
Complexation of Anthocyanin-Bound Blackcurrant Pectin and Whey Protein: Effect of pH and Heat Treatment


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Complexation of Blackcurrant Pectin and Whey Protein
2.3. Stability Measurement
2.4. Particle Size and Zeta-Potential Measurements
2.5. Quantification of Galacturonic Acid
2.6. Fourier Transform Infrared Spectroscopy
2.7. Statistical Analysis
3. Results and Discussion
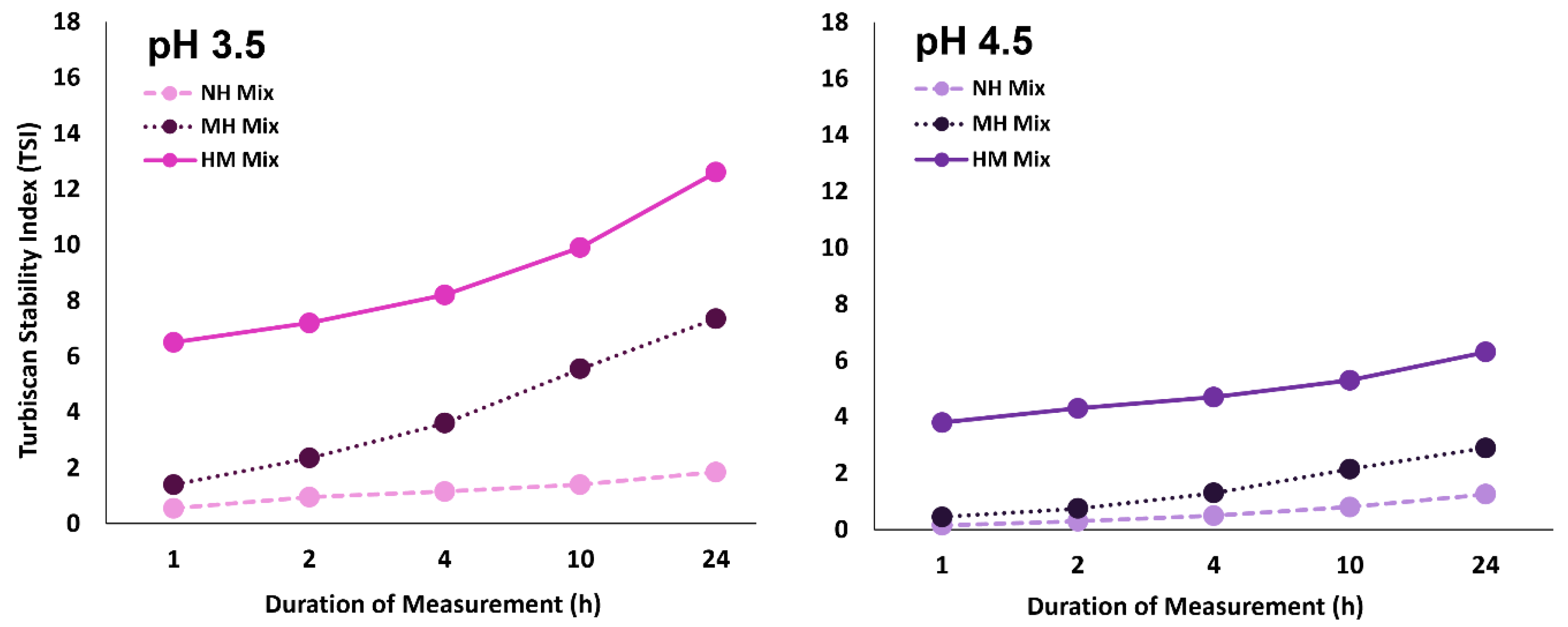
3.1. Visual Appearance and Stability of Blackcurrant Pectin-Whey Protein Mixtures
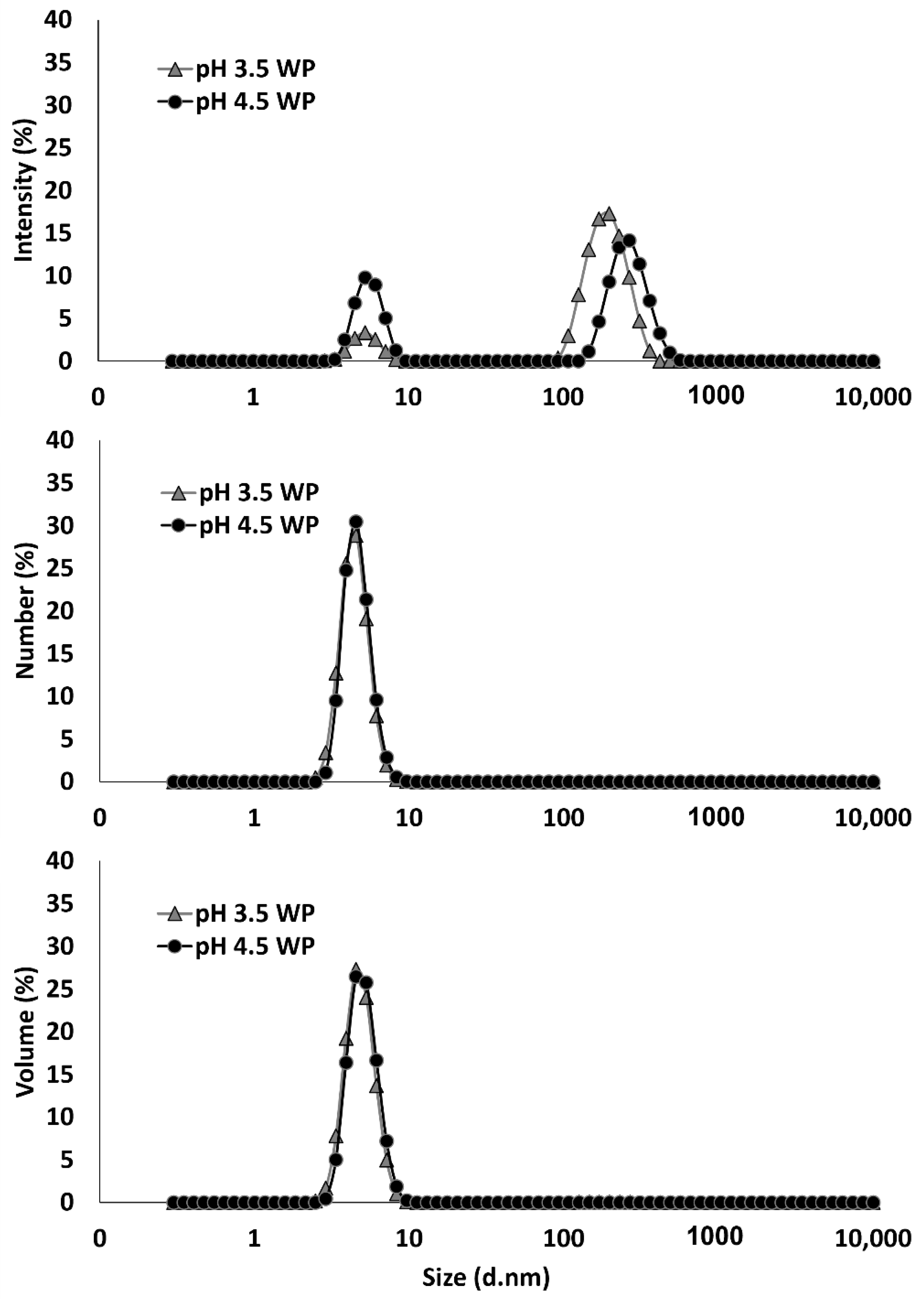
3.2. Particle Size and Zeta-Potential of Blackcurrant Pectin-Whey Protein Complexes
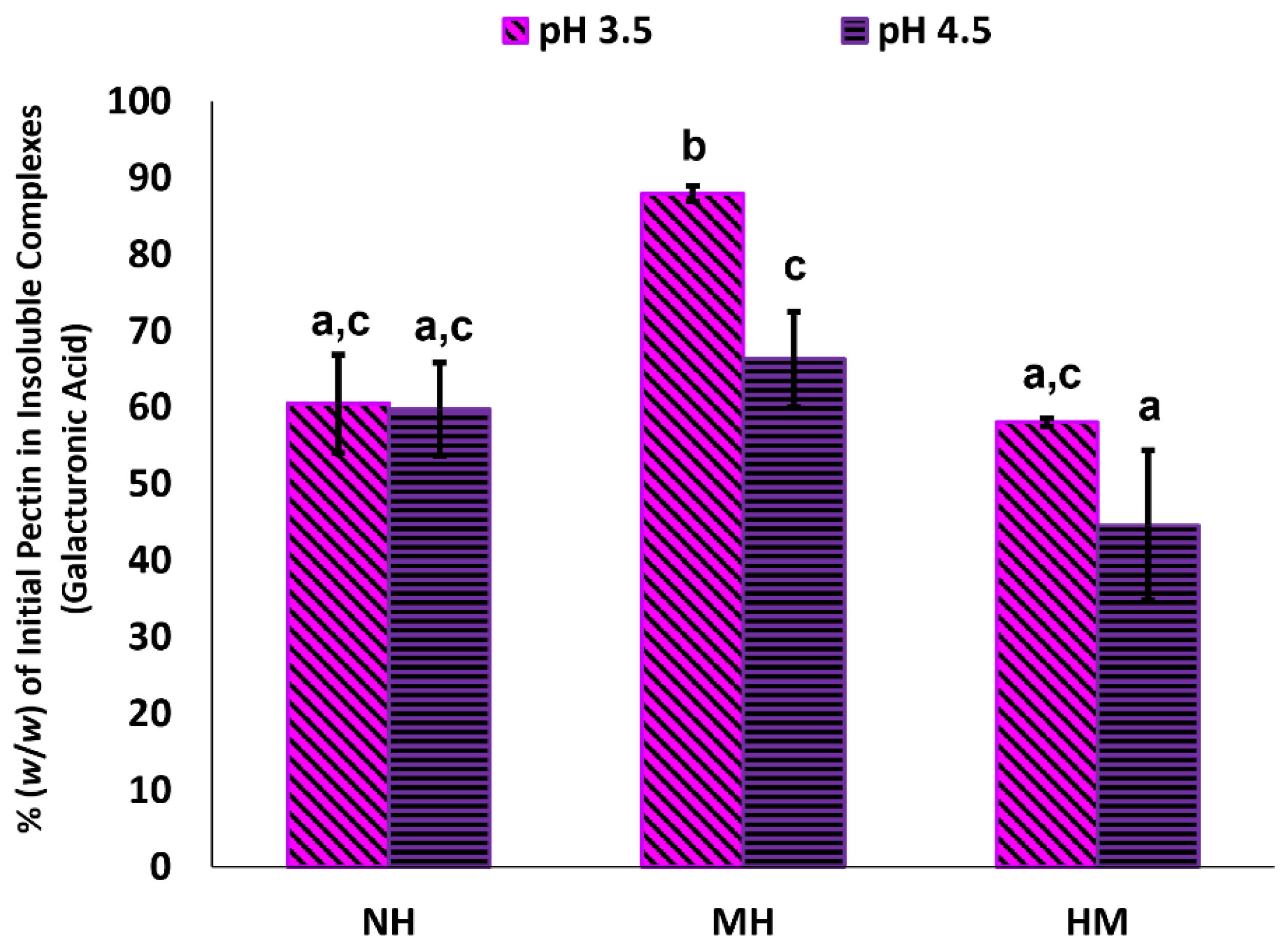
3.3. Insoluble Complexes in Blackcurrant Pectin-Whey Protein Mixtures
3.4. Overall Discussion: Possible Mechanism of Complexation between Blackcurrant Pectin and Whey Protein
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Thongkaew, C.; Gibis, M.; Hinrichs, J.; Weiss, J. Polyphenol interactions with whey protein isolate and whey protein isolate–pectin coacervates. Food Hydrocoll. 2014, 41, 103–112. [Google Scholar] [CrossRef]
- Chevalier, L.M.; Rioux, L.-E.; Angers, P.; Turgeon, S.L. Study of the interactions between pectin in a blueberry puree and whey proteins: Functionality and application. Food Hydrocoll. 2019, 87, 61–70. [Google Scholar] [CrossRef]
- Chung, C.; Rojanasasithara, T.; Mutilangi, W.; McClements, D.J. Enhanced stability of anthocyanin-based color in model beverage systems through whey protein isolate complexation. Food Res. Int. 2015, 76, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Millet, M.; Poupard, P.; Le Quéré, J.-M.; Bauduin, R.; Guyot, S. Haze in apple-based beverages: Detailed polyphenol, polysaccharide, protein, and mineral compositions. J. Agric. Food Chem. 2017, 65, 6404–6414. [Google Scholar] [CrossRef]
- Lin, J.; Tan, Y.X.G.; Leong, L.P.; Zhou, W. Steamed bread enriched with quercetin as an antiglycative food product: Its quality attributes and antioxidant properties. Food Funct. 2018, 9, 3398–3407. [Google Scholar] [CrossRef]
- Jauregi, P.; Olatujoye, J.B.; Cabezudo, I.; Frazier, R.A.; Gordon, M.H. Astringency reduction in red wine by whey proteins. Food Chem. 2016, 199, 547–555. [Google Scholar] [CrossRef]
- Rinaldi, A.; Gambuti, A.; Moine-Ledoux, V.; Moio, L. Evaluation of the astringency of commercial tannins by means of the SDS–PAGE-based method. Food Chem. 2010, 122, 951–956. [Google Scholar] [CrossRef]
