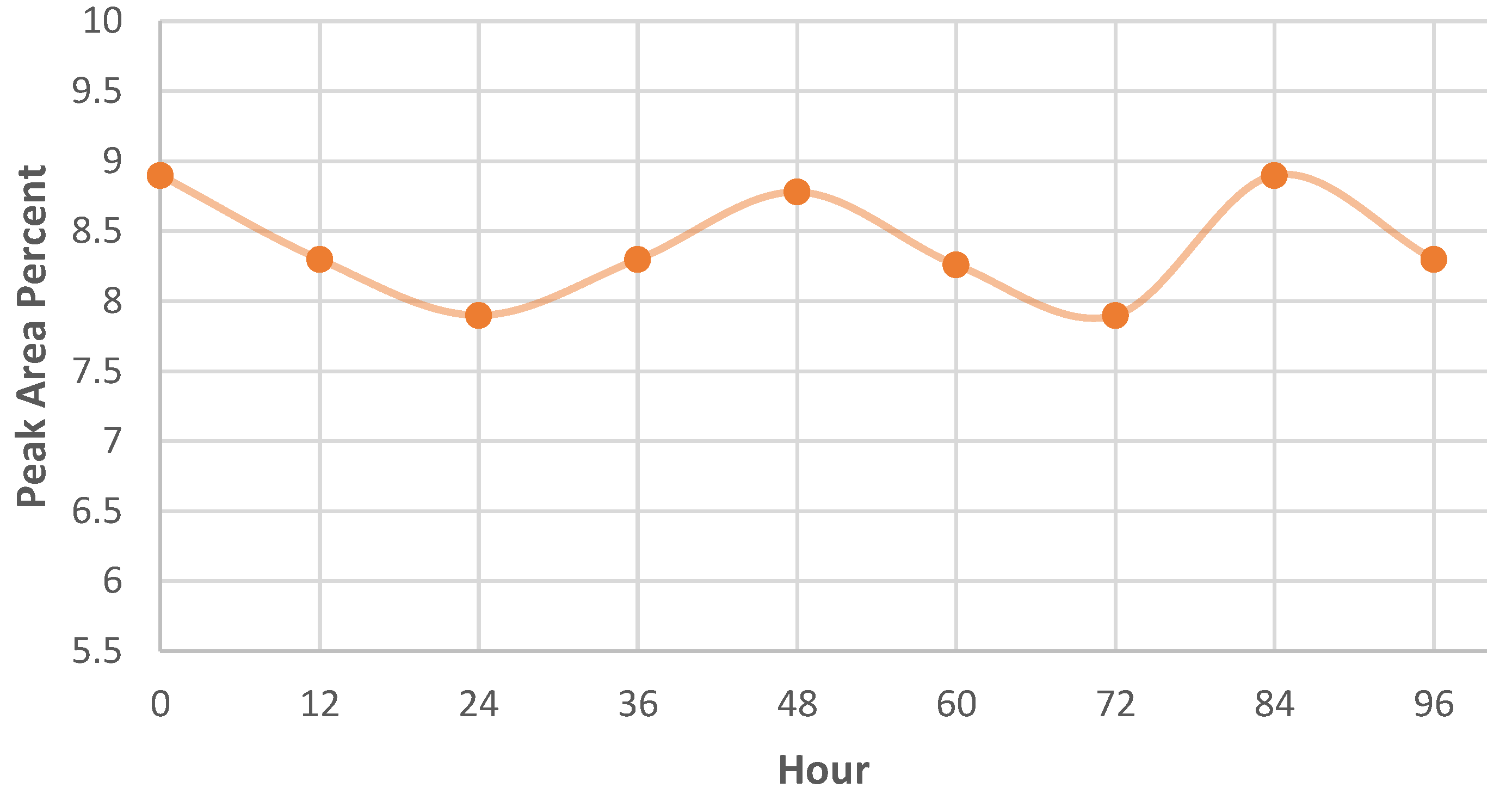
2.3. GC MS Analysis of M. spicata Volatilomes Immobilized Vermiculite Ball
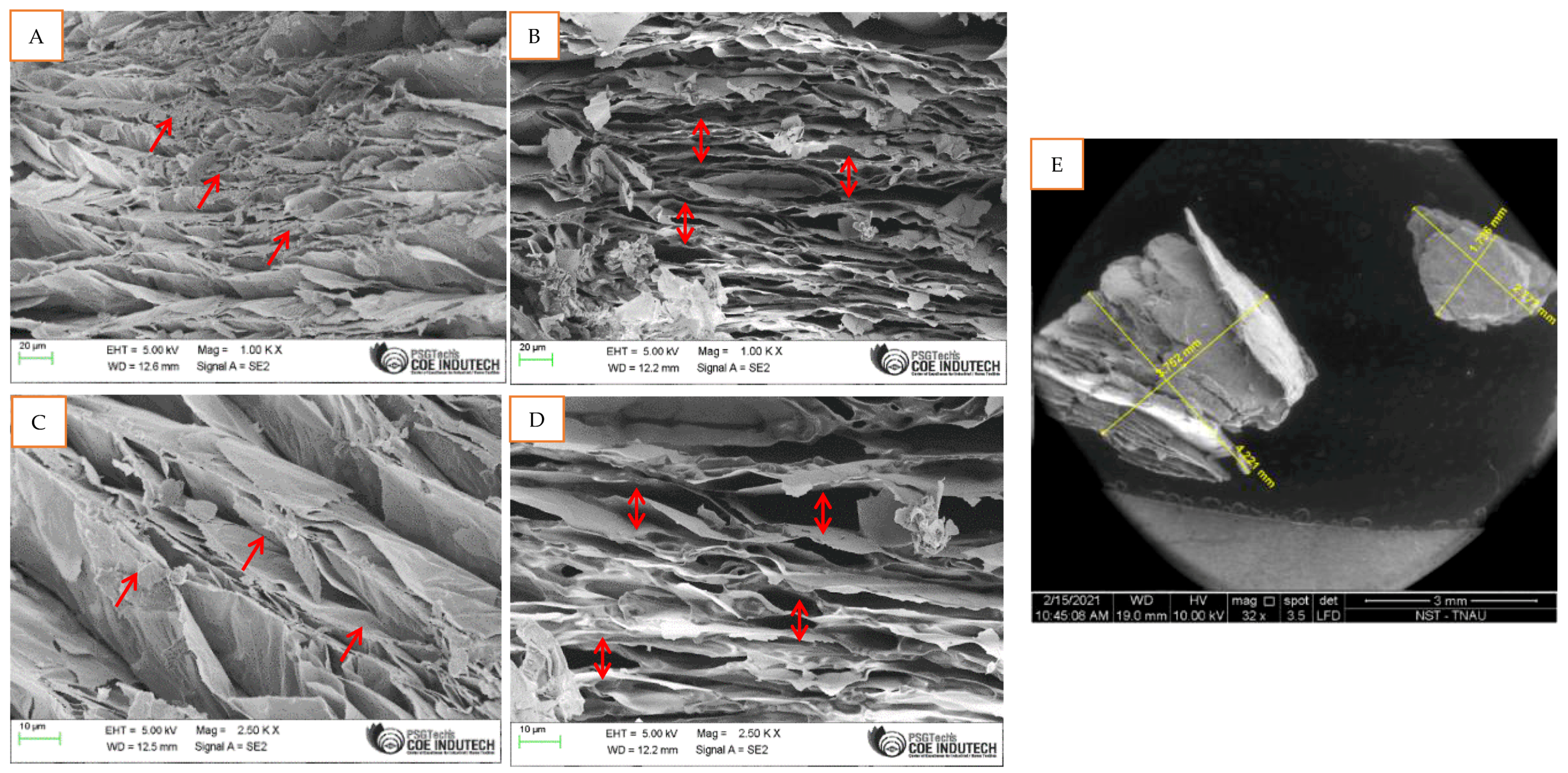
2.4. SEM Imaging of M. spicata Volatilomes Immobilized Vermiculite Ball
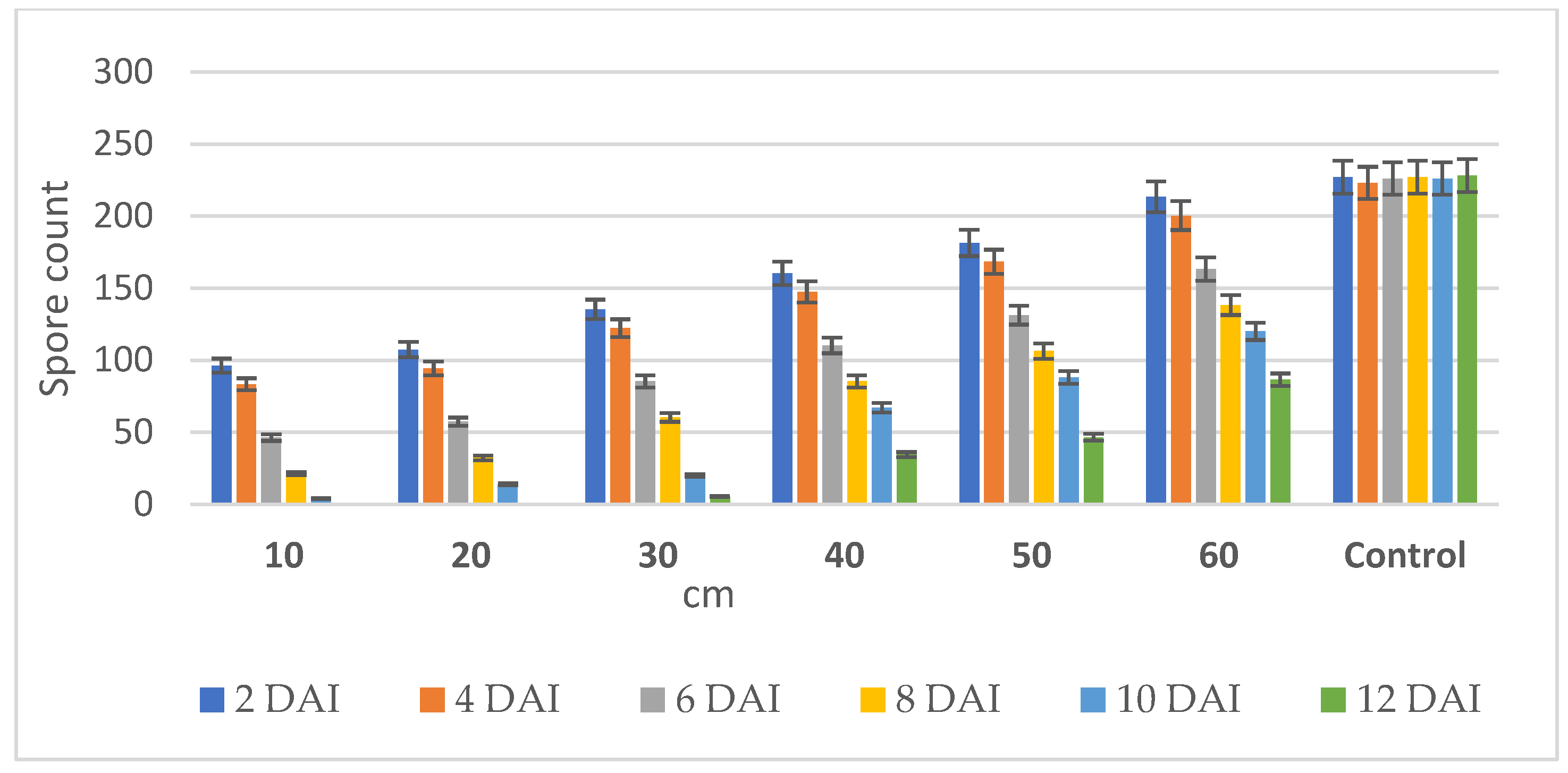
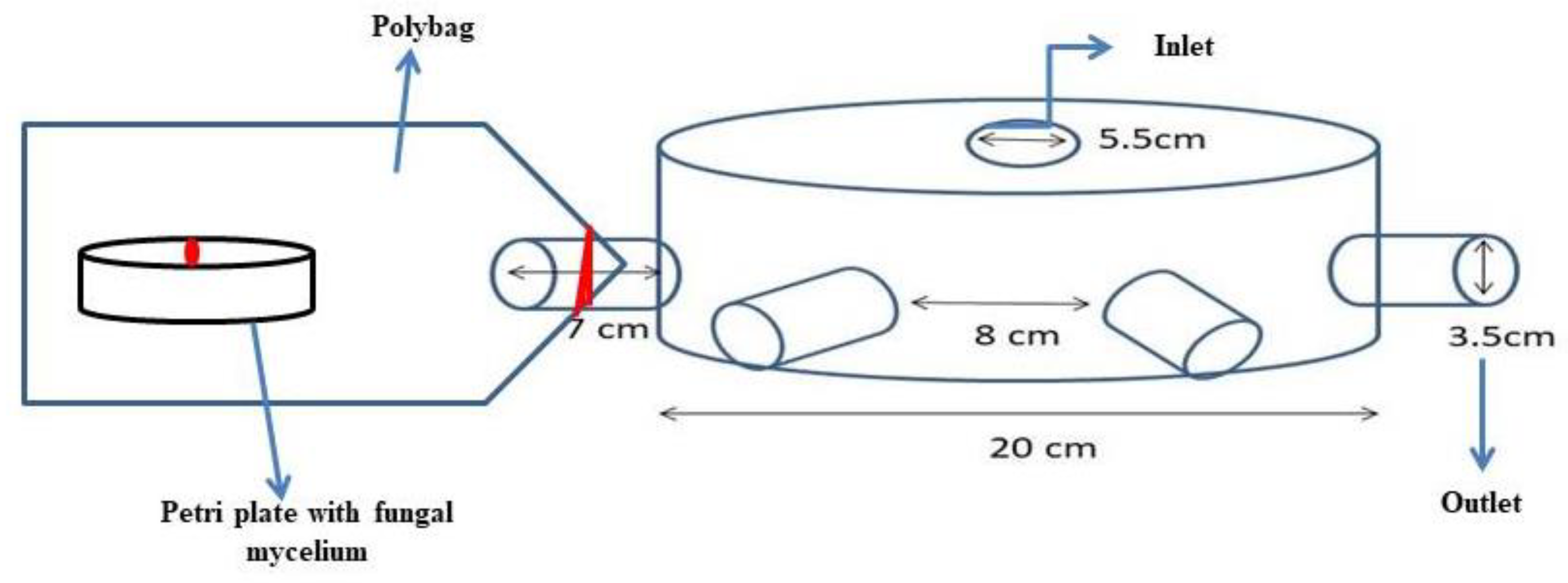
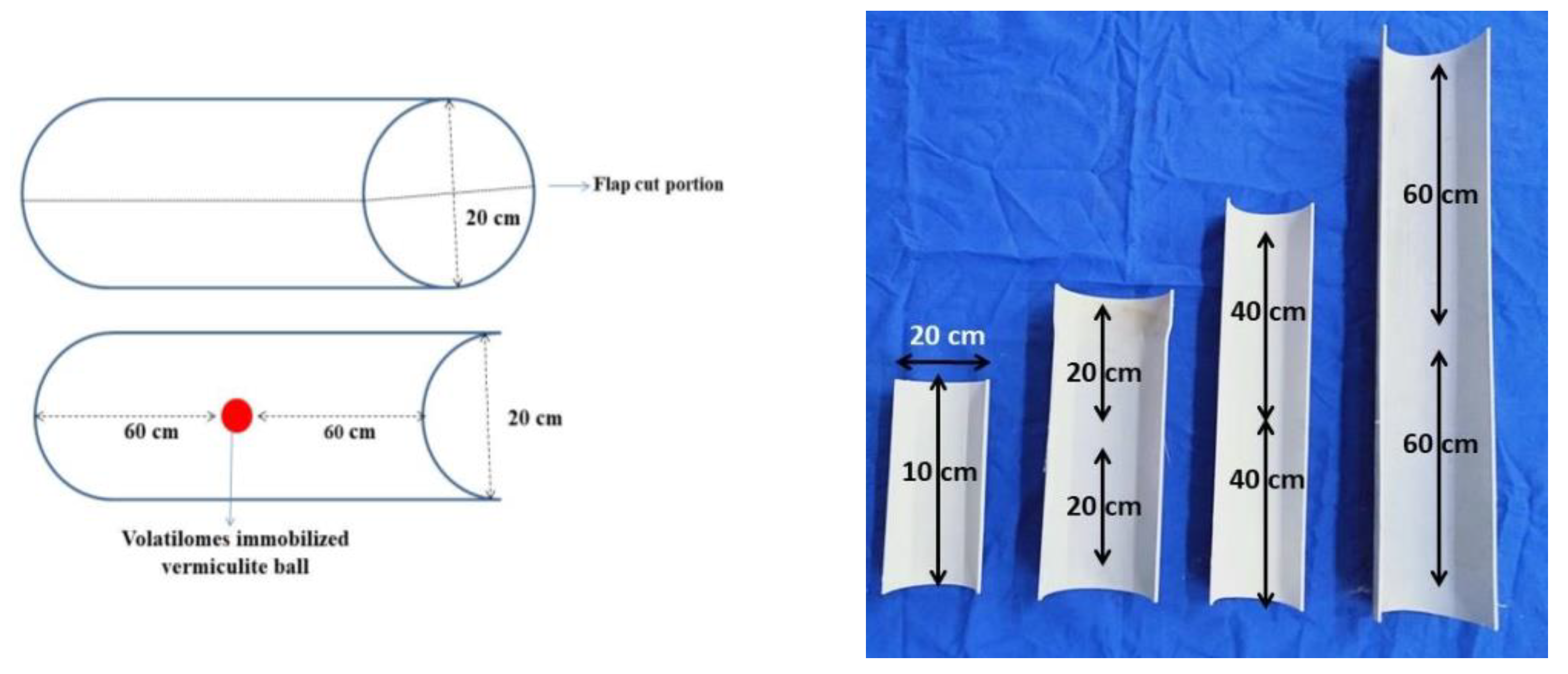
2.5. Testing the Distance Traveled by the Volatilomes Immobilized Vermiculite Ball
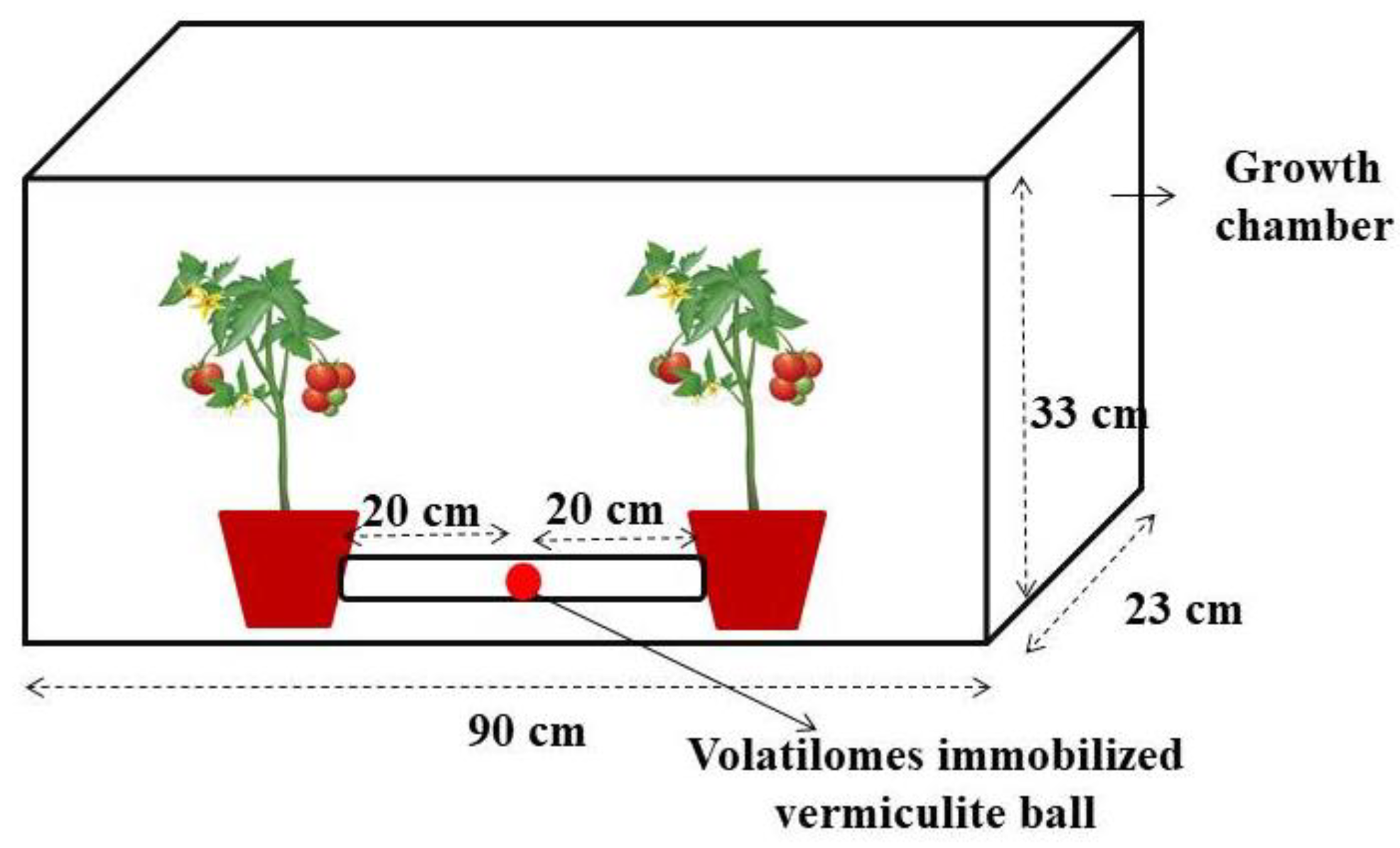
2.6. Effect of Volatilomes Immobilized Vermiculite Balls under Glasshouse Condition
2.7. Defense Genes Expression
3. Discussion
4. Materials and Methods
4.1. Plant and Microbial Volatilomes
4.2. Immobilization of Eluted and Trapped Volatile Compounds
4.3. Testing the Volatilomes Immobilized Vermiculite Balls against Pathogens
4.4. Testing the Distance on Volatilomes Travel
4.5. Effect of Volatilomes Immobilized Vermiculite Balls Tested in Growth Chamber
4.6. Defense Gene Expression Studies
4.6.1. RNA Extraction, cDNA Conversion
4.6.2. Quantification of Defense Related Genes Expression
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vignesh, K.; Rajamohan, K.; Balabaskar, P.; Anandan, R.; Udhayakumar, R. Survey on the incidence of fusarium wilt of tomato incited by Fusarium oxysporum f. sp. lycopersici (FOL) in major tomato growing areas of krishnagiri district. Plant Arch. 2021, 21, 2369–2376. [Google Scholar]
