
Antifeedant Mechanism of Dodonaea viscosa Saponin A Isolated from the Seeds of Dodonaea viscosa


Abstract
:
1. Introduction
2. Results
2.1. Antifeedant Activity of TSDV
2.2. Effects of TSDV on Life-History Traits
2.3. Time-Course of the Change in Feeding Response to TSDV
2.4. Time-Course of Detoxification Enzyme Induction by TSDV
2.5. Effect of Inhibition of Detoxification Enzymes on the Feeding Response to TSDV
2.6. Time-Course of GAD Activity and GABA Content Induction by TSDV
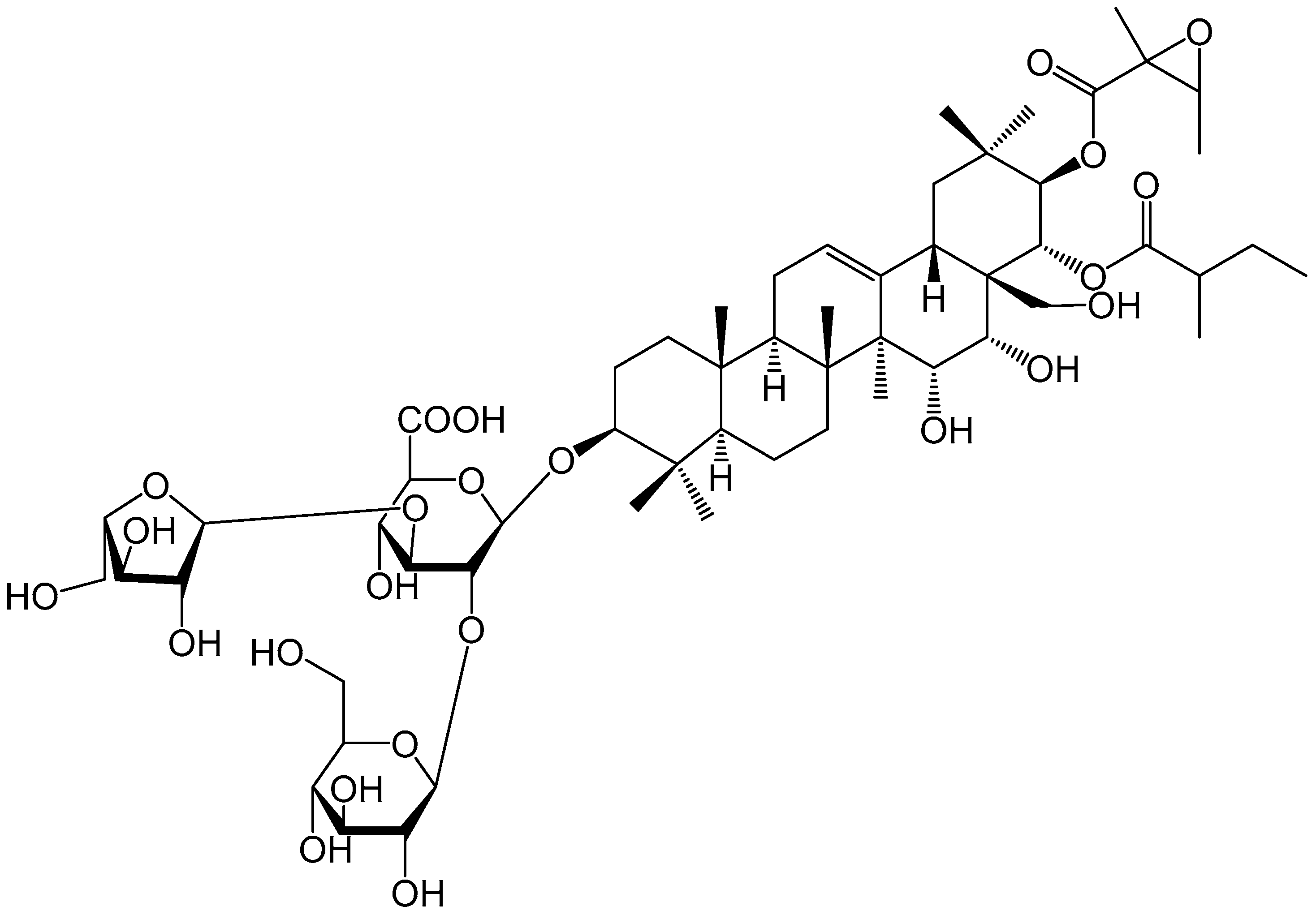
2.7. Isolation of Antifeedant Active Compound
2.8. Evaluation of the Antifeedant Activities of DVSA
2.9. Time-Course of Detoxification Enzyme Induction by DVSA
2.10. Effect of the Inhibition of Detoxification Enzymes on the Feeding Response to DVSA
2.11. Time-Course of GAD Activity and GABA Content Induction by DVSA
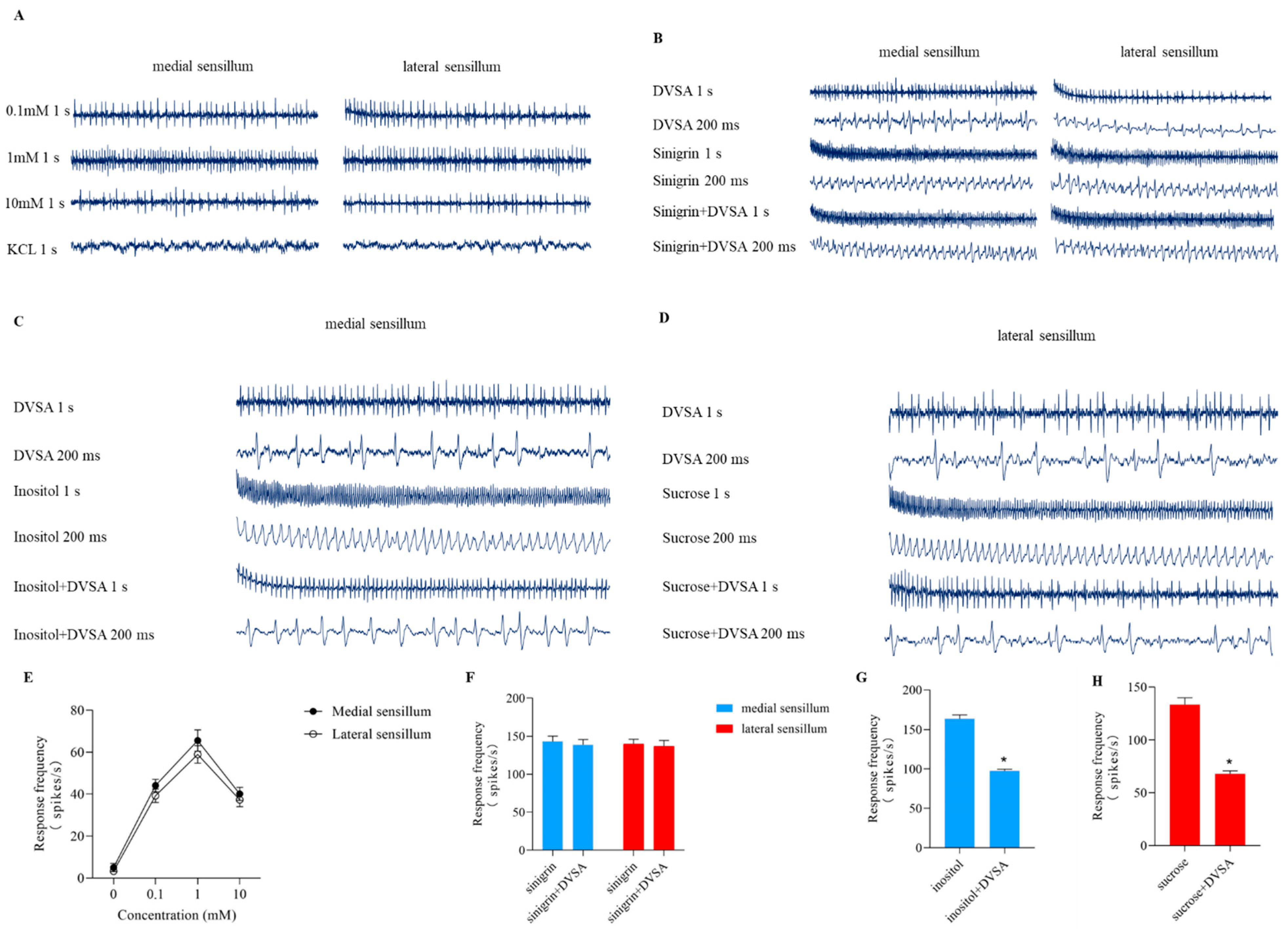
2.12. Effects of DVSA on the Taste Sensillum of S. litura
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Insects
4.4. Assays for Nonselective Antifeedant Activity
4.5. Detoxification Enzymes Assays
4.6. Effect of the Inhibition of Detoxification Enzymes Activities on the Feeding Response in Larvae to D. viscosa
4.7. Levels of GABA Content and GAD Activity Assays
4.8. Effects of DVSA on the Taste Sensillum of S. litura
4.9. Isolation of DVSA
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Muhammad, A.; Tel-Çayan, G.; Öztürk, M.; Duru, M.E.; Nadeem, S.; Anis, I.; Ng, S.W.; Shah, M.R. Phytochemicals from Dodonaea viscosa and their antioxidant and anticholinesterase activities with structure–activity relationships. Pharm. Biol. 2016, 54, 1649–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1979; Volume 58. [Google Scholar]
- Peng, S.L.; Chen, A.Q.; Fang, H.D.; Wu, J.L.; Liu, G.C. Effects of vegetation restoration types on soil quality in Yuanmou dry-hot valley, China. Soil Sci. Plant Nutr. 2013, 59, 347–360. [Google Scholar] [CrossRef] [Green Version]
