
The Intestinal and Biliary Metabolites of Ibuprofen in the Rat with Experimental Hyperglycemia



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Blood Glucose Level
2.2. Fenton Tests
2.3. Udenfriend’s Test Results
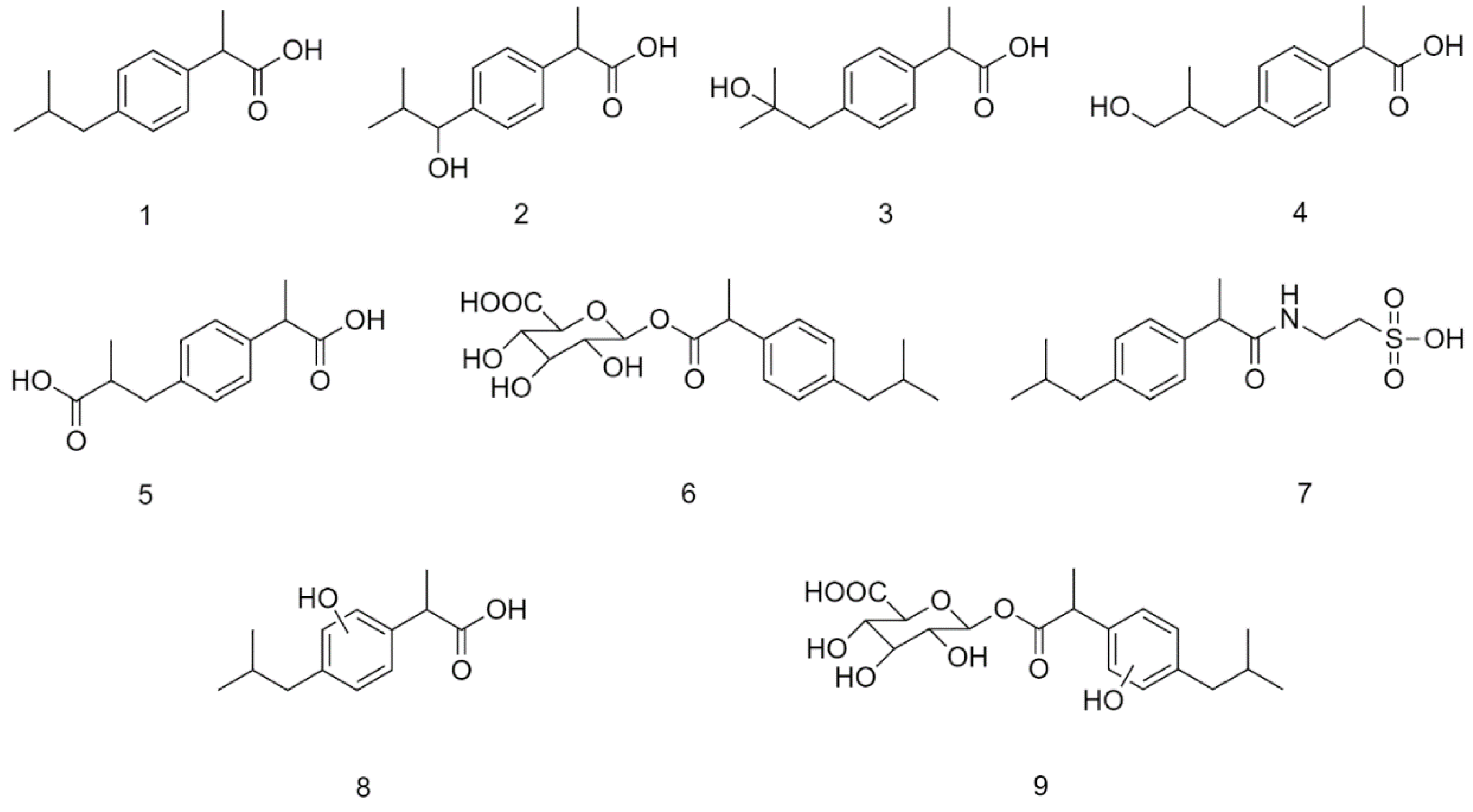
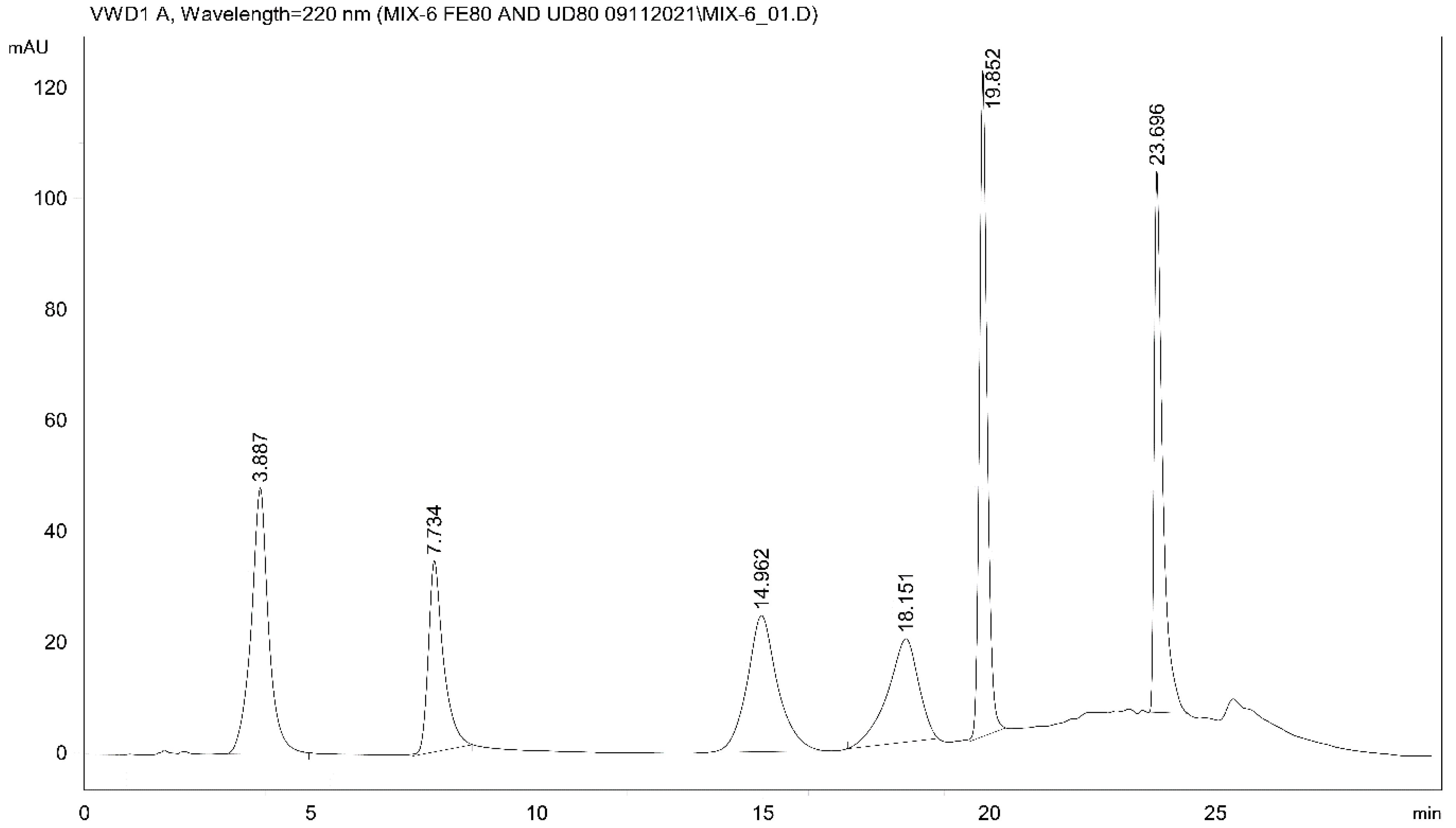
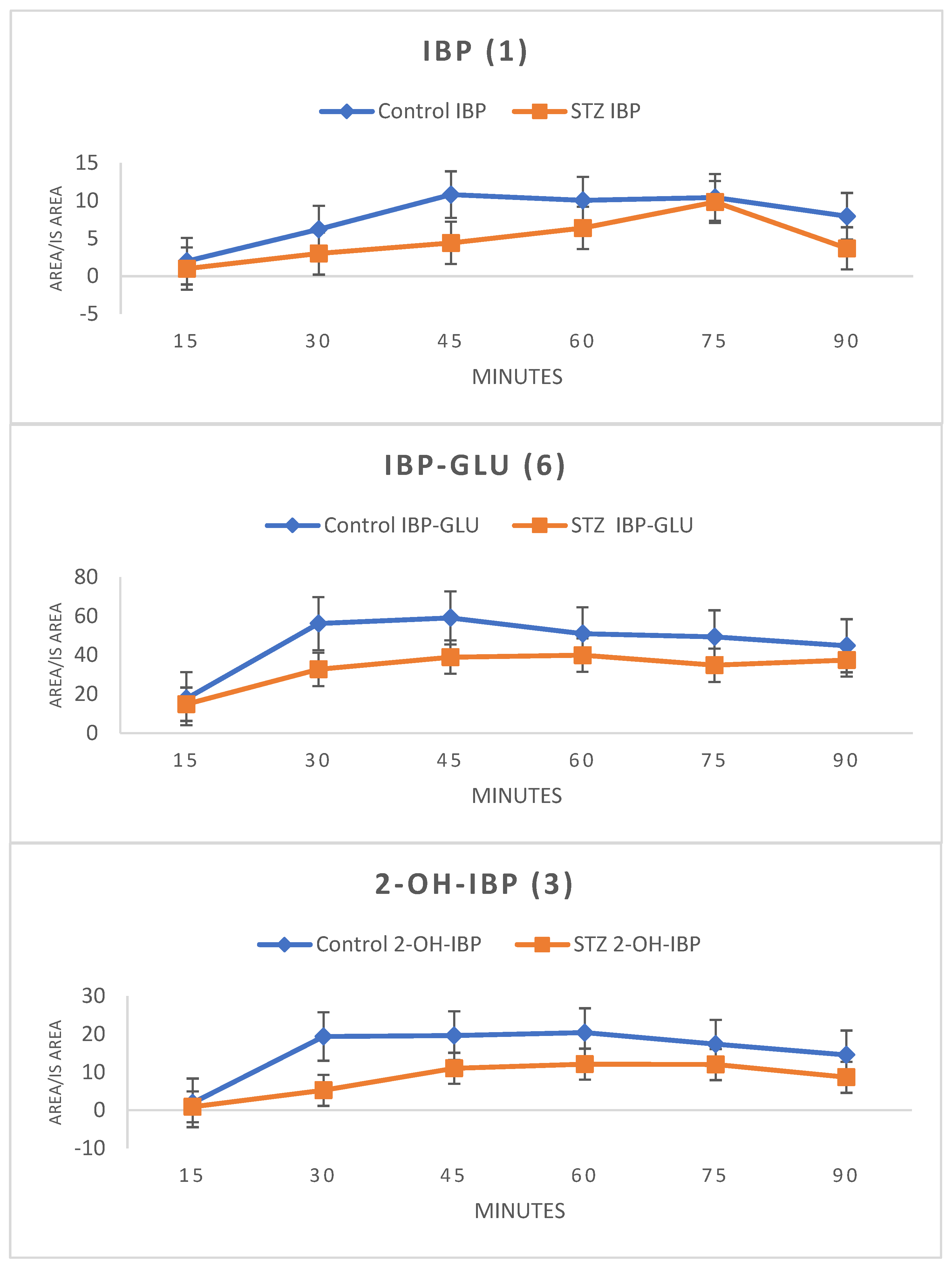
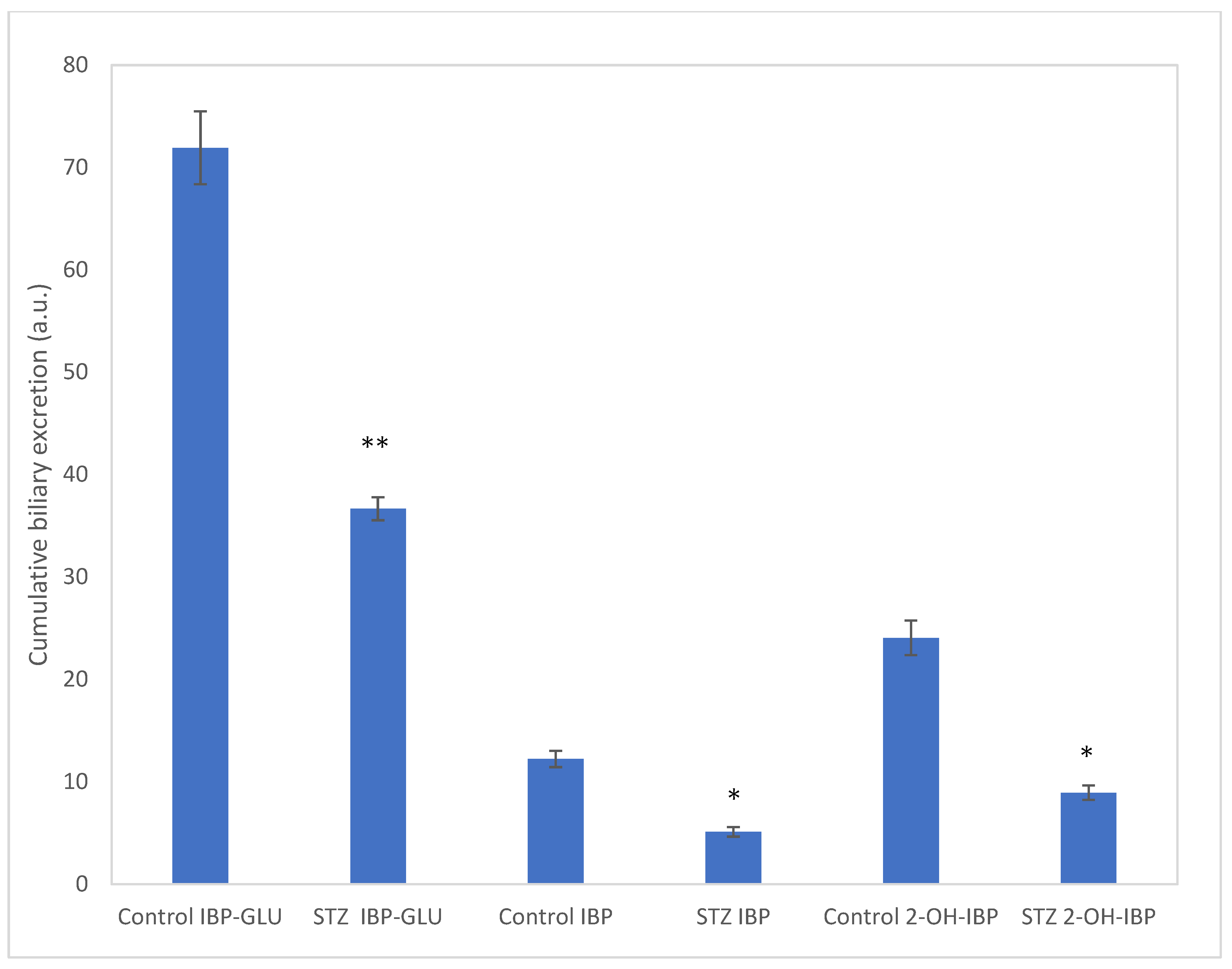
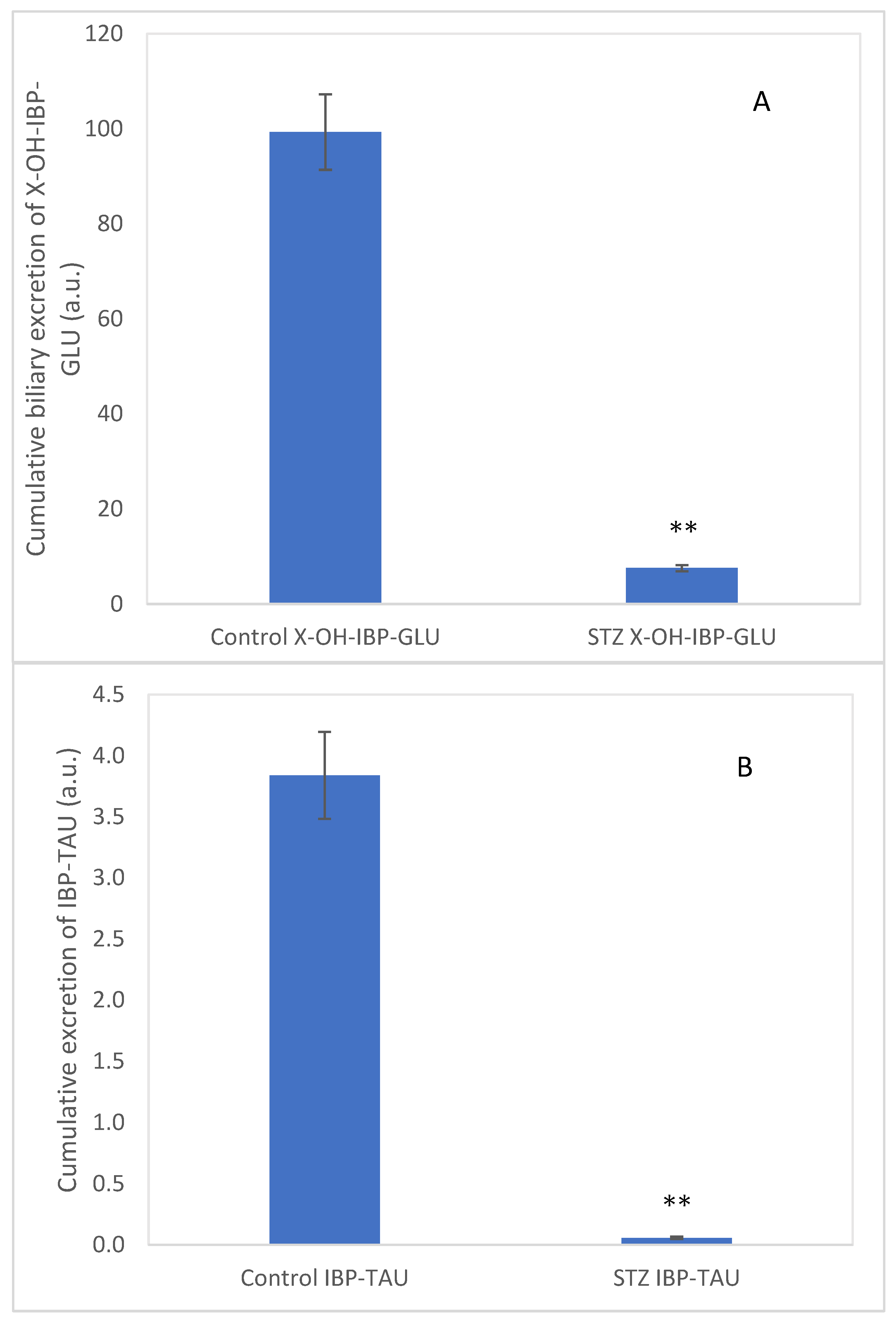
2.4. Analysis of Intestinal Perfusate and Bile Samples
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Fenton Test
4.3. Udenfriend’s Assay
4.4. Animals and Experimental Procedure
4.5. Sample Preparation
4.6. Sample Analysis
4.6.1. HPLC-UV
4.6.2. HPLC-HESI-MS
4.7. Statistical Analysis
4.8. Ethical Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Mazaleuskaya, L.L.; Theken, K.N.; Gong, L.; Thorn, C.F.; FitzGerald, G.A.; Altman, R.B.; Klein, T.E. PharmGKB summary: Ibuprofen pathways. Pharmacogenet. Genom. 2015, 25, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Rainsford, K.D. Ibuprofen: Pharmacology, efficacy and safety. Inflammopharmacology 2009, 17, 275–342. [Google Scholar] [CrossRef] [PubMed]
- Keep, D.R.; Sidelmann, U.G.; Hansen, S.H. Isolation and characterization of major phase I and II metabolites of ibuprofen. Pharm. Res. 1997, 14, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Essman, W.B.; Wollman, S.B. Free radicals, central nervous system processes and brain functions. In Oxygen Radicals: Systemic Events and Disease Processes; Das, D.K., Essman, W.B., Eds.; Karger: Basel, Swizterland, 1990; pp. 172–192. [Google Scholar] [CrossRef]
- Ghiselli, A.; Laurenti, O.; De Mattia, G.; Maiani, G.; Ferro-Luzzi, A. Salicylate hydroxylation as an early marker of in vivo oxidative stress in diabetic patients. Free Radic. Biol. Med. 1992, 13, 621–626. [Google Scholar] [CrossRef]