- Stănciuc, N.; Turturică, M.; Oancea, A.M.; Barbu, V.; Ioniţă, E.; Aprodu, I.; Râpeanu, G. Microencapsulation of anthocyanins from grape skins by whey protein isolates and different polymers. Food Bioprocess Technol. 2017, 10, 1715–1726. [Google Scholar] [CrossRef]
- Wei, Z.; Huang, Q. Assembly of protein–polysaccharide complexes for delivery of bioactive ingredients: A perspective paper. J. Agric. Food Chem. 2019, 67, 1344–1352. [Google Scholar] [CrossRef]
- Yildirim-Elikoglu, S.; Erdem, Y.K. Interactions between milk proteins and polyphenols: Binding mechanisms, related changes, and the future trends in the dairy industry. Food Rev. Int. 2018, 34, 665–697. [Google Scholar] [CrossRef]
- Schmitt, C.; Sanchez, C.; Desobry-Banon, S.; Hardy, J. Structure and technofunctional properties of protein-polysaccharide complexes: A review. Crit. Rev. Food Sci. Nutr. 1998, 38, 689–753. [Google Scholar] [CrossRef] [PubMed]
- Siebert, K.J.; Troukhanova, N.V.; Lynn, P.Y. Nature of polyphenol-protein interactions. J. Agric. Food Chem. 1996, 44, 80–85. [Google Scholar] [CrossRef]
- McClements, D.J. Non-covalent interactions between proteins and polysaccharides. Biotechnol. Adv. 2006, 24, 621–625. [Google Scholar] [CrossRef]
- Spencer, C.M.; Cai, Y.; Martin, R.; Gaffney, S.H.; Goulding, P.N.; Magnolato, D.; Lilley, T.H.; Haslam, E. Polyphenol Complexation—Some thoughts and observations. Phytochemistry 1988, 27, 2397–2409. [Google Scholar] [CrossRef]
- Dangles, O.; Dufour, C. Flavonoid-protein interactions. In Flavonoids: Chemistry, Biochemistry and Applications, 1st ed.; Andersen, Ø.M., Markham, K.R., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 443–469. [Google Scholar]
- Dangles, O.; Fenger, J.-A. The chemical reactivity of anthocyanins and its consequences in food science and nutrition. Molecules 2018, 23, 1970. [Google Scholar] [CrossRef] [Green Version]
- Dumay, E.; Laligant, A.; Zasypkin, D.; Cheftel, J.C. Pressure- and heat-induced gelation of mixed β-lactoglobulin/polysaccharide solutions: Scanning electron microscopy of gels. Food Hydrocoll. 1999, 13, 339–351. [Google Scholar] [CrossRef]
- Wang, Q.; Qvist, K.B. Investigation of the composite system of β-lactoglobulin and pectin in aqueous solutions. Food Res. Int. 2000, 33, 683–690. [Google Scholar] [CrossRef]