- Mendoza-Hernández, P.E.; Orozco-Segovia, A.; Meave, J.A.; Valverde, T.; Martínez-Ramos, M. Vegetation recovery and plant facilitation in a human-disturbed lava field in a megacity: Searching tools for ecosystem restoration. Plant Ecol. 2013, 214, 153–167. [Google Scholar] [CrossRef]
- Estevez-Torres, J.; García-Sánchez, R.; Monroy-Ata, A.; Ríos-Gómez, R. Establecimiento de plantas mediante el uso de micorrizas y de islas de recursos en un matorral xerófilo deteriorado. Bot. Sci. 2007, 80, 49–57. [Google Scholar]
- Sachdev, K.; Kulshreshtha, D.K. Flavonoids from Dodonaea viscosa. Phytochemistry 1983, 22, 1253–1256. [Google Scholar] [CrossRef]
- Rojas, A.; Cruz, S.; Ponce-monter, H. Smooth muscle relaxing compounds from Dodonaea viscosa. Planta Med. 1996, 62, 154–159. [Google Scholar] [CrossRef]
- Sachdev, K.; Kulshreshtha, D.K. Dodonic acid, a new diterpenoid from Dodonaea viscosa. Planta Med. 1984, 50, 448–449. [Google Scholar] [CrossRef]
- Shugeng, C.; Pegg, B. Antiproliferative triterpenoid saponins of Dodonaea viscosa from the madagascar dry forest. J. Nat. Prod. 2009, 72, 1705–1707. [Google Scholar]
- Khalil, N.M.; Sperotto, J.S.; Manfron, M.P. Anti-inflammatory activity and acute toxicity of Dodonaea viscosa. Fitoterapia 2006, 77, 478–480. [Google Scholar] [CrossRef]
- Getie, M.; Gebre, M.T. Evaluation of the anti-microbial and anti-inflammatory activities of the medicinal plants Dodonaea viscosa, rumex nervosus and ru-mex abyssinicu. Fitoterapia 2003, 74, 139–143. [Google Scholar] [CrossRef]
- Arun, M.; Asha, V.V. Gastroprotective effect of Dodonaea viscosa (L.) on various experimental ulcer models. J. Ethnopharmacol. 2008, 118, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.T.; Lei, C.; You, J.Q.; Zhao, T.; Yu, M.H.; Shi, X.L.; Hu, X.; Hou, A.J. Dimeric clerodane diterpenoids and antiviral constituents of Dodonaea viscosa. Bioorg. Chem. 2021, 112, 104916. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Wang, X.H.; Liu, Y.N.; Zhao, T.; Hu, Z.; Li, J.Y.; Hou, A.J. Clerodane diterpenoids from Dodonaea viscosa and their inhibitory effects on ATP citrate lyase. Phytochemistry 2020, 183, 112614. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fu, L.X.; Fan, L.M.; Qin, X.P.; Ye, M. Isolation and structure identification of dodoneaviscoside A from the seeds of Dodonaea viscosa and its antifeedant activity against Plutella xylostella (L.). Chin. J. Pestic. Sci. 2018, 20, 390–396. [Google Scholar]
- Tuan, S.J.; Li, N.J.; Yeh, C.C.; Tang, L.C.; Chi, H. Effects of green manure cover crops on Spodoptera litura (Lepidoptera: Noctuidae) populations. J. Econ. Entomol. 2014, 107, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Mehmood, R. Monitoring of resistance to new chemistry insecticides in Spodoptera litura (Lepidoptera: Noctuidae) in Pakistan. J. Econ. Entomol. 2015, 108, 1279–1288. [Google Scholar] [CrossRef]
- Su, J.; Lai, T.; Li, J. Susceptibility of field populations of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) in China to chlorantraniliprole and the activities of detoxification enzymes. Crop Prot. 2012, 42, 217–222. [Google Scholar] [CrossRef]
- Li, M.Y.; Gao, X.; Lan, M.X.; Liao, X.B.; Su, F.W.; Fan, L.M.; Zhao, Y.H.; Hao, X.J.; Wu, G.X.; Ding, X. Inhibitory activities of flavonoids from Eupatorium adenophorum against acetylcholinesterase. Pestic. Biochem. Physiol. 2020, 170, 104701. [Google Scholar] [CrossRef]
- Ding, X.; Ouyang, M.A.; Liu, X.; Wang, R.Z. Acetylcholinesterase inhibitory activities of flavonoids from the leaves of Ginkgo biloba against brown planthopper. J. Chem. 2013, 2013, 645086. [Google Scholar] [CrossRef] [Green Version]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lan, M.; Gao, X.; Duan, X.; Li, H.; Yu, H.; Li, J.; Zhao, Y.; Hao, X.; Zhao, Y.; Ding, X.; et al. Nematicidal activity of tirotundin and parthenolide isolated from Tithonia diversifolia and Chrysanthemum parthenium. J. Environ. Sci. Health B 2022, 57, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; van Loon, J.J.A.; Wang, C.