- Sies, H. Antioxidants in Disease Mechanisms and Therapy. Advances in Pharmacology, 1st ed.; Academic Press: San Diego, CA, USA, 1996; Volume 38. [Google Scholar]
- Chatterjee, S. Chapter Two—Oxidative Stress, Inflammation, and Disease. In Oxidative Stress and Biomaterials; Dziubla, T., Butterfield, D.A., Eds.; Academic Press: Amsterdam, The Netherlands, 2016; pp. 35–58. [Google Scholar] [CrossRef]
- Asmat, U.; Abad, K.; Ismail., K. Diabetes mellitus and oxidative stress—A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Brandes, R.P.; Weissmann, N.; Schröder, K. NADPH oxidases in cardiovascular disease. Free Radic. Biol. Med. 2010, 49, 687–706. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef]
- Dostalek, M.; Sam, W.J.; Paryani, K.R.; Macwan, J.S.; Gohh, R.Y.; Akhlaghi, F. Diabetes mellitus reduces the clearance of atorvastatin lactone. Clin. Pharmacokinet. 2012, 51, 591–606. [Google Scholar] [CrossRef]
- Li, Y.; Meng, Q.; Yang, M.; Liu, D.; Hou, X.; Tang, L.; Wang, X.; Lyu, Y.; Chen, X.; Liu, K.; et al. Current trends in drug metabolism and pharmacokinetics. Acta Pharm. Sin. B. 2019, 9, 1113–1144. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, X. Imbalance of drug transporter-CYPs interplay by diabetes and its clinical significance. Pharmaceutics 2020, 12, 348. [Google Scholar] [CrossRef]
- Almási, A.; Pinto, É.I.L.N.; Kovács, N.P.; Fischer, T.; Markovics, Z.; Fischer, E.; Perjési, P. Changes in hepatic metabolic enzyme activities and biliary excretion of 4-nitrophenol in streptozotocin-induced diabetic rats. Braz. J. Pharm. Sci. 2018, 54, e17347. [Google Scholar] [CrossRef]