- Girard, M.; Sanchez, C.; Laneuville, S.I.; Turgeon, S.L.; Gauthier, S.F. Associative phase separation of β-lactoglobulin/pectin solutions: A kinetic study by small angle static light scattering. Colloids Surf. B 2004, 35, 15–22. [Google Scholar] [CrossRef]
- Girard, M.; Turgeon, S.L.; Gauthier, S.F. Quantification of the interactions between β-lactoglobulin and pectin through capillary electrophoresis analysis. J. Agric. Food Chem. 2003, 51, 6043–6049. [Google Scholar] [CrossRef]
- Girard, M.; Turgeon, S.L.; Gauthier, S.F. Thermodynamic parameters of β-lactoglobulin−pectin complexes assessed by isothermal titration calorimetry. J. Agric. Food Chem. 2003, 51, 4450–4455. [Google Scholar] [CrossRef]
- Girard, M.; Turgeon, S.L.; Gauthier, S.F. Interbiopolymer complexing between β-lactoglobulin and low- and high-methylated pectin measured by potentiometric titration and ultrafiltration. Food Hydrocoll. 2002, 16, 585–591. [Google Scholar] [CrossRef]
- Sperber, B.L.H.M.; Schols, H.A.; Cohen Stuart, M.A.; Norde, W.; Voragen, A.G.J. Influence of the overall charge and local charge density of pectin on the complex formation between pectin and β-lactoglobulin. Food Hydrocoll. 2009, 23, 765–772. [Google Scholar] [CrossRef]
- Salleh, N.; Goh, K.K.T.; Sims, I.M.; Bell, T.J.; Huffman, L.M.; Weeks, M.; Matia-Merino, L. Characterization of anthocyanin-bound pectin-rich fraction extracted from New Zealand blackcurrant (Ribes nigrum) juice. ACS Food Sci. Technol. 2021, 1, 1130–1142. [Google Scholar] [CrossRef]
- Oancea, S. A review of the current knowledge of thermal stability of anthocyanins and approaches to their stabilization to heat. Antioxidants 2021, 10, 1337. [Google Scholar] [CrossRef]
- Rivas-Gonzalo, J.C. Analysis of anthocyanins. In Methods in Polyphenol Analysis, 1st ed.; Santos-Buelga, C., Williamson, G., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2003; pp. 338–358. [Google Scholar]
- Brouillard, R.; Delaporte, B. Chemistry of anthocyanin pigments. 2. Kinetic and thermodynamic study of proton-transfer, hydration, and tautomeric reactions of malvidin 3-glucoside. J. Am. Chem. Soc. 1977, 99, 8461–8468. [Google Scholar] [CrossRef]
- Vuong, T.T.; Hongsprabhas, P. Influences of pH on binding mechanisms of anthocyanins from butterfly pea flower (Clitoria ternatea) with whey powder and whey protein isolate. Cogent Food Agric. 2021, 7, 1889098. [Google Scholar] [CrossRef]
