
67-kDa Laminin Receptor-Mediated Cellular Sensing System of Green Tea Polyphenol EGCG and Functional Food Pairing


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
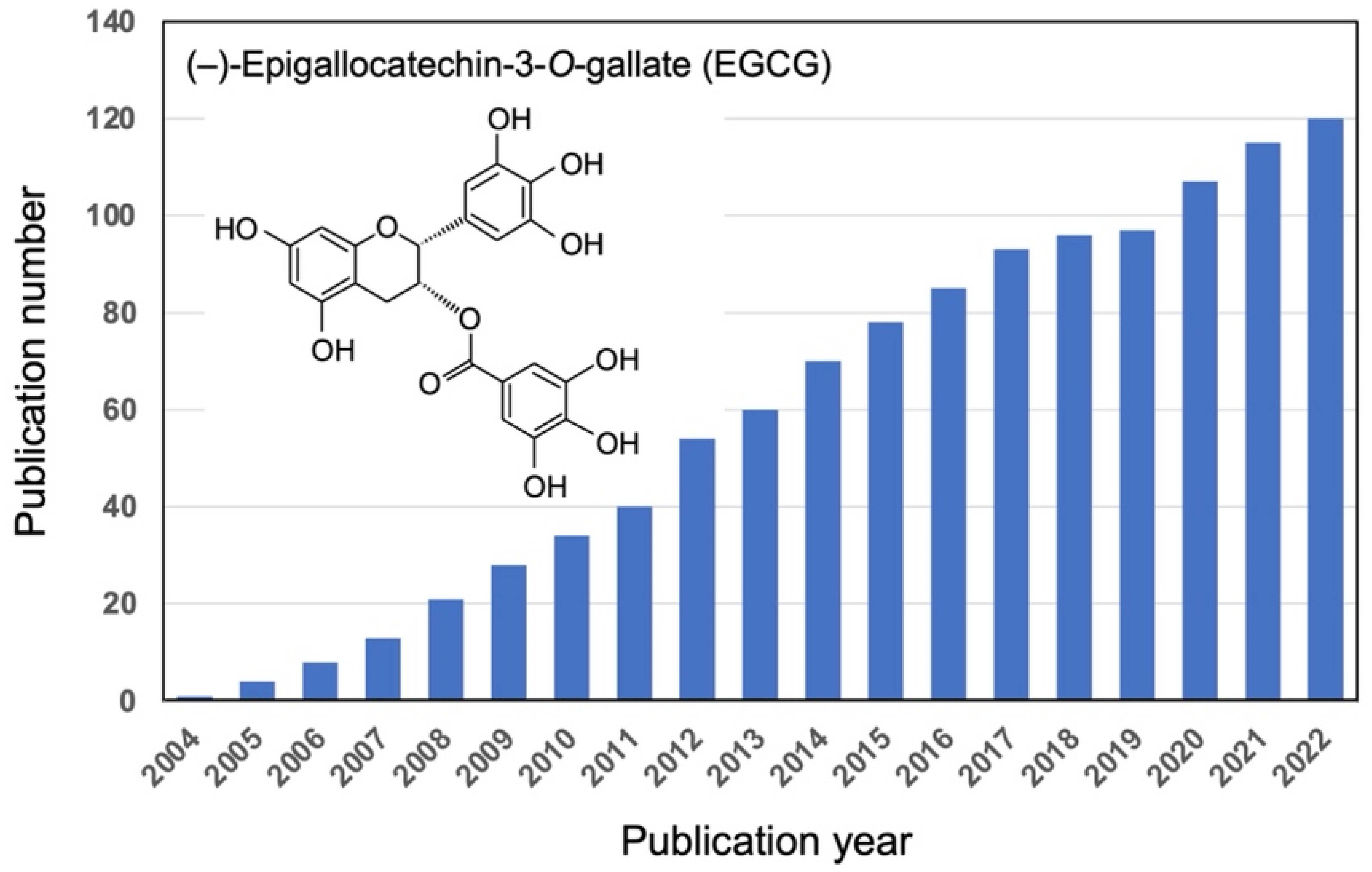
:1. Introduction
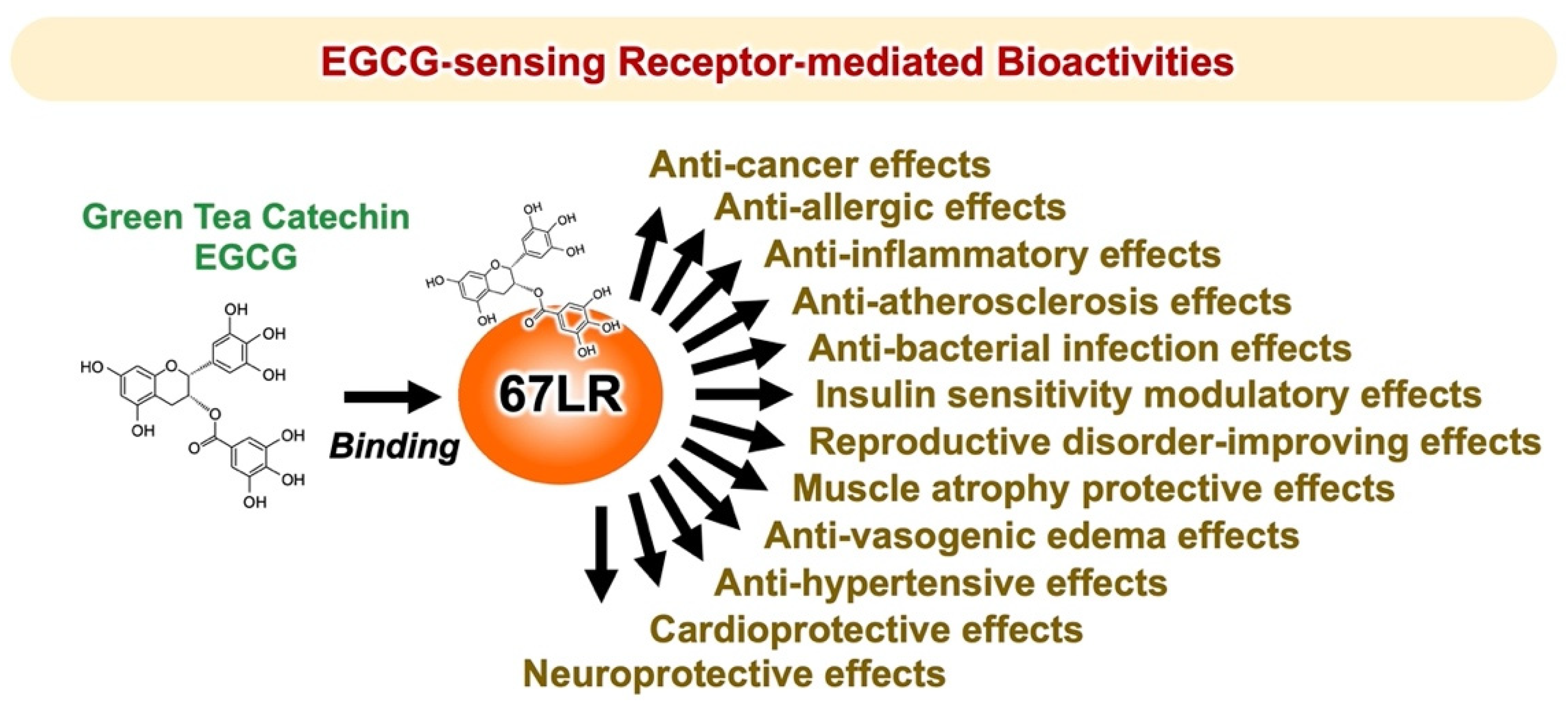
2. Green Tea Catechin EGCG-Sensing Receptor
2.1. Identification of the Green Tea Polyphenol EGCG-Sensing Receptor
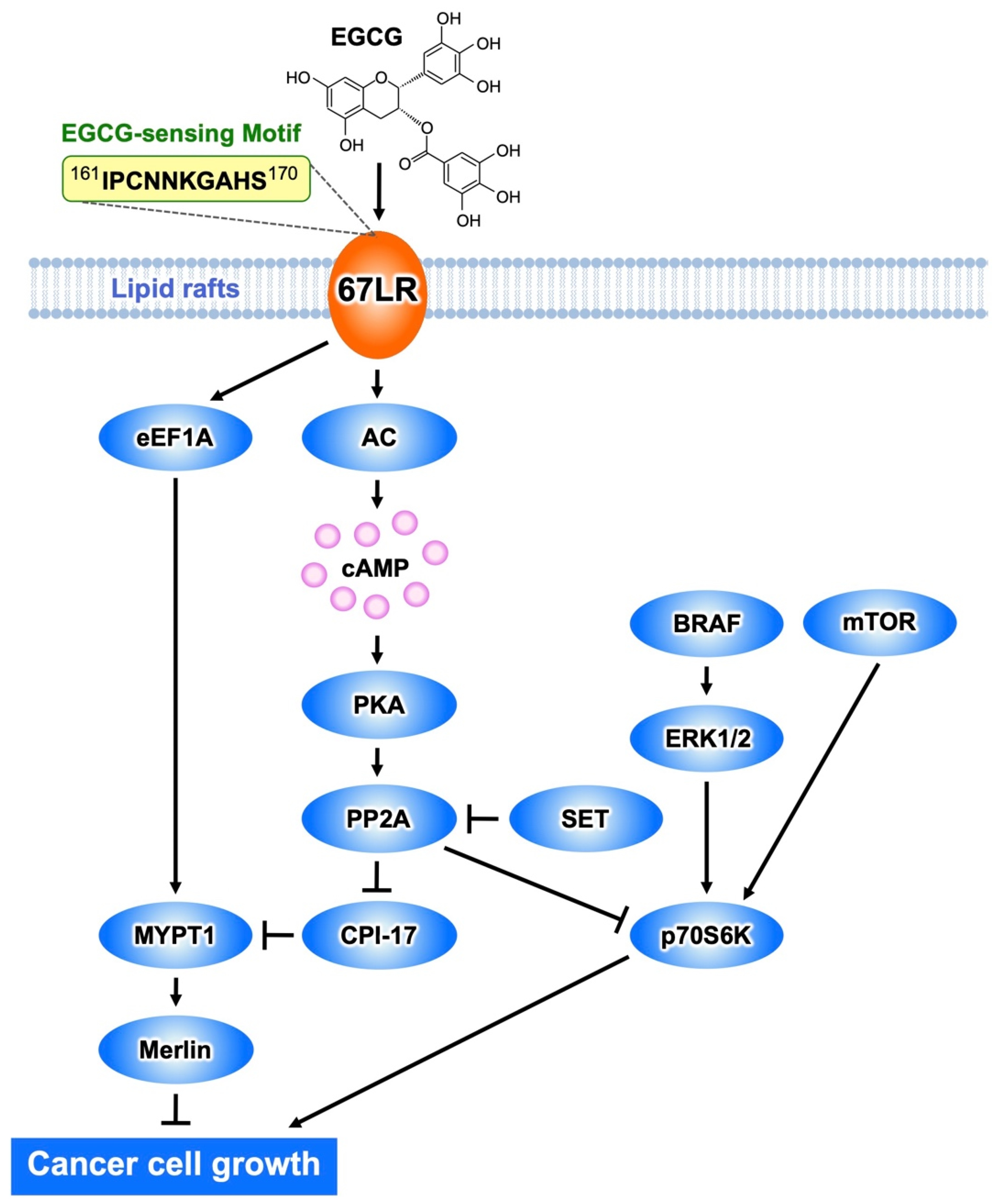
2.2. Inhibitory Actions of EGCG on Cancer Cell Growth Mediated by 67LR
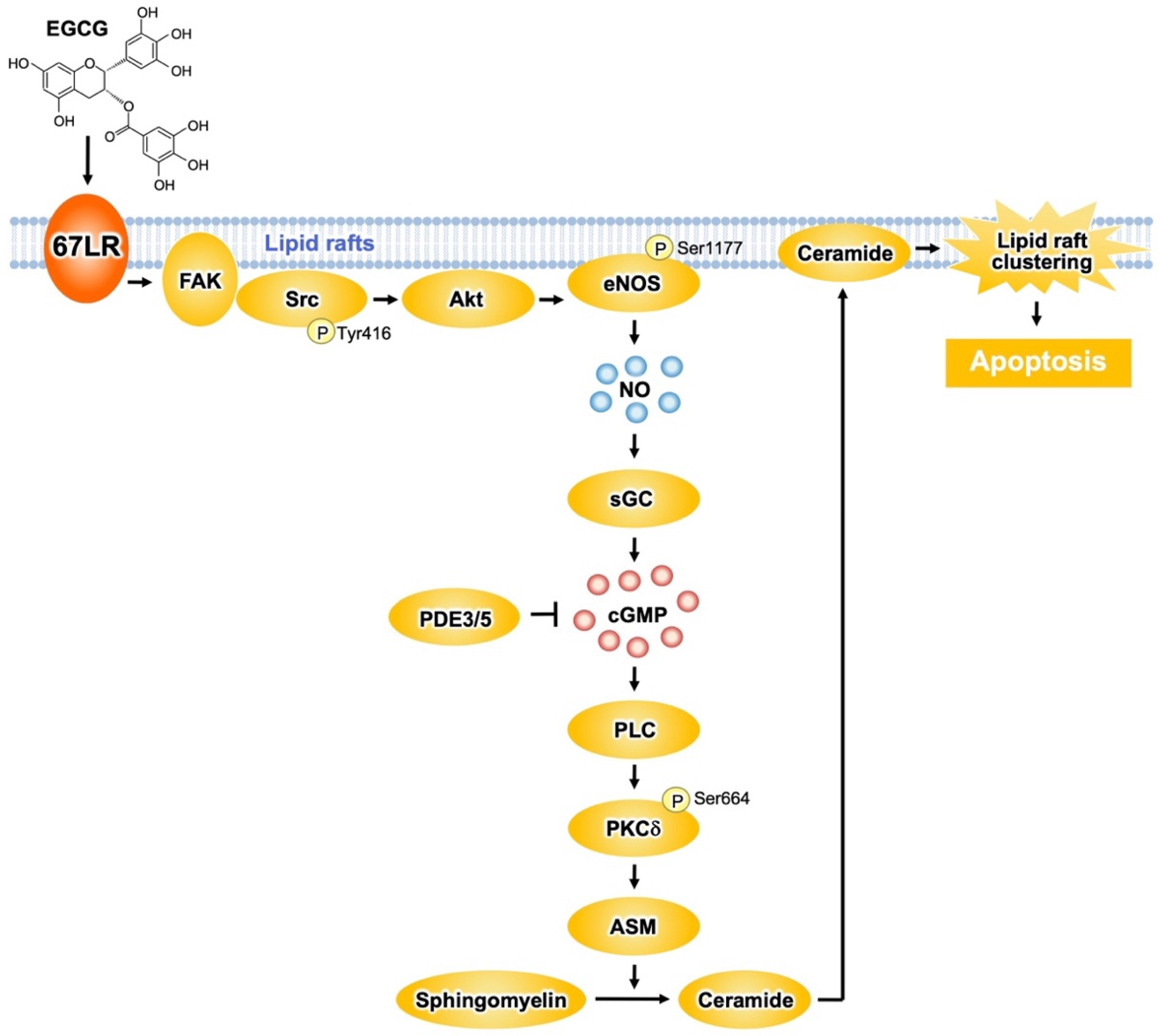
2.3. Cancer Cell Killing Effects of EGCG Mediated by 67LR
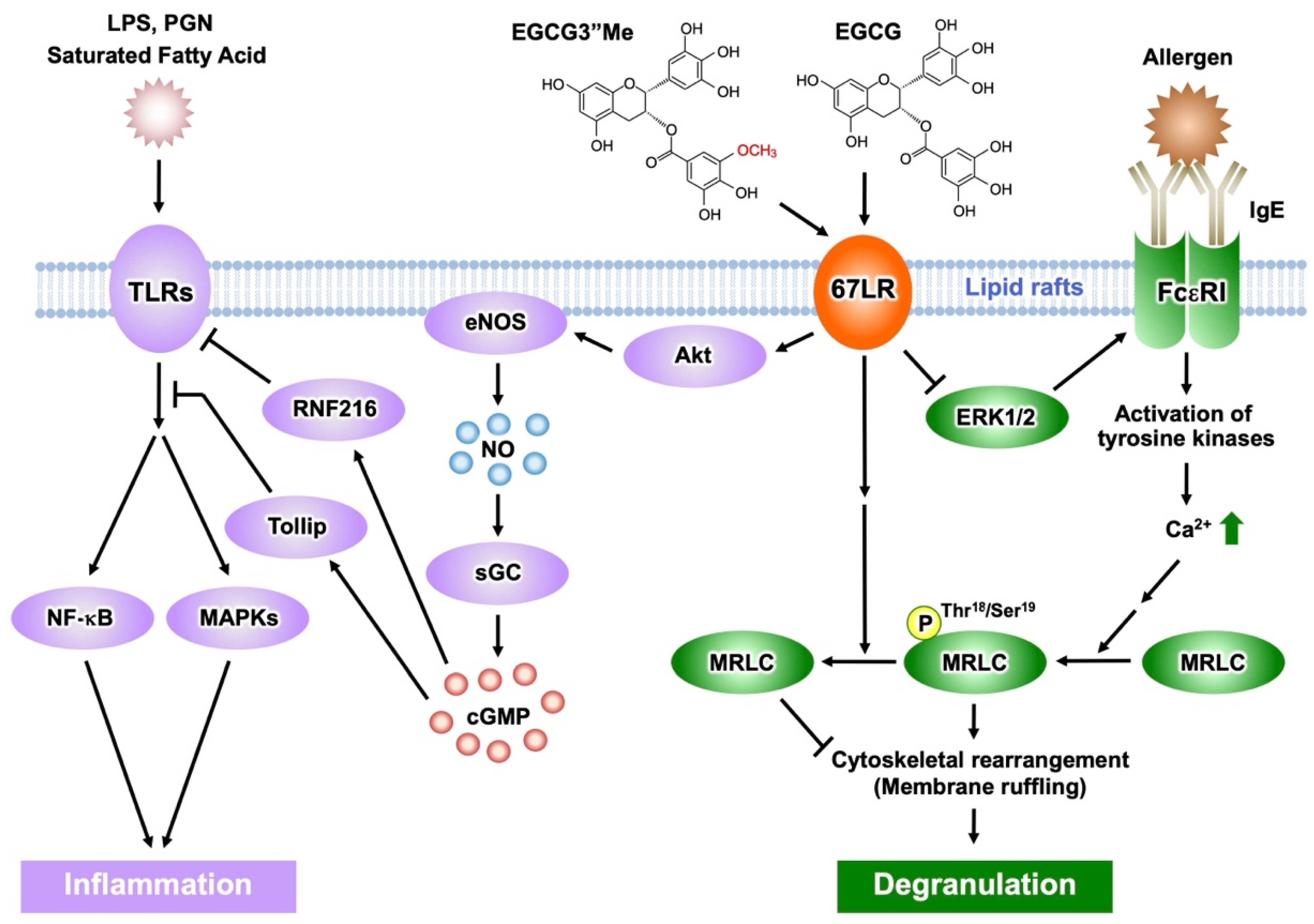
2.4. Anti-Inflammatory and Anti-Allergic Actions of EGCG Mediated by 67LR
2.5. MicroRNA-Mediated Anti-Cancer Effect of EGCG via 67LR
3. Potentiation of EGCG Activity by Modulating the 67LR-Dependent EGCG-Sensing Pathways
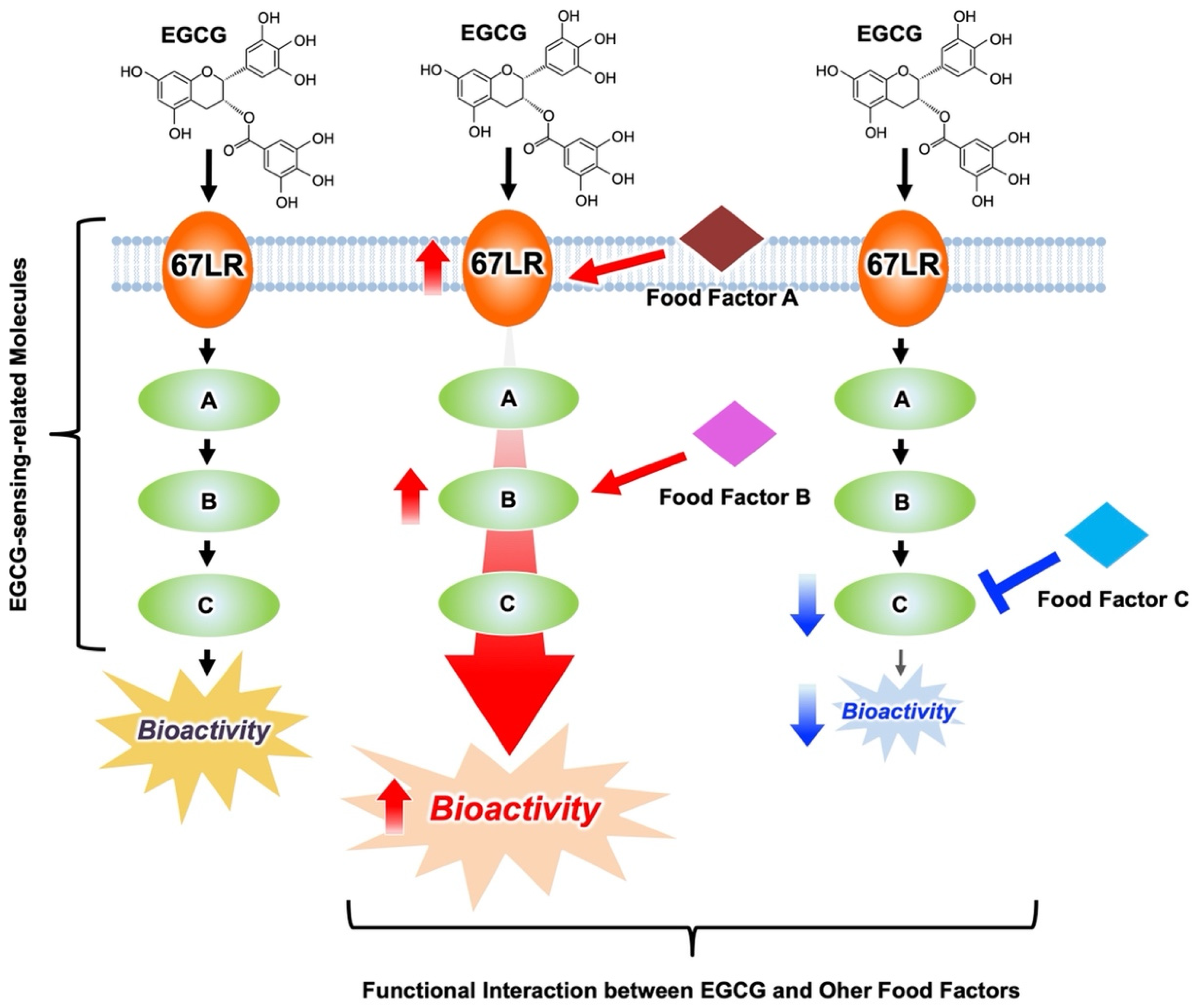
3.1. Anti-Cancer Effects of EGCG Are Potentiated by Modulation of Several EGCG-Sensing-Related Molecules
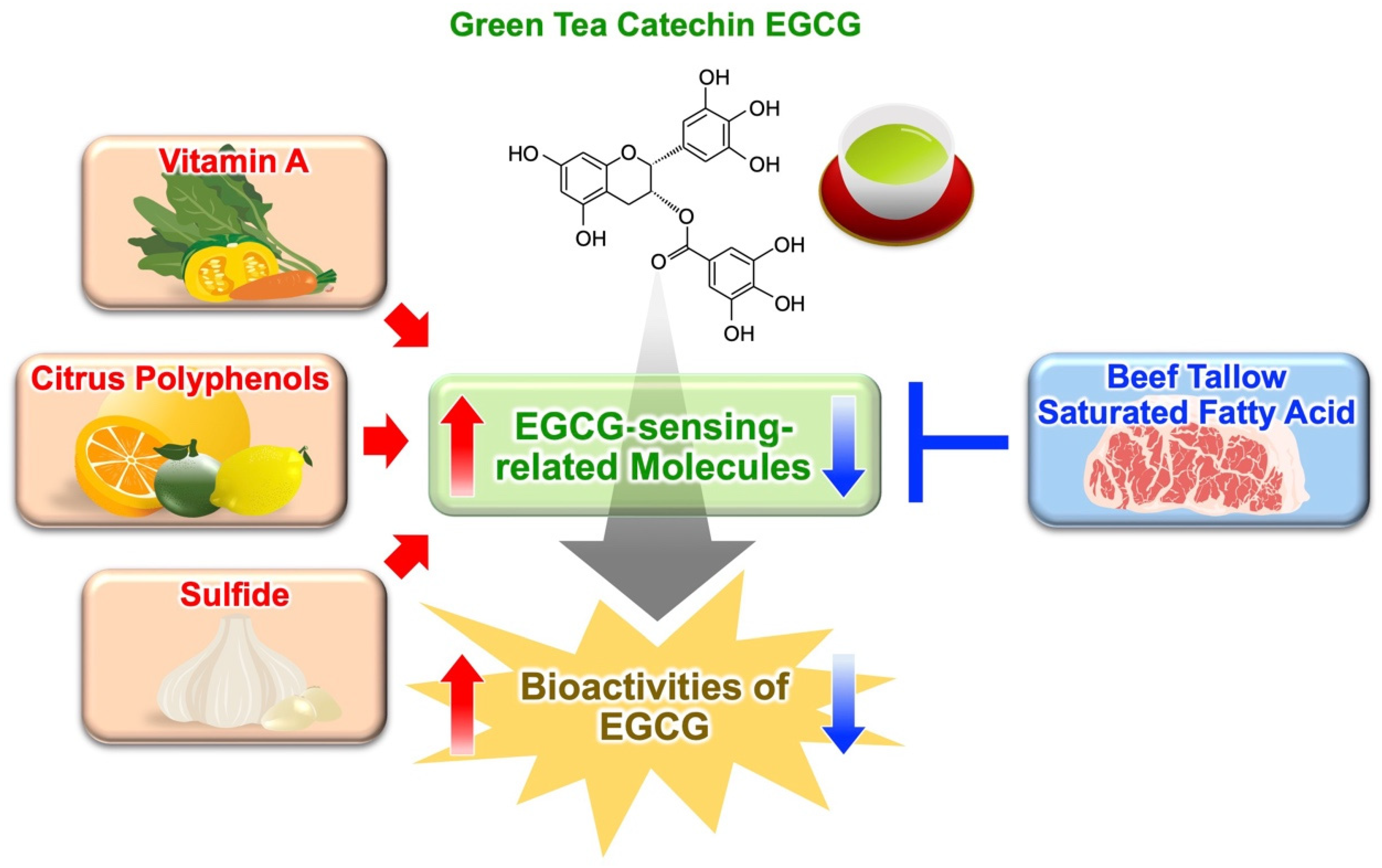
3.2. Citrus Polyphenols Potentiate Bioactivites of EGCG
3.3. Sulfur-Containing Food Factor Potentiates Bioactivites of EGCG
3.4. Fatty Acids Modulate Anti-Obesity Effect of Green Tea Extract
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| 67LR | 67-kDa laminin receptor |
| AC | adenylate cyclase |
| AML | acute myeloid leukemia |
| ASM | acid sphingomyelinase |
| ATRA | all-trans-retinoic acid |
| BMI | body mass index |
| b.w. | body weight |
| C | (−)-catechin |
| CG | (−)-catechin-3-O-gallate |
| Cmax | maximum plasma concentration |
| CPI17 | C-kinase potentiated protein phosphatase-1 inhibitor protein of 17-kDa |
| DADS | diallyl disulfide |
| DAG | diacylglycerol |
| DATS | diallyl trisulfide |
| EC | (−)-epicatechin |
| ECG | (−)-epicatechin-3-O-gallate |
| eEF1A | eukaryotic elongation factor 1A |
| EGC | (−)-epigallocatechin |
| EGCG | (−)-epigallocatechin-3-O-gallate |
| EGCG3″Me | (−)-epigallocatechin-3-O-(3-O-methyl) gallate |
| EGCG4″Me | (−)-epigallpcatechin-3-O-(4-O-methyl) gallate |
| EGCG4′4″diMe | (−)-4′-O-methyl-epigallocatechin-3-O-(4-O-methyl) gallate |
| Elf-1 | E74-like ETS factor 1 |
| eNOS | endothelial nitric oxide synthase |
| ER | estrogen receptor |
| ERK1/2 | extracellular signal-regulated kinase1/2 |
| FAK | focal adhesion kinase |
| FcεRI | high-affinity IgE receptor |
| GCG | (−)-gallocatechin-3-O-gallate |
| GT-gH | green tea combined with α-glucosyl hesperidin |
| HDL | high-density lipoprotein |
| HF/HS | high-fat and high-sucrose diet |
| HF/HS + Eri | HF/HS with eriodictyol diet |
| HF/HS + GT | HF/HS with green tea extract diet |
| HF/HS + GT + Eri | HF/HS with green tea extract and eriodictyol diet |
| HIF-1 | hypoxia-inducible factor 1 |
| HMG-CoA | 3-hydroxy-3-methylglutaryl-coenzyme A |
| HMG-CR | HMG-CoA reductase |
| HMG-CS | HMG-CoA synthase |
| HMGA2 | high-mobility group A2 |
| Ig | immunoglobulin |
| IL | interleukin |
| IL-4R | IL-4 receptor |
| Kd | dissociation constant |
| LDL | low-density lipoprotein |
| LDLR | LDL receptor |
| LPS | lipopolysaccharide |
| MAPKs | mitogen-activated protein kinases |
| miRNA/miR | microRNA |
| mTOR | mammalian target of rapamycin |
| MM | multiple myeloma |
| MRLC | myosin regulatory light chain |
| mTOR | mammalian target of rapamycin |
| MuRF1 | muscle-specific RING-finger protein 1 |
| MYPT1 | myosin phosphatase target subunit 1 |
| NF-κB | nuclear factor kappa B |
| p70S6K | p70S6 kinase |
| PAPD5 | PAP-associated domain containing 5 |
| PB1 | procyanidin B1 |
| PB2 | procyanidin B2 |
| PBMCs | peripheral blood mononuclear cells |
| PC1 | procyanidin C1 |
| PCA | passive cutaneous anaphylaxis |
| PDEs | phosphodiesterases |
| PGN | peptidoglycan |
| PKA | protein kinase A |
| PKCδ | protein kinase Cδ |
| PLC | phospholipase C |
| PP2A | protein phosphatase 2A |
| PPAR | peroxisome proliferator-activated receptor |
| RAR | retinoic acid receptor |
| RNF216 | E3 ubiquitin-protein ring finger protein 216 |
| RXRs | retinoid X receptors |
| SET | Suvar3–9, enhancer-of-zeste, trithorax |