- Kovács, N.P.; Almási, A.; Garai, K.; Kuzma, M.; Vancea, S.; Fischer, E.; Perjési, P. Investigation of intestinal elimination and biliary excretion of ibuprofen in hyperglycemic rats. Can. J. Physiol. Pharmacol. 2019, 97, 1080–1089. [Google Scholar] [CrossRef]
- Fischer, E.; Almási, A.; Bojcsev, S.; Fischer, T.; Kovács, N.P.; Perjési, P. Effect of experimental diabetes and insulin replacement on intestinal metabolism and excretion of 4-nitrophenol in rats. Can. J. Physiol. Pharmacol. 2015, 93, 459–464. [Google Scholar] [CrossRef]
- Illés, E.; Takács, E.; Dombi, A.; Gajda-Schrantz, K.; Rácz, G.; Gonter, K.; Wojnárovits, L. Hydroxyl radical induced degradation of ibuprofen. Sci. Total Environ. 2013, 447, 286–292. [Google Scholar] [CrossRef]
- Wang-Fischer, Y.; Garyantes, T. Improving the reliability and utility of streptozotocin-induced rat diabetic model. J. Diabetes Res. 2018, 2018, 8054073. [Google Scholar] [CrossRef] [PubMed]
- Bojcsev, S.; Rafiei, A.; Fischer, E. Changes in the biliary excretion of exogenous organic anions by streptozotocin-induced diabetes. Acta Physiol. Hung. 1996, 84, 263–264. [Google Scholar]
- Nyúl, E.; Kuzma, M.; Mayer, M.; Lakatos, S.; Almási, A.; Perjési, P. HPLC study on Fenton-reaction initiated oxidation of salicylic acid. Biological relevance of the reaction in intestinal biotransformation of salicylic acid. Free Radic. Res. 2018, 52, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Udenfriend, S.; Clark, C.T.; Axelrod, J.; Brodie, B.B. Ascorbic acid in aromatic hydroxylation. I. A model system for aromatic hydroxylation. J. Biol. Chem. 1954, 208, 731–740. [Google Scholar] [CrossRef]