- Srinivas, C.; Devi, D.N.; Murthy, K.N.; Mohan, C.D.; Lakshmeesha, T.; Singh, B.; Kalagatur, N.K.; Niranjana, S.; Hashem, A.; Alqarawi, A.A. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity—A Review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Yucel, S.; Ozarslandan, A.; Colak, A.; Ay, T.; Can, C. Effect of solarization and fumigant applications on soilborne pathogens and root-knot nematodes in greenhouse-grown tomato in turkey. Phytoparasitica 2007, 35, 450–456. [Google Scholar] [CrossRef]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Katan, J. Diseases caused by soilborne pathogens: Biology, management and challenges. J. Plant Pathol. 2017, 99, 305–315. [Google Scholar]
- Larkin, R.P.; Brewer, M.T. Effects of crop rotation and biocontrol amendments on rhizoctonia disease of potato and soil microbial communities. Agriculture 2020, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, W.; Achouri, M.; Chtaina, N. Rhizoctonia solani-green beans pathosystem uncover bio control efficacy of Trichoderma spp. Adv. Plants Agric. Res. 2019, 9, 96–99. [Google Scholar]
- Ameye, M.; Audenaert, K.; De Zutter, N.; Steppe, K.; Van Meulebroek, L.; Vanhaecke, L.; De Vleesschauwer, D.; Haesaert, G.; Smagghe, G. Priming of wheat with the green leaf volatile z-3-hexenyl acetate enhances defense against Fusarium graminearum but boosts deoxynivalenol production. Plant Physiol. 2015, 167, 1671–1684. [Google Scholar] [CrossRef] [Green Version]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting plant volatile organic compounds (VOCs) in agriculture to improve sustainable defense strategies and productivity of crops. Front. Plant Sci. 2019, 10, 264. [Google Scholar] [CrossRef]