-Z. Experience-based behavioral and chemosensory changes in the generalist insect herbivore Helicoverpa armigera exposed to two deterrent plant chemicals. J. Comp. Physiol. A 2010, 196, 791–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.; Liao, C.Y. Behavioural and sensory responses to some neem compounds by Pieris brassicae larvae. Physiol. Entomol. 1995, 20, 134–140. [Google Scholar]
- Mordue, L.; Qiu, Y. Chemoreception of oviposition inhibition terpenoids in the diamondback moth Plutella xylostella. Entomol. Exp. Appl. 1998, 87, 143–155. [Google Scholar]
- Qin, D.Q.; Zhang, P.W.; Zhou, Y.; Liu, B.J.; Xiao, C.X.; Chen, W.B.; Zhang, Z.X. Antifeeding effects of azadirachtin on the fifth instar Spodoptera litura larvae and the analysis of azadirachtin on target sensilla around mouthparts. Arch. Insect Biochem. 2020, 103, e21646. [Google Scholar] [CrossRef]
- Waladde, S.; Hassanali, A.; Ochieng, S. Taste sensilla responses to limonoids, natural insect antifeedants. Int. J. Trop. Insect Sci. 1989, 10, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Messchendorp, L.; Loon, J.; Gols, G. Behavioural and sensory responses to drimane antifeedants in Pieris brassicae larvae. Entomol. Exp. Appl. 1996, 79, 195–202. [Google Scholar] [CrossRef]
- Mullin, C.A.; Chyb, S.; Eichenseer, H.; Hollister, B.; Frazier, J.L. Neuroreceptor mechanisms in insect gustation: A pharmacological approach. J. Insect Physiol. 1994, 40, 913–931. [Google Scholar] [CrossRef]
- Mitchell, B.K. Interaction of alkaloids with galeal chemosensory cells of Colorado potato beetle. J. Chem. Ecol. 1987, 13, 2009–2022. [Google Scholar] [CrossRef]
- Gonzalez-Coloma, A.; Reina, M.; Cabrera, R.; Castañera, P.; Gutierrez, C. Antifeedant and toxic effects of sesquiterpenes from Senecio palmensis to colorado potato beetle. J. Chem. Ecol. 1995, 21, 1255–1270. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Nur, F.A.; Ma, J.Y.; Wang, J.G.; Cao, C.W. Effects of poplar secondary metabolites on performance and detoxification enzyme activity of Lymantria dispar. Comp. Biochem. Phys. C 2019, 225, 108587. [Google Scholar] [CrossRef] [PubMed]