- Fernandes, A.; Brás, N.F.; Mateus, N.; de Freitas, V. Understanding the molecular mechanism of anthocyanin binding to pectin. Langmuir 2014, 30, 8516–8527. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Fischer, J.; Wicker, L. Intermolecular binding of blueberry pectin-rich fractions and anthocyanin. Food Chem. 2016, 194, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Dannenberg, F.; Kessler, H.-G. Reaction kinetics of the denaturation of whey proteins in milk. J. Food Sci. 1988, 53, 258–263. [Google Scholar] [CrossRef]
- Anema, S.G. The thermal denaturation of the total whey protein in reconstituted whole milk. Int. J. Dairy Technol. 2017, 70, 332–338. [Google Scholar] [CrossRef]
- Law, A.J.R.; Leaver, J. Effect of pH on the thermal denaturation of whey proteins in milk. J. Agric. Food Chem. 2000, 48, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; von Staszewski, M.; Pizones Ruiz-Henestrosa, V.M.; Pintado, M.; Pilosof, A.M.R. Impact of pectin or chitosan on bulk, interfacial and antioxidant properties of (+)-catechin and β-lactoglobulin ternary mixtures. Food Hydrocoll. 2016, 55, 119–127. [Google Scholar] [CrossRef]
- Stănciuc, N.; Oancea, A.M.; Aprodu, I.; Turturică, M.; Barbu, V.; Ioniţă, E.; Râpeanu, G.; Bahrim, G. Investigations on binding mechanism of bioactives from elderberry (Sambucus nigra L.) by whey proteins for efficient microencapsulation. J. Food Eng. 2018, 223, 197–207. [Google Scholar] [CrossRef]
- Cheng, H.; Fang, Z.; Liu, T.; Gao, Y.; Liang, L. A study on β-lactoglobulin-triligand-pectin complex particle: Formation, characterization and protection. Food Hydrocoll. 2018, 84, 93–103. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Yang, Y.; Wang, C.; Zhang, T. Formation and characterization of noncovalent ternary complexes based on whey protein concentrate, high methoxyl pectin, and phenolic acid. J. Dairy Sci. 2022, 105, 2963–2977. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xiao, L.; Wen, Y.; Qin, W.; Wu, D.; Chen, H.; Zhang, Q.; Zhang, Q. Comparison of apple polyphenol-gelatin binary complex and apple polyphenol-gelatin-pectin ternary complex: Antioxidant and structural characterization. LWT 2021, 148, 111740. [Google Scholar] [CrossRef]
- Schneider, M.; Esposito, D.; Lila, M.A.; Foegeding, E.A. Formation of whey protein-polyphenol mesostructures as a natural means of creating functional particles. Food Funct. 2016, 7, 1306–1318. [Google Scholar] [CrossRef]