| sGC | soluble guanylate cyclase |
| shRNA | short hairpin RNA |
| SREBP-1 | sterol regulatory element-binding protein-1 |
| TLR4 | Toll-like receptor 4 |
| Tollip | Toll-interacting protein |
References
- Tachibana, H.; Koga, K.; Fujimura, Y.; Yamada, K. A Receptor for Green Tea Polyphenol EGCG. Nat. Struct. Mol. Biol. 2004, 11, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kishikawa, M.; Kumazoe, M.; Yamada, K.; Tachibana, H. Vitamin A Enhances Antitumor Effect of a Green Tea Polyphenol on Melanoma by Upregulating the Polyphenol Sensing Molecule 67-KDa Laminin Receptor. PLoS ONE 2010, 5, e11051. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Huang, Y.; Umeda, D.; Yamada, S.; Yamashita, S.; Kumazoe, M.; Kim, Y.; Murata, M.; Yamada, K.; Tachibana, H. 67-KDa Laminin Receptor-Dependent Protein Phosphatase 2A (PP2A) Activation Elicits Melanoma-Specific Antitumor Activity Overcoming Drug Resistance. J. Biol. Chem. 2014, 289, 32671–32681. [Google Scholar] [CrossRef] [PubMed]
- Kumazoe, M.; Sugihara, K.; Tsukamoto, S.; Huang, Y.; Tsurudome, Y.; Suzuki, T.; Suemasu, Y.; Ueda, N.; Yamashita, S.; Kim, Y.; et al. 67-KDa Laminin Receptor Increases CGMP to Induce Cancer-Selective Apoptosis. J. Clin. Investig. 2013, 123, 787–799. [Google Scholar] [CrossRef]
- Hong Byun, E.; Fujimura, Y.; Yamada, K.; Tachibana, H. TLR4 Signaling Inhibitory Pathway Induced by Green Tea Polyphenol Epigallocatechin-3-Gallate through 67-KDa Laminin Receptor. J. Immunol. 2010, 185, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Kumazoe, M.; Nakamura, Y.; Yamashita, M.; Suzuki, T.; Takamatsu, K.; Huang, Y.; Bae, J.; Yamashita, S.; Murata, M.; Yamada, S.; et al. Green Tea Polyphenol Epigallocatechin-3-Gallate Suppresses Toll-like Receptor 4 Expression via up-Regulation of E3 Ubiquitin-Protein Ligase RNF216. J. Biol. Chem. 2017, 292, 4077–4088. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Kumazoe, M.; Nakamura, Y.; Won, S.; Bae, J.; Yamashita, S.; Tachibana, H. The Combination of Green Tea Extract and Eriodictyol Inhibited High-Fat/High-Sucrose Diet-Induced Cholesterol Upregulation Is Accompanied by Suppression of Cholesterol Synthesis Enzymes. J. Nutr. Sci. Vitaminol. 2016, 62, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Shimizu, Y.; Marugame, Y.; Nezu, A.; Fujino, K.; Yamada, S.; Kumazoe, M.; Fujimura, Y.; Tachibana, H. EGCG Down-Regulates MuRF1 Expression through 67-KDa Laminin Receptor and the Receptor Signaling Is Amplified by Eriodictyol. J. Nat. Med. 2020, 74, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Kumazoe, M.; Fujimura, Y.; Hidaka, S.; Kim, Y.; Murayama, K.; Takai, M.; Huang, Y.; Yamashita, S.; Murata, M.; Miura, D.; et al. Metabolic Profiling-Based Data-Mining for an Effective Chemical Combination to Induce Apoptosis of Cancer Cells. Sci. Rep. 2015, 5, 9474. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y.; Fujino, K.; Yoshimoto, T.; Nezu, A.; Marugame, Y.; Bae, J.; Kumazoe, M.; Tachibana, H. Eriodictyol-Amplified 67-KDa Laminin Receptor Signaling Potentiates the Antiallergic Effect of O-Methylated Catechin. J. Nat. Prod. 2021, 84, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Kuriyama, S.; Li, Q.; Nagai, M.; Hozawa, A.; Nishino, Y.; Tsuji, I. Citrus Consumption and Cancer Incidence: The Ohsaki Cohort Study. Int. J. Cancer 2010, 127, 1913–1922. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, R.; Yamamoto, M.; Kumazoe, M.; Fujimura, Y.; Yonekura, M.; Shimamoto, Y.; Nakasone, A.; Kondo, S.; Hattori, H.; Haseda, A.; et al. The Combined Effect of Green Tea and α-Glucosyl Hesperidin in Preventing Obesity: A Randomized Placebo-Controlled Clinical Trial. Sci. Rep. 2021, 11, 19067. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Zhu, Q.Y.; Tsang, D.; Huang, Y. Degradation of Green Tea Catechins in Tea Drinks. J. Agric. Food Chem. 2001, 49, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Howells, L.M.; Moiseeva, E.P.; Neal, C.P.; Foreman, B.E.; Andreadi, C.K.; Sun, Y.Y.; Hudson, E.A.; Manson, M.M. Predicting the Physiological Relevance of in Vitro Cancer Preventive Activities of Phytochemicals. Acta Pharmacol. Sin. 2007, 28, 1274–1304. [Google Scholar] [CrossRef]