- Snyder, M.J.; Glendinning, J.I. Causal connection between detoxification enzyme activity and consumption of a toxic plant compound. J. Comp. Physiol. A 1996, 179, 255–261. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (μg/mL) | No. of Tested Insects | Feeding Area (mm2) | Inhibition Rate (%) |
|---|---|---|---|
| 0 | 10 | 985.36 ± 86.79 a | |
| 500 | 10 | 687.68 ± 83.06 b | 30.21 |
| 1625 | 10 | 585.14 ± 45.89 c | 40.62 |
| 2750 | 10 | 450.36 ± 62.50 d | 54.33 |
| 3875 | 10 | 271.27 ± 38.30 e | 72.47 |
| 5000 | 10 | 149.38 ± 7.50 f | 84.84 |
| Azadirachtin | 10 | 302.70 ± 64.83 e | 69.26 |
| Concentration | Developmental Period/d | ||||
|---|---|---|---|---|---|
| 4th–6th Instar Larva | Female Pupae | Male Pupae | Female Adult | Male Adult | |
| CK | 12.10 ± 0.067 c,d | 12.73 ± 0.014 a | 11.11 ± 0.070 a | 6.84 ± 0.076 a | 3.88 ± 0.101 a |
| AFC10 | 12.27 ± 0.094 b,c | 12.75 ± 0.063 a | 11.07 ± 0.047 a | 6.79 ± 0.083 a | 3.97 ± 0.056 a |
| AFC30 | 12.42 ± 0.268 a,b | 12.83 ± 0.085 a | 11.07 ± 0.051 a | 6.78 ± 0.047 a | 3.64 ± 0.050 b |
| AFC50 | 12.65 ± 0.106 a | 12.73 ± 0.065 a | 10.75 ± 0.084 b | 5.59 ± 0.076 b | 3.00 ± 0.035 c |
| Concentration | Pupation Rate | Emergence Rate |
|---|---|---|
| CK | 100.00 ± 0.00 a | 88.00 ± 2.00 a |
| AFC10 | 98.00 ± 2.00 a | 83.56 ± 2.65 a |
| AFC30 | 84.00 ± 5.10 b | 69.12 ± 2.12 b |
| AFC50 | 76.00 ± 5.10 b | 70.68 ± 2.76 b |
| Concentration (μg/mL) | No. of Tested Insects | Feeding Area (mm2) | Inhibition Rate (%) |
|---|---|---|---|
| 0 | 10 | 985.36 ± 86.36 a | |
| 50 | 10 | 658.12 ± 89.57 b | 23.52 |
| 100 | 10 | 565.31 ± 45.93 c | 32.35 |
| 200 | 10 | 499.28 ± 55.33 c | 39.57 |
| 400 | 10 | 359.81 ± 39.11 d | 52.63 |
| 800 | 10 | 292.06 ± 20.95 d | 70.36 |
| Azadirachtin | 10 | 302.70 ± 64.83 d | 69.26 |