- Jamiołkowska, A. Natural compounds as elicitors of plant resistance against diseases and new biocontrol strategies. Agronomy 2020, 10, 173. [Google Scholar] [CrossRef] [Green Version]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7, 11050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamova, T.; Hradecky, J.; Panek, M. Volatile organic compounds (VOCs) from wood and wood-based panels: Methods for evaluation, potential health risks, and mitigation. Polymers 2020, 12, 2289. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.; Schuurink, R.C.; Bleeker, P.M.; Schiestl, F. The role of volatiles in plant communication. Plant J. 2019, 100, 892–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; De Jager, V.; Zühlke, D.; Wolff, C.; Bernhardt, J.; Cankar, K.; Beekwilder, J.; Van Ijcken, W.; Sleutels, F.; De Boer, W. Fungal volatile compounds induce production of the secondary metabolite sodorifen in Serratia plymuthica pri-2c. Sci. Rep. 2017, 7, 862. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D.; Hossain, O.; Weinhold, A.; Rose, U.S.; Wei, Q. Trends and applications in plant volatile sampling and analysis. Plant J. 2021, 106, 314–325. [Google Scholar] [CrossRef]
- Akter, A.; Neela, F.; Khan, M.; Islam, M.; Alam, M. Screening of ethanol, petroleum ether and chloroform extracts of medicinal plants, Lawsonia inermis L. and Mimosa pudica L. For antibacterial activity. Indian J. Pharm. Sci. 2010, 72, 388. [Google Scholar]