- Yang, C.S.; Wang, X.; Lu, G.; Picinich, S.C. Cancer Prevention by Tea: Animal Studies, Molecular Mechanisms and Human Relevance. Nat. Rev. Cancer 2009, 9, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Negri, A.; Naponelli, V.; Rizzi, F.; Bettuzzi, S. Molecular Targets of Epigallocatechin—Gallate (EGCG): A Special Focus on Signal Transduction and Cancer. Nutrients 2018, 10, 1936. [Google Scholar] [CrossRef]
- Fujimura, Y.; Sumida, M.; Sugihara, K.; Tsukamoto, S.; Yamada, K.; Tachibana, H. Green Tea Polyphenol EGCG Sensing Motif on the 67-KDa Laminin Receptor. PLoS ONE 2012, 7, e37942. [Google Scholar] [CrossRef]
- Fujimura, Y.; Yamada, K.; Tachibana, H. A Lipid Raft-Associated 67 KDa Laminin Receptor Mediates Suppressive Effect of Epigallocatechin-3-O-Gallate on FcεRI Expression. Biochem. Biophys. Res. Commun. 2005, 336, 674–681. [Google Scholar] [CrossRef]
- Fujimura, Y.; Tachibana, H.; Yamada, K. Lipid Raft-Associated Catechin Suppresses the FcεRI Expression by Inhibiting Phosphorylation of the Extracellular Signal-Regulated Kinase1/2. FEBS Lett. 2004, 556, 204–210. [Google Scholar] [CrossRef]
- Umeda, D.; Yano, S.; Yamada, K.; Tachibana, H. Green Tea Polyphenol Epigallocatechin-3-Gallate Signaling Pathway through 67-KDa Laminin Receptor. J. Biol. Chem. 2008, 283, 3050–3058. [Google Scholar] [CrossRef]
- Mafune, K.; Ravikumar, T.S. Anti-Sense RNA of 32-KDa Laminin-Binding Protein Inhibits Attachment and Invasion of a Human Colon Carcinoma Cell Line. J. Surg. Res. 1992, 52, 340–346. [Google Scholar] [CrossRef]
- Gauczynski, S.; Peyrin, J.M.; Haïk, S.; Leucht, C.; Hundt, C.; Rieger, R.; Krasemann, S.; Deslys, J.P.; Dormont, D.; Lasmézas, C.I.; et al. The 37-KDa/67-KDa Laminin Receptor Acts as the Cell-Surface Receptor for the Cellular Prion Protein. EMBO J. 2001, 20, 5863–5875. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.S.; Kuhn, R.J.; Strauss, E.G.; Ou, S.; Strauss, J.H. High-Affinity Laminin Receptor Is a Receptor for Sindbis Virus in Mammalian Cells. J. Virol. 1992, 66, 4992–5001. [Google Scholar] [CrossRef]
- Thepparit, C.; Smith, D.R. Serotype-Specific Entry of Dengue Virus into Liver Cells: Identification of the 37-Kilodalton/67-Kilodalton High-Affinity Laminin Receptor as a Dengue Virus Serotype 1 Receptor. J. Virol. 2004, 78, 12647–12656. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y.; Umeda, D.; Yamada, K.; Tachibana, H. The Impact of the 67 KDa Laminin Receptor on Both Cell-Surface Binding and Anti-Allergic Action of Tea Catechins. Arch. Biochem. Biophys. 2008, 476, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Kubo, T.; Miyase, T.; Tanino, S.; Yoshimoto, M.; Sano, M.; Yamamoto-Maeda, M.; Yamada, K. Identification of an Inhibitor for Interleukin 4-Induced ε Germline Transcription and Antigen-Specific IgE Production In Vivo. Biochem. Biophys. Res. Commun. 2001, 280, 53–60. [Google Scholar] [CrossRef]
- Kim, Y.H.; Ninomiya, Y.; Yamashita, S.; Kumazoe, M.; Huang, Y.; Nakahara, K.; Won, Y.S.; Murata, M.; Fujimura, Y.; Yamada, K.; et al. IL-4 Receptor α in Non-Lipid Rafts Is the Target Molecule of Strictinin in Inhibiting STAT6 Activation. Biochem. Biophys. Res. Commun. 2014, 450, 824–830. [Google Scholar] [CrossRef]
- Huang, Y.; Sumida, M.; Kumazoe, M.; Sugihara, K.; Suemasu, Y.; Yamada, S.; Yamashita, S.; Miyakawa, J.; Takahashi, T.; Tanaka, H.; et al. Oligomer Formation of a Tea Polyphenol, EGCG, on Its Sensing Molecule 67 KDa Laminin Receptor. Chem. Commun. 2017, 53, 1941–1944. [Google Scholar] [CrossRef]
- Gossé, F.; Guyot, S.; Roussi, S.; Lobstein, A.; Fischer, B.; Seiler, N.; Raul, F. Chemopreventive Properties of Apple Procyanidins on Human Colon Cancer-Derived Metastatic SW620 Cells and in a Rat Model of Colon Carcinogenesis. Carcinogenesis 2005, 26, 1291–1295. [Google Scholar] [CrossRef]
- Miura, T.; Chiba, M.; Kasai, K.; Nozaka, H.; Nakamura, T.; Shoji, T.; Kanda, T.; Ohtake, Y.; Sato, T. Apple Procyanidins Induce Tumor Cell Apoptosis through Mitochondrial Pathway Activation of Caspase-3. Carcinogenesis 2008, 29, 585–593. [Google Scholar] [CrossRef]