| Concentration | Developmental Period/d | ||||
|---|---|---|---|---|---|
| 4th–6th Instar Larva | Female Pupae | Male Pupae | Female Adult | Male Adult | |
| CK | 12.09 ± 0.014 c | 12.68 ± 0.014 a | 11.11 ± 0.010 a | 6.84 ± 0.011 a | 3.88 ± 0.016 a |
| AFC10 | 12.12 ± 0.012 c | 12.71 ± 0.015 a | 11.10 ± 0.012 a | 6.79 ± 0.013 a | 3.88 ± 0.011 a |
| AFC30 | 12.48 ± 0.008 b | 12.70 ± 0.019 a | 10.92 ± 0.014 b | 6.08 ± 0.012 b | 3.90 ± 0.012 a |
| AFC50 | 13.04 ± 0.011 a | 12.35 ± 0.018 b | 10.85 ± 0.016 c | 5.51 ± 0.017 c | 3.87 ± 0.016 a |
| Concentration | Pupation Rate | Emergence Rate |
|---|---|---|
| CK | 100.00 ± 0.000 a | 96.00 ± 2.449 a |
| AFC10 | 98.00 ± 2.000 a | 94.00 ± 4.000 a |
| AFC30 | 90.00 ± 3.162 b | 79.89 ± 5.512 b |
| AFC50 | 78.00 ± 3.741 c | 76.98 ± 2.403 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Li, J.; Wu, G.; Tang, Q.; Duan, X.; Liu, Q.; Lan, M.; Zhao, Y.; Hao, X.; Qin, X.; et al. Antifeedant Mechanism of Dodonaea viscosa Saponin A Isolated from the Seeds of Dodonaea viscosa. Molecules 2022, 27, 4464. https://doi.org/10.3390/molecules27144464
Yu H, Li J, Wu G, Tang Q, Duan X, Liu Q, Lan M, Zhao Y, Hao X, Qin X, et al. Antifeedant Mechanism of Dodonaea viscosa Saponin A Isolated from the Seeds of Dodonaea viscosa. Molecules. 2022; 27(14):4464. https://doi.org/10.3390/molecules27144464
Chicago/Turabian StyleYu, Hang, Jinliang Li, Guoxing Wu, Qingbo Tang, Xiuan Duan, Quanjun Liu, Mingxian Lan, Yuhan Zhao, Xiaojiang Hao, Xiaoping Qin, and et al. 2022. "Antifeedant Mechanism of Dodonaea viscosa Saponin A Isolated from the Seeds of Dodonaea viscosa" Molecules 27, no. 14: 4464. https://doi.org/10.3390/molecules27144464
APA StyleYu, H., Li, J., Wu, G., Tang, Q., Duan, X., Liu, Q., Lan, M., Zhao, Y., Hao, X., Qin, X., & Ding, X. (2022). Antifeedant Mechanism of Dodonaea viscosa Saponin A Isolated from the Seeds of Dodonaea viscosa. Molecules, 27(14), 4464. https://doi.org/10.3390/molecules27144464