- Khan, M.; Neela, F.; Aktar, A.; Rahman, M.; Alam, M. Antibacterial activity of Achyranthes aspera L.: An in vitro study. J. Environ. Sci. Nat. Resour. 2009, 2, 45–48. [Google Scholar]
- Soylu, E.M.; Soylu, S.; Kurt, S. Antimicrobial activities of the essential oils of various plants against tomato late blight disease agent Phytophthora infestans. Mycopathologia 2006, 161, 119–128. [Google Scholar] [CrossRef]
- Prsic, J.; Ongena, M. Elicitors of plant immunity triggered by beneficial bacteria. Front. Plant Sci. 2020, 11, 1675. [Google Scholar] [CrossRef]
- Jangir, M.; Pathak, R.; Sharma, S.; Sharma, S. Biocontrol mechanisms of Bacillus sp., isolated from tomato rhizosphere, against Fusarium oxysporum f. sp. lycopersici. Biol. Control 2018, 123, 60–70. [Google Scholar] [CrossRef]
- Pickett, J.A.; Khan, Z.R. Plant volatile-mediated signalling and its application in agriculture: Successes and challenges. New Phytol. 2016, 212, 856–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, W.; Yuan, J.; Ling, N.; Huang, Q.; Shen, Q. Production of volatile organic compounds by an antagonistic strain Paenibacillus polymyxa wr-2 in the presence of root exudates and organic fertilizer and their antifungal activity against Fusarium oxysporum f. sp. niveum. Biol. Control 2015, 80, 89–95. [Google Scholar] [CrossRef]
- Schalchli, H.; Tortella, G.; Rubilar, O.; Parra, L.; Hormazabal, E.; Quiroz, A. Fungal volatiles: An environmentally friendly tool to control pathogenic microorganisms in plants. Crit. Rev. Biotechnol. 2016, 36, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Li, B. Antibacterial vermiculite nano-material. J. Miner. Mater. Charact. Eng. 2002, 1, 61–68. [Google Scholar]