- Bae, J.; Kumazoe, M.; Murata, K.; Fujimura, Y.; Tachibana, H. procyanidin c1 inhibits melanoma cell growth by activating 67-kda laminin receptor signaling. Mol. Nutr. Food Res. 2020, 64, 1900986. [Google Scholar] [CrossRef] [PubMed]
- Salama, A.K.S.; Flaherty, K.T. Braf in melanoma: Current strategies and Future Directions. Clin. Cancer Res. 2013, 19, 4326–4334. [Google Scholar] [CrossRef] [PubMed]
- Chin, L.; Garraway, L.A.; Fisher, D.E. Malignant Melanoma: Genetics and Therapeutics in the Genomic Era. Genes Dev. 2006, 20, 2149–2182. [Google Scholar] [CrossRef] [PubMed]
- Umeda, D.; Tachibana, H.; Yamada, K. Epigallocatechin-3-O-Gallate Disrupts Stress Fibers and the Contractile Ring by Reducing Myosin Regulatory Light Chain Phosphorylation Mediated through the Target Molecule 67 KDa Laminin Receptor. Biochem. Biophys. Res. Commun. 2005, 333, 628–635. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Huang, Y.; Kumazoe, M.; Lesnick, C.; Yamada, S.; Ueda, N.; Suzuki, T.; Yamashita, S.; Kim, Y.H.; Fujimura, Y.; et al. Sphingosine Kinase-1 Protects Multiple Myeloma from Apoptosis Driven by Cancer-Specific Inhibition of RTKs. Mol. Cancer Ther. 2015, 14, 2303–2312. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Sun, Z.; Han, C.; Chen, J. The Chemopreventive Effects of Tea on Human Oral Precancerous Mucosa Lesions. Proc. Soc. Exp. Biol. Med. 1999, 220, 218–224. [Google Scholar] [PubMed]
- Shimizu, M.; Fukutomi, Y.; Ninomiya, M.; Nagura, K.; Kato, T.; Araki, H.; Suganuma, M.; Fujiki, H.; Moriwaki, H. Green Tea Extracts for the Prevention of Metachronous Colorectal Adenomas: A Pilot Study. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3020–3025. [Google Scholar] [CrossRef] [PubMed]
- Bettuzzi, S.; Brausi, M.; Rizzi, F.; Castagnetti, G.; Peracchia, G.; Corti, A. Chemoprevention of Human Prostate Cancer by Oral Administration of Green Tea Catechins in Volunteers with High-Grade Prostate Intraepithelial Neoplasia: A Preliminary Report from a One-Year Proof-of-Principle Study. Cancer Res. 2006, 66, 1234–1240. [Google Scholar] [CrossRef]
- Shanafelt, T.D.; Call, T.G.; Zent, C.S.; Leis, J.F.; Laplant, B.; Bowen, D.A.; Roos, M.; Laumann, K.; Ghosh, A.K.; Lesnick, C.; et al. Phase 2 Trial of Daily, Oral Polyphenon e in Patients with Asymptomatic, Rai Stage 0 to II Chronic Lymphocytic Leukemia. Cancer 2013, 119, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Shammas, M.A.; Neri, P.; Koley, H.; Batchu, R.B.; Bertheau, R.C.; Munshi, V.; Prabhala, R.; Fulciniti, M.; Yu, T.T.; Treon, S.P.; et al. Specific Killing of Multiple Myeloma Cells by (−)-Epigallocatechin-3- Gallate Extracted from Green Tea: Biologic Activity and Therapeutic Implications. Blood 2006, 108, 2804–2810. [Google Scholar] [CrossRef]
- Kumazoe, M.; Kim, Y.; Bae, J.; Takai, M.; Murata, M.; Suemasu, Y.; Sugihara, K.; Yamashita, S.; Tsukamoto, S.; Huang, Y.; et al. Phosphodiesterase 5 Inhibitor Acts as a Potent Agent Sensitizing Acute Myeloid Leukemia Cells to 67-KDa Laminin Receptor-Dependent Apoptosis. FEBS Lett. 2013, 587, 3052–3057. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Hirotsu, K.; Kumazoe, M.; Goto, Y.; Sugihara, K.; Suda, T.; Tsurudome, Y.; Suzuki, T.; Yamashita, S.; Kim, Y.; et al. Green Tea Polyphenol EGCG Induces Lipid-Raft Clustering and Apoptotic Cell Death by Activating Protein Kinase Cδ and Acid Sphingomyelinase through a 67 KDa Laminin Receptor in Multiple Myeloma Cells. Biochem. J. 2012, 443, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Gloe, T.; Riedmayr, S.; Sohn, H.Y.; Pohl, U. The 67-KDa Laminin-Binding Protein Is Involved in Shear Stress-Dependent Endothelial Nitric-Oxide Synthase Expression. J. Biol. Chem. 1999, 274, 15996–16002. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Kumazoe, M.; Takeuchi, C.; Hidaka, S.; Fujimura, Y.; Tachibana, H. Epigallocatechin-3-O-Gallate Induces Acid Sphingomyelinase Activation through Activation of Phospholipase C. Biochem. Biophys. Res. Commun. 2019, 520, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Kumazoe, M.; Kadomatsu, M.; Bae, J.; Otsuka, Y.; Fujimura, Y.; Tachibana, H. Src Mediates Epigallocatechin-3-O-Gallate-Elicited Acid Sphingomyelinase Activation. Molecules 2020, 25, 5481. [Google Scholar] [CrossRef]