- Brodie, B.B.; Axelrod, J.; Shore, P.A.; Udenfriend, S. Ascorbic acid in aromatic hydroxylation. II. Products formed by reaction of substrates with ascorbic acid, ferrous ion, and oxygen. J. Biol. Chem. 1954, 208, 741–750. [Google Scholar] [CrossRef]
- Leemann, T.D.; Transon, C.; Bonnabry, P.; Dayer, P. A major role for cytochrome P450TB (CYP2C subfamily) in the actions of non-steroidal anti-inflammatory drugs. Drugs Exp. Clin. Res. 1993, 19, 189–195. [Google Scholar] [PubMed]
- Chang, S.Y.; Li, W.; Traeger, S.C.; Wang, B.; Cui, D.; Zhang, H.; Wen, B.; Rodrigues, A.D. Confirmation that cytochrome P450 2C8 (CYP2C8) plays a minor role in (S)-(+)- and (R)-(−)-ibuprofen hydroxylation in vitro. Drug Metab. Dispos. 2008, 36, 2513–2522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamman, M.A.; Thompson, G.A.; Hall, S.D. Regioselective and stereoselective metabolism of ibuprofen by human cytochrome P450 2C. Biochem. Pharmacol. 1997, 54, 33–41. [Google Scholar] [CrossRef]
- Neunzig, I.; Göhring, A.; Dragan, C.A.; Zapp, J.; Peters, F.T.; Maurer, H.H.; Bureik, M. Production and NMR analysis of the human ibuprofen metabolite 3-hydroxyibuprofen. J. Biotechnol. 2012, 157, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Gliszczyńska, A.; Sánchez-López, E. Dexibuprofen therapeutic advances: Prodrugs and nanotechnological formulations. Pharmaceutics 2021, 13, 414. [Google Scholar] [CrossRef] [PubMed]
- Hammer, H.; Schmidt, F.; Marx-Stoelting, P.; Pötz, O.; Braeuning, A. Cross-species analysis of hepatic cytochrome P450 and transport protein expression. Arch. Toxicol. 2021, 95, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.F.; Adams, S.S.; Cliffe, E.E.; Dickinson, W.; Nicholson, J.S. The metabolism of ibuprofen. Xenobiotica 1973, 3, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Yamazoe, Y.; Murayama, N.; Shimada, M.; Yamauchi, K.; Kato, R. Cytochrome P450 in livers of diabetic rats: Regulation by growth hormone and insulin. Arch. Biochem. Biophys. 1989, 268, 567–575. [Google Scholar] [CrossRef]