- Xie, Y.; Wang, A. Effects of modified vermiculite on water absorbency and swelling behavior of chitosan-g-poly (acrylic acid)/vermiculite superabsorbent composite. J. Compos. Mater. 2009, 43, 2401–2417. [Google Scholar] [CrossRef]
- Barabaszova, K.C.; Holesova, S.; Bily, M.; Hundakova, M. Cuo and cuo/vermiculite based nanoparticles in antibacterial PVAC nanocomposites. J. Inorg. Organomet. Polym. Mater. 2020, 30, 4218–4227. [Google Scholar] [CrossRef]
- Cech Barabaszova, K.; Holesova, S.; Sulcova, K.; Hundakova, M.; Thomasova, B. Effects of ultrasound on zinc oxide/vermiculite/chlorhexidine nanocomposite preparation and their antibacterial activity. Nanomaterials 2019, 9, 1309. [Google Scholar] [CrossRef] [Green Version]
- Conchou, L.; Lucas, P.; Meslin, C.; Proffit, M.; Staudt, M.; Renou, M. Insect odorscapes: From plant volatiles to natural olfactory scenes. Front. Physiol. 2019, 10, 972. [Google Scholar] [CrossRef]
- Afify, A.; Potter, C.J. Insect repellents mediate species-specific olfactory behaviours in mosquitoes. Malar. J. 2020, 19, 127. [Google Scholar] [CrossRef] [Green Version]
- Cirlini, M.; Mena, P.; Tassotti, M.; Herrlinger, K.A.; Nieman, K.M.; Dall’Asta, C.; Del Rio, D. Phenolic and volatile composition of a dry spearmint (Mentha spicata L.) extract. Molecules 2016, 21, 1007. [Google Scholar] [CrossRef] [Green Version]