- Li, W.; Ashok, M.; Li, J.; Yang, H.; Sama, A.E.; Wang, H. A Major Ingredient of Green Tea Rescues Mice from Lethal Sepsis Partly by Inhibiting HMGB1. PLoS ONE 2007, 2, e1153. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. TLR Signaling Pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Byun, E.H.; Omura, T.; Yamada, K.; Tachibana, H. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits TLR2 Signaling Induced by Peptidoglycan through the Polyphenol Sensing Molecule 67-KDa Laminin Receptor. FEBS Lett. 2011, 585, 814–820. [Google Scholar] [CrossRef]
- Kumazoe, M.; Yamashita, M.; Nakamura, Y.; Takamatsu, K.; Bae, J.; Yamashita, S.; Yamada, S.; Onda, H.; Nojiri, T.; Kangawa, K.; et al. Green Tea Polyphenol EGCG Upregulates Tollip Expression by Suppressing Elf-1 Expression. J. Immunol. 2017, 199, 3261–3269. [Google Scholar] [CrossRef] [PubMed]
- Ravetch, J.V.; Kinet, J.P. Fc Receptors. Annu. Rev. Immunol. 1991, 9, 457–492. [Google Scholar] [CrossRef] [PubMed]
- Metzger, H. The Receptor with High Affinity for IgE. Immunol. Rev. 1992, 125, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Turner, H.; Kinet, J.P. Signalling through the High-Affinity IgE Receptor FcεRI. Nature 1999, 402, 24–30. [Google Scholar] [CrossRef]
- Rivera, J. Molecular Adapters in FcεRI Signaling and the Allergic Response. Curr. Opin. Immunol. 2002, 14, 688–693. [Google Scholar] [CrossRef]
- Pfeiffer, J.R.; Seagrave, J.C.; Davis, B.H.; Deanin, G.G.; Oliver, J.M. Membrane and Cytoskeletal Changes Associated with IgE-Mediated Serotonin Release from Rat Basophilic Leukemia Cells. J. Cell Biol. 1985, 101, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.H.; Adelstein, R.S.; Beaven, M.A. Secretion from Rat Basophilic RBL-2H3 Cells Is Associated with Diphosphorylation of Myosin Light Chains by Myosin Light Chain Kinase as Well as Phosphorylation by Protein Kinase C. J. Biol. Chem. 1994, 269, 536–541. [Google Scholar] [CrossRef]
- Edgar, A.J.; Bennett, J.P. Circular Ruffle Formation in Rat Basophilic Leukemia Cells in Response to Antigen Stimulation. Eur. J. Cell Biol. 1997, 73, 132–140. [Google Scholar]
- Fujimura, Y.; Umeda, D.; Kiyohara, Y.; Sunada, Y.; Yamada, K.; Tachibana, H. The Involvement of the 67 KDa Laminin Receptor-Mediated Modulation of Cytoskeleton in the Degranulation Inhibition Induced by Epigallocatechin-3-O-Gallate. Biochem. Biophys. Res. Commun. 2006, 348, 524–531. [Google Scholar] [CrossRef]
- Ludowyke, R.I.; Peleg, I.; Beaven, M.A.; Adelstein, R.S. Antigen-Induced Secretion of Histamine and the Phosphorylation of Myosin by Protein Kinase C in Rat Basophilic Leukemia Cells. J. Biol. Chem. 1989, 264, 12492–12501. [Google Scholar] [CrossRef]
- Dombrowicz, D.; Flamand, V.; Brigman, K.K.; Koller, B.H.; Kinet, J.P. Abolition of Anaphylaxis by Targeted Disruption of the High Affinity Immunoglobulin E Receptor α Chain Gene. Cell 1993, 75, 969–976. [Google Scholar] [CrossRef]
- Sano, M.; Suzuki, M.; Miyase, T.; Yoshino, K.; Maeda-Yamamoto, M. Novel Antiallergic Catechin Derivatives Isolated from Oolong Tea. J. Agric. Food Chem. 1999, 47, 1906–1910. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Sunada, Y.; Miyase, T.; Sano, M.; Maeda-Yamamoto, M.; Yamada, K. Identification of a Methylated Tea Catechin as an Inhibitor of Degranulation in Human Basophilic KU812 Cells. Biosci. Biotechnol. Biochem. 2000, 64, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Yoshino, K.; Maeda-Yamamoto, M.; Miyase, T.; Sano, M. Inhibitory Effects of Tea Catechins and O-Methylated Derivatives of (−)-Epigallocatechin-3-O-Gallate on Mouse Type IV Allergy. J. Agric. Food Chem. 2000, 48, 5649–5653. [Google Scholar] [CrossRef] [PubMed]