- Shimojo, N. Cytochrome P450 changes in rats with streptozocin-induced diabetes. Int. J. Biochem. 1994, 26, 1261–1268. [Google Scholar] [CrossRef]
- Richmond, R.; Halliwell, B.; Chauhan, J.; Darbre, A. Superoxide-dependent formation of hydroxyl radicals: Detection of hydroxyl radicals by the hydroxylation of aromatic compounds. Anal. Biochem. 1981, 118, 328–335. [Google Scholar] [CrossRef]
- Forman, H.J. Redox signaling: An evolution from free radicals to aging. Free Radic. Biol. Med. 2016, 97, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82–83, 969–974. [Google Scholar] [CrossRef]
- Biswas, S.; Das, R.; Banerjee, E.R. Role of free radicals in human inflammatory diseases. AIMS Biophys. 2017, 4, 596–614. [Google Scholar] [CrossRef]
- Neyens, E.; Baeyens, J. A review of classic Fenton’s peroxidation as an advanced oxidation technique. J. Hazard. Mater. 2003, 98, 33–50. [Google Scholar] [CrossRef]
- Prousek, J. Fenton chemistry in biology and medicine. Pure Appl. Chem. 2007, 79, 2325–2338. [Google Scholar] [CrossRef]
- Kuzma, M.; Kovács, N.; Sziva, L.; Maász, G.; Avar, P.; Perjési, P. Oxidation of hydroxy- and dihydroxybenzoic acids under the Udenfriend’s conditions. An HPLC study. Open Med. Chem. J. 2018, 12, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, R.B.; Bonin, J.L.; Ardill, L.P.; Rourk, E.J.; Patterson, H.H.; Stemmler, E.A. Photocatalytic degradation of ibuprofen over BiOCl nanosheets with identification of intermediates. J. Hazard. Mat. 2018, 358, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, L.; Varshney, M.; Seubert, C.N.; Stevens, S.M.; Johnson, J.V.; Richard, A.; Yost, R.A.; Dennis, D.M. Mass spectral fragmentation of the intravenous anesthetic propofol and structurally related phenols. J. Am. Soc. Mass Spectrom. 