- Muttalib, S.A.; Edros, R.; Azah, N.; Kutty, R.V. A review: The extraction of active compound from Cymbopogon sp. and its potential for medicinal applications. Int. J. Eng. Technol. Sci. 2018, 5, 82–98. [Google Scholar] [CrossRef]
- Kalleli, F.; Ghassen, A.; Salem, I.B.; Boughalleb-M’hamdi, N.; M’Hamdi, M. Essential oil from fennel seeds (Foeniculum vulgare) reduces fusarium wilt of tomato (Solanum lycopersicon). Phytopathol. Mediterr. 2020, 59, 63–76. [Google Scholar]
- Choi, H.K.; Song, G.C.; Yi, H.S.; Ryu, C.M. Field evaluation of the bacterial volatile derivative 3-pentanol in priming for induced resistance in pepper. J. Chem. Ecol. 2014, 40, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhang, H.; Yao, J.; Wang, X.; Xu, J.; Han, Q.; Wei, G.; Huang, L.; Kang, Z. Characterization of non-host resistance in broad bean to the wheat stripe rust pathogen. BMC Plant Biol. 2012, 12, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naznin, H.A.; Kiyohara, D.; Kimura, M.; Miyazawa, M.; Shimizu, M.; Hyakumachi, M. Systemic resistance induced by volatile organic compounds emitted by plant growth-promoting fungi in Arabidopsis thaliana. PLoS ONE 2014, 9, e86882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kottb, M.; Gigolashvili, T.; Grobkinsky, D.K.; Piechulla, B. Trichoderma volatiles effecting Arabidopsis: From inhibition to protection against phytopathogenic fungi. Front. Microbiol. 2015, 6, 995. [Google Scholar] [CrossRef] [Green Version]