- Maeda-Yamamoto, M.; Inagaki, N.; Kitaura, J.; Chikumoto, T.; Kawahara, H.; Kawakami, Y.; Sano, M.; Miyase, T.; Tachibana, H.; Nagai, H.; et al. O-Methylated Catechins from Tea Leaves Inhibit Multiple Protein Kinases in Mast Cells. J. Immunol. 2004, 172, 4486–4492. [Google Scholar] [CrossRef] [PubMed]
- Maeda-Yamamoto, M.; Ema, K.; Shibuichi, I. In Vitro and in Vivo Anti-Allergic Effects of “benifuuki” Green Tea Containing O-Methylated Catechin and Ginger Extract Enhancement. Cytotechnology 2007, 55, 135–142. [Google Scholar] [CrossRef]
- Fujimura, Y.; Tachibana, H.; Maeda-Yamamoto, M.; Miyase, T.; Sano, M.; Yamada, K. Antiallergic Tea Catechin, (−)-Epigallocatechin-3-O-(3-O-Methyl)-Gallate, Suppresses FcεRl Expression in Human Basophilic KU812 Cells. J. Agric. Food Chem. 2002, 50, 5729–5734. [Google Scholar] [CrossRef]
- Fujimura, Y.; Umeda, D.; Yano, S.; Maeda-Yamamoto, M.; Yamada, K.; Tachibana, H. The 67 KDa Laminin Receptor as a Primary Determinant of Anti-Allergic Effects of O-Methylated EGCG. Biochem. Biophys. Res. Commun. 2007, 364, 79–85. [Google Scholar] [CrossRef]
- Lambert, J.D.; Yang, C.S. Cancer Chemopreventive Activity and Bioavailability of Tea and Tea Polyphenols. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2003, 523–524, 201–208. [Google Scholar] [CrossRef]
- Yano, S.; Fujimura, Y.; Umeda, D.; Miyase, T.; Yamada, K.; Tachibana, H. Relationship between the Biological Activities of Methylated Derivatives of (−)-Epigallocatechin-3-O-Gallate (EGCG) and Their Cell Surface Binding Activities. J. Agric. Food Chem. 2007, 55, 7144–7148. [Google Scholar] [CrossRef]
- Ambros, V. MicroRNAs: Tiny Regulators with Great Potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef]
- Rathod, S.S.; Rani, S.B.; Khan, M.; Muzumdar, D.; Shiras, A. Tumor Suppressive MiRNA-34a Suppresses Cell Proliferation and Tumor Growth of Glioma Stem Cells by Targeting Akt and Wnt Signaling Pathways. FEBS Open Bio 2014, 4, 485–495. [Google Scholar] [CrossRef]
- Zhao, L.; Gu, H.; Chang, J.; Wu, J.; Wang, D.; Chen, S.; Yang, X.; Qian, B. MicroRNA-383 Regulates the Apoptosis of Tumor Cells through Targeting Gadd45g. PLoS ONE 2014, 9, e110472. [Google Scholar] [CrossRef] [PubMed]
- Rusca, N.; Dehò, L.; Montagner, S.; Zielinski, C.E.; Sica, A.; Sallusto, F.; Monticelli, S. MiR-146a and NF-ΚB1 Regulate Mast Cell Survival and T Lymphocyte Differentiation. Mol. Cell. Biol. 2012, 32, 4432–4444. [Google Scholar] [CrossRef]
- Sun, X.; Icli, B.; Wara, A.K.; Belkin, N.; He, S.; Kobzik, L.; Hunninghake, G.M.; Vera, M.P.; Blackwell, T.S.; Baron, R.M.; et al. MicroRNA-181b Regulates NF-ΚB-Mediated Vascular Inflammation. J. Clin. Investig. 2012, 122, 1973–1990. [Google Scholar] [CrossRef]
- Kornfeld, J.W.; Baitzel, C.; Könner, A.C.; Nicholls, H.T.; Vogt, M.C.; Herrmanns, K.; Scheja, L.; Haumaitre, C.; Wolf, A.M.; Knippschild, U.; et al. Obesity-Induced Overexpression of MiR-802 Impairs Glucose Metabolism through Silencing of Hnf1b. Nature 2013, 494, 111–115. [Google Scholar] [CrossRef]
- Zou, Q.; Li, J.; Hong, Q.; Lin, Z.; Wu, Y.; Shi, H.; Ju, Y. Prediction of MicroRNA-Disease Associations Based on Social Network Analysis Methods. Biom. Res. Int. 2015, 2015, 810514. [Google Scholar] [CrossRef]
- Zou, Q.; Li, J.; Song, L.; Zeng, X.; Wang, G. Similarity Computation Strategies in the MicroRNA-Disease Network: A Survey. Brief. Funct. Genom. 2016, 15, 55–64. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA Expression Profiles Classify Human Cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A MicroRNA Expression Signature of Human Solid Tumors Defines Cancer Gene Targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef]
- Parikh, A.; Lee, C.; Joseph, P.; Marchini, S.; Baccarini, A.; Kolev, V.; Romualdi, C.; Fruscio, R.; Shah, H.; Wang, F.; et al. MicroRNA-181a Has a Critical Role in Ovarian Cancer Progression through the Regulation of the Epithelial-Mesenchymal Transition. Nat. Commun. 2014, 5, 2977. [Google Scholar] [CrossRef]
- Milenkovic, D.; Deval, C.; Gouranton, E.; Landrier, J.F.; Scalbert, A.; Morand, C.; Mazur, A. Modulation of MiRNA Expression by Dietary Polyphenols in ApoE Deficient Mice: A New Mechanism of the Action of Polyphenols. PLoS ONE 2012, 7, e29837. [Google Scholar] [CrossRef]