2005, 16, 814–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, L.E.; Fimmen, R.L.; Chin, Y.P.; Mash, H.E.; Weavers, L.K. Fulvic acid mediated photolysis of ibuprofen in water. Water Res. 2011, 45, 4449–4458. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Noerpel, M.; Luk, H.L.; Wei, Z.; Spinney, R. Thermodynamic and kinetic study of ibuprofen with hydroxyl radical: A density functional theory approach. Int. J. Quant. Chem. 2014, 114, 74–83. [Google Scholar] [CrossRef]
- Chopra, S.; Kumar, D. Ibuprofen as an emerging organic contaminant in environment, distribution and remediation. Heliyon 2020, 6, e04087. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Ueno, K.; Mitarai, A.; Sasaki, K. Evidence for hydroxyl radicals as an active species generated from Udenfriend’s reagent. J. Chem. Soc. Perkin Trans. 1993, 255–259. [Google Scholar] [CrossRef]
- Enamia, S.; Sakamoto, Y.; Colussie, A.J. Fenton chemistry at aqueous interfaces. Proc. Natl. Acad. Sci. USA 2014, 111, 623–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Carlson, S.; Kinzer, J.A.; Perpall, H.J. HPLC and LC-MS studies of hydroxylation of phenylalanine as an assay for hydroxyl radicals generated from Udenfriend’s reagent. Biochem. Biophys. Res. Commun. 