- Aamir, M.; Singh, V.K.; Dubey, M.K.; Kashyap, S.P.; Zehra, A.; Upadhyay, R.S.; Singh, S. Structural and functional dissection of differentially expressed tomato WRKY transcripts in host defense response against the vascular wilt pathogen (Fusarium oxysporum f. sp. lycopersici). PLoS ONE 2018, 13, e0193922. [Google Scholar] [CrossRef] [Green Version]
- Ojola, P.O.; Nyaboga, E.N.; Njiru, P.N.; Orinda, G. Overexpression of rice thaumatin-like protein (OSTLP) gene in transgenic cassava results in enhanced tolerance to Colletotrichum gloeosporioides f. sp. manihotis. J. Genet. Eng. Biotechnol. 2018, 16, 125–131. [Google Scholar] [CrossRef]
- Praveen, T.; Krishnamoorthy, A.S.; Nakkeeran, S.; Sivakumar, U.; Amirtham, D. Antifungal volatiles from medicinal herbs suppress Fusarium oxysporum f. sp. lycopersici. J. Entomol. Zool. Stud. 2021, 9, 1083. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Shoot Length(cm) | Root Length (cm) | No of Branches | Percent Wilt Incidence | Percent Reduction over Control |
|---|---|---|---|---|---|
| Vermiculite + M. spicata in pathogen inoculated | 31.83 | 22.67 | 9.33 | 8.33 c (16.78) | 91.67 a (73.22) |
| Uninoculated control | 25.83 | 15.87 | 7.33 | 83.33 b (65.91) | 16.67 b (24.09) |
| Inoculated control | 21.67 | 3.10 | 3.33 | 100.00 a (90.00) | 0.00 c (0.02) |
| Defense Gene | Forward and Reverse Primer Sequence | Reference |
|---|---|---|
| SlPR 1 | Forward: 5′-ACGTCTTGGTTGTGCTAGGG-3′ | [32] |
| Reverse: 5′-TCAAAAGCCGGTTGATTTTC-3′ | ||
| SlLOX | Forward: 5′-TGGGATTAAACTGCCAGACC-3′ | |
| Reverse: 5′-GGCATCGGAAATTTGAGAAA-3′ | ||
| SlWRKY | Forward:5′-TCTCGATCTGACCAGGTTCC-3′ | |
| Reverse: 5′-TTGCCGTCTTCGTTCTCTTT-3′ | ||