- Hui, C.; Yujie, F.; Lijia, Y.; Long, Y.; Hongxia, X.; Yong, Z.; Jundong, Z.; Qianyong, Z.; Mantian, M. MicroRNA-34a and MicroRNA-21 Play Roles in the Chemopreventive Effects of 3,6-Dihydroxyflavone on 1-Methyl-1-Nitrosourea-Induced Breast Carcinogenesis. Breast Cancer Res. 2012, 14, R80. [Google Scholar] [CrossRef]
- Ohno, M.; Shibata, C.; Kishikawa, T.; Yoshikawa, T.; Takata, A.; Kojima, K.; Akanuma, M.; Kang, Y.J.; Yoshida, H.; Otsuka, M.; et al. The Flavonoid Apigenin Improves Glucose Tolerance through Inhibition of MicroRNA Maturation in MiRNA103 Transgenic Mice. Sci. Rep. 2013, 3, 2553. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, J.X.; Yang, C.S.; Yang, M.Q.; Deng, Y.; Wang, H. Gene Regulation Mediated by MicroRNAs in Response to Green Tea Polyphenol EGCG in Mouse Lung Cancer. BMC Genom. 2014, 15, S3. [Google Scholar] [CrossRef]
- Yamashita, S.; Lin, I.; Oka, C.; Kumazoe, M.; Komatsu, S.; Murata, M.; Kamachi, S.; Tachibana, H. Soy Isoflavone Metabolite Equol Inhibits Cancer Cell Proliferation in a PAP Associated Domain Containing 5-Dependent and an Estrogen Receptor-Independent Manner. J. Nutr. Biochem. 2022, 100, 108910. [Google Scholar] [CrossRef]
- Murata, M.; Nonaka, H.; Komatsu, S.; Goto, M.; Morozumi, M.; Yamada, S.; Lin, I.C.; Yamashita, S.; Tachibana, H. Delphinidin Prevents Muscle Atrophy and Upregulates MIR-23a Expression. J. Agric. Food Chem. 2017, 65, 45–50. [Google Scholar] [CrossRef]
- Yamada, S.; Tsukamoto, S.; Huang, Y.; Makio, A.; Kumazoe, M.; Yamashita, S.; Tachibana, H. Epigallocatechin-3-O-Gallate up-Regulates MicroRNA-Let-7b Expression by Activating 67-KDa Laminin Receptor Signaling in Melanoma Cells. Sci. Rep. 2016, 6, 19225. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Yamashita, S.; Kim, Y.H.; Kumazoe, M.; Huang, Y.; Yamada, K.; Tachibana, H. Oxygen Partial Pressure Modulates 67-KDa Laminin Receptor Expression, Leading to Altered Activity of the Green Tea Polyphenol, EGCG. FEBS Lett. 2012, 586, 3441–3447. [Google Scholar] [CrossRef]
- BOOTH, A.N.; JONES, F.T.; DeEDS, F. Metabolic Fate of Hesperidin, Eriodictyol, Homoeridictyol, and Diosmin. J. Biol. Chem. 1958, 230, 661–668. [Google Scholar] [CrossRef]
- Punyasiri, P.A.N.; Abeysinghe, I.S.B.; Kumar, V.; Treutter, D.; Duy, D.; Gosch, C.; Martens, S.; Forkmann, G.; Fischer, T.C. Flavonoid Biosynthesis in the Tea Plant Camellia Sinensis: Properties of Enzymes of the Prominent Epicatechin and Catechin Pathways. Arch. Biochem. Biophys. 2004, 431, 22–30. [Google Scholar] [CrossRef]
- Kumazoe, M.; Fujimura, Y.; Yoshitomi, R.; Shimada, Y.; Tachibana, H. Fustin, a Flavanonol, Synergically Potentiates the Anticancer Effect of Green Tea Catechin Epigallocatechin-3-O-Gallate with Activation of the ENOS/CGMP Axis. J. Agric. Food Chem. 2022, 70, 3458–3466. [Google Scholar] [CrossRef]
- Byung, C.P.; Yong, S.L.; Park, H.J.; Kwak, M.K.; Bong, K.Y.; Joo, Y.K.; Kim, J.A. Protective Effects of Fustin, a Flavonoid from Rhus Verniciflua Stokes, on 6-Hydroxydopamine-Induced Neuronal Cell Death. Exp. Mol. Med. 2007, 39, 316–326. [Google Scholar] [CrossRef]
- Kamei, R.; Fujimura, T.; Matsuda, M.; Kakihara, K.; Hirakawa, N.; Baba, K.; Ono, K.; Arakawa, K.; Kawamoto, S. A Flavanone Derivative from the Asian Medicinal Herb (Perilla frutescens) Potently Suppresses IgE-Mediated Immediate Hypersensitivity Reactions. Biochem. Biophys. Res. Commun. 2017, 483, 674–679. [Google Scholar] [CrossRef]
- Bai, Y.; Peng, W.; Yang, C.; Zou, W.; Liu, M.; Wu, H.; Fan, L.; Li, P.; Zeng, X.; Su, W. Pharmacokinetics and Metabolism of Naringin and Active Metabolite Naringenin in Rats, Dogs, Humans, and the Differences Between Species. Front. Pharmacol. 2020, 11, 364. [Google Scholar] [CrossRef]
- Kumazoe, M.; Takai, M.; Hiroi, S.; Takeuchi, C.; Yamanouchi, M.; Nojiri, T.; Onda, H.; Bae, J.; Huang, Y.; Takamatsu, K.; et al. PDE3 Inhibitor and EGCG Combination Treatment Suppress Cancer Stem Cell Properties in Pancreatic Ductal Adenocarcinoma. Sci. Rep. 2017, 7, 1917. [Google Scholar] [CrossRef]