2003, 312, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Shirley, M.A.; Guan, X.; Kaiser, D.G.; Halstead, G.W.; Baillie, T.A. Taurine conjugation of ibuprofen in humans and in rat liver in vitro. Relationship to metabolic chiral inversion. J. Pharmacol. Exp. Ther. 1994, 269, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Kishimoto, S.; Shibatani, N.; Nomura, H.; Ishii, Y.; Onishi, M.; Inotsume, N.; Takeuchi, Y.; Fukushima, S. The pharmacokinetics of morphine and its glucuronide conjugate in a rat model of streptozotocin-induced diabetes and the expression of MRP2, MRP3 and UGT2B1 in the liver. J. Pharm. Pharmacol. 2010, 62, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Anger, G.J.; Magomedova, L.; Piquette-Miller, M. Impact of acute streptozotocin-induced diabetes on ABC transporter expression in rats. Chem. Biodiv. 2009, 6, 1943–1959. [Google Scholar] [CrossRef]
- Nawa, A.; Hamabe, W.F.; Tokuyama, S. Inducible nitric oxide synthase-mediated decrease of intestinal P-glycoprotein expression under streptozotocin-induced diabetic conditions. Life Sci. 2010, 86, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Lu, L.; Jin, S.; Jing, X.Y.; Yao, D.; Hu, N..; Liu, L.; Duan, R.; Liu, X.D.; Wang, G.J.; et al. Tissue-specific alterations in expression and function of P-glycoprotein in streptozotocin-induced diabetic rats. Acta Pharmacol. Sin. 2011, 32, 956–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jetter, A.; Kullak-Ublick, G.A. Drugs and hepatic transporters: A review. Pharmacol. Res. 2020, 154, 104234. [Google Scholar] [CrossRef]
- Schaffer, S.W.; Azuma, J.; Mozaffari, M. Role of antioxidant activity of taurine in diabetes. Can. J. Physiol. Pharmacol. 2009, 87, 91–99. [Google Scholar] [CrossRef]
- Baliou, S.; Adamaki, M.; Ioannou, P.; Pappa, A.; Panayiotidis, M.I.; Spandidos, D.A.; Christodoulou, I.; Kyriakopoulos, A.M.; Zoumpourlis, V. Protective role of taurine against oxidative stress (Review). Mol. Med. Rep. 2021, 24, 605. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D. Bile flow and composition during bile acid depletion and administration. Can. J. Physiol. Pharmacol. 1974, 52, 334–348. [Google Scholar] [CrossRef] [PubMed]