- Rubio-Senent, F.; Rodríguez-Gutiérrez, G.; Lama-Muñoz, A.; García, A.; Fernández-Bolaños, J. Novel pectin present in new olive mill wastewater with similar emulsifying and better biological properties than citrus pectin. Food Hydrocoll. 2015, 50, 237–246. [Google Scholar] [CrossRef]
- Izydorczyk, M. Understanding the chemistry of food carbohydrates. In Food Carbohydrates: Chemistry, Physical Properties, and Applications, 1st ed.; Cui, S.W., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 1–65. [Google Scholar]
- Wu, J.; Du, B.; Li, J.; Zhang, H. Influence of homogenisation and the degradation of stabilizer on the stability of acidified milk drinks stabilized by carboxymethylcellulose. LWT—Food Sci. Technol. 2014, 56, 370–376. [Google Scholar] [CrossRef]
- Schmitt, C.; Sanchez, C.; Despond, S.; Renard, D.; Thomas, F.; Hardy, J. Effect of protein aggregates on the complex coacervation between β-lactoglobulin and acacia gum at pH 4.2. Food Hydrocoll. 2000, 14, 403–413. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- O’Donoghue, E.M.; Somerfield, S.D.; Deroles, S.C.; Sutherland, P.W.; Hallett, I.C.; Erridge, Z.A.; Brummell, D.A.; Hunter, D.A. Simultaneous knock-down of six beta-galactosidase genes in petunia petals prevents loss of pectic galactan but decreases petal strength. Plant Physiol. Biochem. 2017, 113, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.J.; von Elbe, J.H.; Giusti, M.M. Colorants. In Fennema’s Food Chemistry, 4th ed.; Damodaran, S., Parkin, K.L., Fennema, O.R., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 571–638. [Google Scholar]
- Patras, A.; Brunton, N.P.; O’Donnell, C.; Tiwari, B.K. Effect of thermal processing on anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends Food Sci. Technol. 2010, 21, 3–11. [Google Scholar] [CrossRef]
- Schmidt, U.S.; Schütz, L.; Schuchmann, H.P. Interfacial and emulsifying properties of citrus pectin: Interaction of pH, ionic strength and degree of esterification. Food Hydrocoll. 2017, 62, 288–298. [Google Scholar] [CrossRef]
- Donovan, M.; Mulvihill, D.M. Thermal denaturation and aggregation of whey proteins. Ir. J. Food Sci. Technol. 1987, 11, 87–100. [Google Scholar]
- Spiegel, T.; Huss, M. Whey protein aggregation under shear conditions—effects of pH-value and removal of calcium. Int. J. Food Sci. Technol. 2002, 37, 559–568. [Google Scholar] [CrossRef]