| SlTLP | Forward:5′-CCATCTTTGCTTCCCACATT-3′ | |
| Reverse: 5′-ATCGGTTTACCTGCACTTGG-3′ | ||
| SlActin | Forward:5′-AGGCACACAGGTGTTATGGT-3′ | |
| Reverse: 5′-AGCAACTCGAAGCTCATTGT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thangaraj, P.; Balamurali, A.S.; Subbiah, K.A.; Sevugapperumal, N.; Gurudevan, T.; Uthandi, S.; Shanmugam, H. Antifungal Volatilomes Mediated Defense Mechanism against Fusarium oxysporum f. sp. lycopersici, the Incitant of Tomato Wilt. Molecules 2022, 27, 3631. https://doi.org/10.3390/molecules27113631
Thangaraj P, Balamurali AS, Subbiah KA, Sevugapperumal N, Gurudevan T, Uthandi S, Shanmugam H. Antifungal Volatilomes Mediated Defense Mechanism against Fusarium oxysporum f. sp. lycopersici, the Incitant of Tomato Wilt. Molecules. 2022; 27(11):3631. https://doi.org/10.3390/molecules27113631
Chicago/Turabian StyleThangaraj, Praveen, Akshaya Subbaih Balamurali, Krishnamoorthy Akkanna Subbiah, Nakkeeran Sevugapperumal, Thiribhuvanamala Gurudevan, Sivakumar Uthandi, and Haripriya Shanmugam. 2022. "Antifungal Volatilomes Mediated Defense Mechanism against Fusarium oxysporum f. sp. lycopersici, the Incitant of Tomato Wilt" Molecules 27, no. 11: 3631. https://doi.org/10.3390/molecules27113631
APA StyleThangaraj, P., Balamurali, A. S., Subbiah, K. A., Sevugapperumal, N., Gurudevan, T., Uthandi, S., & Shanmugam, H. (2022). Antifungal Volatilomes Mediated Defense Mechanism against Fusarium oxysporum f. sp. lycopersici, the Incitant of Tomato Wilt. Molecules, 27(11), 3631. https://doi.org/10.3390/molecules27113631