- European Legislation (Directive 2010/63/E.U.). ELI. Available online: https://eur-lex.europa.eu/eli/dir/2010/63/oj (accessed on 4 June 2022).
- Hungarian Government Regulation (40/2013., II. 14). Available online: https://www.fao.org/faolex/results/details/en/c/LEX-FAOC124420 (accessed on 4 June 2022).
- Kuzma, M.; Fodor, K.; Maász, G.; Avar, P.; Mózsik, G.; Past, T.; Fischer, E.; Perjési, P. A validated HPLC-FLD method for analysis of intestinal absorption and metabolism of capsaicin and dihydrocapsaicin in the rat. J. Pharm. Biomed. Anal. 2015, 103, 59–66. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, H.O.; Almási, A.; Molnár, S.; Perjési, P. The Intestinal and Biliary Metabolites of Ibuprofen in the Rat with Experimental Hyperglycemia. Molecules 2022, 27, 4000. https://doi.org/10.3390/molecules27134000
Mohammed HO, Almási A, Molnár S, Perjési P. The Intestinal and Biliary Metabolites of Ibuprofen in the Rat with Experimental Hyperglycemia. Molecules. 2022; 27(13):4000. https://doi.org/10.3390/molecules27134000
Chicago/Turabian StyleMohammed, Hawsar Othman, Attila Almási, Szilárd Molnár, and Pál Perjési. 2022. "The Intestinal and Biliary Metabolites of Ibuprofen in the Rat with Experimental Hyperglycemia" Molecules 27, no. 13: 4000. https://doi.org/10.3390/molecules27134000
APA StyleMohammed, H. O., Almási, A., Molnár, S., & Perjési, P. (2022). The Intestinal and Biliary Metabolites of Ibuprofen in the Rat with Experimental Hyperglycemia. Molecules, 27(13), 4000. https://doi.org/10.3390/molecules27134000