- Masuelli, M.A. Viscometric study of pectin. Effect of temperature on the hydrodynamic properties. Int. J. Biol. Macromol. 2011, 48, 286–291. [Google Scholar] [CrossRef]
- Anema, S.G. The whey proteins in milk: Thermal denaturation, physical interactions and effects on the functional properties of milk. In Milk Proteins: From Expression to Food, 1st ed.; Boland, M., Singh, H., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 239–281. [Google Scholar]
- Damodaran, S. Amino acids, peptides, and proteins. In Fennema’s Food Chemistry, 4th ed.; Damodaran, S., Parkin, K.L., Fennema, O.R., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 217–329. [Google Scholar]
- Jiang, S.; Altaf Hussain, M.; Cheng, J.; Jiang, Z.; Geng, H.; Sun, Y.; Sun, C.; Hou, J. Effect of heat treatment on physicochemical and emulsifying properties of polymerized whey protein concentrate and polymerized whey protein isolate. LWT 2018, 98, 134–140. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiao, Q. Effect of chitosan on the heat stability of whey protein solution as a function of pH. J. Sci. Food Agric. 2017, 97, 1576–1581. [Google Scholar] [CrossRef]
- Bhattacharjee, S. DLS and zeta potential—What they are and what they are not? J. Controll. Release 2016, 235, 337–351. [Google Scholar] [CrossRef]
- Peña-Ramos, E.A.; Xiong, Y.L.; Arteaga, G.E. Fractionation and characterisation for antioxidant activity of hydrolysed whey protein. J. Sci. Food Agric. 2004, 84, 1908–1918. [Google Scholar] [CrossRef]
- Choong, Y.K.; Mohd Yousof, N.S.A.; Jamal, J.A.; Wasiman, M.I. Determination of anthocyanin content in two varieties of Hibiscus Sabdariffa from Selangor, Malaysia using a combination of chromatography and spectroscopy. J. Plant Sci. Phytopathol. 2019, 3, 067–075. [Google Scholar] [CrossRef] [Green Version]
- Pretsch, E.; Bühlmann, P.; Badertscher, M. IR spectroscopy. In Structure Determination of Organic Compounds: Tables of Spectral Data, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–67. [Google Scholar]
- Zaleska, H.; Ring, S.G.; Tomasik, P. Apple pectin complexes with whey protein isolate. Food Hydrocoll. 2000, 14, 377–382. [Google Scholar] [CrossRef]
- Mierczynska, J.; Cybulska, J.; Zdunek, A. Rheological and chemical properties of pectin enriched fractions from different sources extracted with citric acid. Carbohydr. Polym. 2017, 156, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Wahyuningsih, S.; Wulandari, L.; Wartono, M.W.; Munawaroh, H.; Ramelan, A.H. The effect of pH and color stability of anthocyanin on food colorant. IOP Conf. Ser. Mater. Sci. Eng. 2017, 193, 012047. [Google Scholar] [CrossRef] [Green Version]
- Loypimai, P.; Moongngarm, A.; Chottanom, P. Thermal and pH degradation kinetics of anthocyanins in natural food colorant prepared from black rice bran. J. Food Sci. Technol. 2016, 53, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Santipanichwong, R.; Suphantharika, M.; Weiss, J.; McClements, D.J. Core-shell biopolymer nanoparticles produced by electrostatic deposition of beet pectin onto heat-denatured β-lactoglobulin aggregates. J. Food Sci. 2008, 73, N23–N30. [Google Scholar] [CrossRef]
- Jones, O.G.; McClements, D.J. Recent progress in biopolymer nanoparticle and microparticle formation by heat-treating electrostatic protein–polysaccharide complexes. Adv. Colloid Interface Sci. 2011, 167, 49–62. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polydispersity Index | ||
|---|---|---|
| pH 3.5 | pH 4.5 | |
| NH Mix | 0.21 ± 0.02 a | 0.39 ± 0.05 b |
| MH Mix | 0.12 ± 0.03 c | 0.26 ± 0.01 a |
| HM Mix | 0.52 ± 0.03 d | 0.37 ± 0.03 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salleh, N.; Goh, K.K.T.; Waterland, M.R.; Huffman, L.M.; Weeks, M.; Matia-Merino, L. Complexation of Anthocyanin-Bound Blackcurrant Pectin and Whey Protein: Effect of pH and Heat Treatment. Molecules 2022, 27, 4202. https://doi.org/10.3390/molecules27134202
Salleh N, Goh KKT, Waterland MR, Huffman LM, Weeks M, Matia-Merino L. Complexation of Anthocyanin-Bound Blackcurrant Pectin and Whey Protein: Effect of pH and Heat Treatment. Molecules. 2022; 27(13):4202. https://doi.org/10.3390/molecules27134202
Chicago/Turabian StyleSalleh, Nurhazwani, Kelvin K. T. Goh, Mark R. Waterland, Lee M. Huffman, Mike Weeks, and Lara Matia-Merino. 2022. "Complexation of Anthocyanin-Bound Blackcurrant Pectin and Whey Protein: Effect of pH and Heat Treatment" Molecules 27, no. 13: 4202. https://doi.org/10.3390/molecules27134202
APA StyleSalleh, N., Goh, K. K. T., Waterland, M. R., Huffman, L. M., Weeks, M., & Matia-Merino, L. (2022). Complexation of Anthocyanin-Bound Blackcurrant Pectin and Whey Protein: Effect of pH and Heat Treatment. Molecules, 27(13), 4202. https://doi.org/10.3390/molecules27134202