- Bae, J.; Kumazoe, M.; Yamashita, S.; Tachibana, H. Hydrogen Sulphide Donors Selectively Potentiate a Green Tea Polyphenol EGCG-Induced Apoptosis of Multiple Myeloma Cells. Sci. Rep. 2017, 7, 6665. [Google Scholar] [CrossRef]
- Bae, J.; Kumazoe, M.; Fujimura, Y.; Tachibana, H. Diallyl Disulfide Potentiates Anti-Obesity Effect of Green Tea in High-Fat/High-Sucrose Diet-Induced Obesity. J. Nutr. Biochem. 2019, 64, 152–161. [Google Scholar] [CrossRef]
- Yamashita, S.; Hirashima, A.; Lin, I.C.; Bae, J.; Nakahara, K.; Murata, M.; Yamada, S.; Kumazoe, M.; Yoshitomi, R.; Kadomatsu, M.; et al. Saturated Fatty Acid Attenuates Anti-Obesity Effect of Green Tea. Sci. Rep. 2018, 8, 10023. [Google Scholar] [CrossRef]
- Shukla, R.; Chanda, N.; Zambre, A.; Upendran, A.; Katti, K.; Kulkarni, R.R.; Nune, S.K.; Casteel, S.W.; Smith, C.J.; Vimal, J.; et al. Laminin Receptor Specific Therapeutic Gold Nanoparticles (198AuNP-EGCg) Show Efficacy in Treating Prostate Cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 12426–12431. [Google Scholar] [CrossRef]
- Santilli, G.; Piotrowska, I.; Cantilena, S.; Chayka, O.; D’Alicarnasso, M.; Morgenstern, D.A.; Himoudi, N.; Pearson, K.; Anderson, J.; Thrasher, A.J.; et al. Polyphenol e Enhances the Antitumor Immune Response in Neuroblastoma by Inactivating Myeloid Suppressor Cells. Clin. Cancer Res. 2013, 19, 1116–1125. [Google Scholar] [CrossRef]
- Mocanu, M.M.; Ganea, C.; Georgescu, L.; Váradi, T.; Shrestha, D.; Baran, I.; Katona, E.; Nagy, P.; Szöllosi, J. Epigallocatechin 3-O-Gallate Induces 67 KDa Laminin Receptor-Mediated Cell Death Accompanied by Downregulation of ErbB Proteins and Altered Lipid Raft Clustering in Mammary and Epidermoid Carcinoma Cells. J. Nat. Prod. 2014, 77, 250–257. [Google Scholar] [CrossRef]
- Holy, E.W.; Stämpfli, S.F.; Akhmedov, A.; Holm, N.; Camici, G.G.; Lüscher, T.F.; Tanner, F.C. Laminin Receptor Activation Inhibits Endothelial Tissue Factor Expression. J. Mol. Cell. Cardiol. 2010, 48, 1138–1145. [Google Scholar] [CrossRef]
- Wang, Q.M.; Wang, H.; Li, Y.F.; Xie, Z.Y.; Ma, Y.; Yan, J.J.; Gao, Y.F.W.; Wang, Z.M.; Wang, L.S. Inhibition of EMMPRIN and MMP-9 Expression by Epigallocatechin-3-Gallate through 67-KDa Laminin Receptor in PMA-Induced Macrophages. Cell. Physiol. Biochem. 2016, 39, 2308–2319. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Li, Y.; Yu, H.; Wang, W.; Wu, C.; Yang, Y.; Hu, Y.; Shi, X.; Li, J. Epigallocatechin-3-Gallate Inhibits H2O2-Induced Apoptosis in Mouse Vascular Smooth Muscle Cells via 67kD Laminin Receptor. Sci. Rep. 2017, 7, 7774. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.F.; Tsuei, Y.W.; Liu, C.W.; Kao, C.C.; Shih, L.J.; Ho, L.T.; Wu, L.Y.; Wu, C.P.; Tsai, P.H.; Chang, H.H.; et al. Green Tea Epigallocatechin Gallate Inhibits Insulin Stimulation of Adipocyte Glucose Uptake via the 67-Kilodalton Laminin Receptor and AMP-Activated Protein Kinase Pathways. Planta Med. 2010, 76, 1694–1698. [Google Scholar] [CrossRef] [PubMed]
- Ku, H.C.; Chang, H.H.; Liu, H.C.; Hsiao, C.H.; Lee, M.J.; Hu, Y.J.; Hung, P.F.; Liu, C.W.; Kao, Y.H. Green Tea (−)-Epigallocatechin Gallate Inhibits Insulin Stimulation of 3T3-L1 Preadipocyte Mitogenesis via the 67-KDa Laminin Receptor Pathway. Am. J. Physiol.—Cell Physiol. 2009, 297, C121–C132. [Google Scholar] [CrossRef] [PubMed]
- Ku, H.C.; Liu, H.S.; Hung, P.F.; Chen, C.L.; Liu, H.C.; Chang, H.H.; Tsuei, Y.W.; Shih, L.J.; Lin, C.L.; Lin, C.M.; et al. Green Tea (−)-Epigallocatechin Gallate Inhibits IGF-I and IGF-IIstimulation of 3T3-L1 Preadipocyte Mitogenesis via the 67-KDa Laminin Receptor, but Not AMP-Activated Protein Kinase Pathway. Mol. Nutr. Food Res. 2012, 56, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Ku, H.C.; Tsuei, Y.W.; Kao, C.C.; Weng, J.T.; Shih, L.J.; Chang, H.H.; Liu, C.W.; Tsai, S.W.; Kuo, Y.C.; Kao, Y.H. Green Tea (−)-Epigallocatechin Gallate Suppresses IGF-I and IGF-II Stimulation of 3T3-L1 Adipocyte Glucose Uptake via the Glucose Transporter 4, but Not Glucose Transporter 1 Pathway. Gen. Comp. Endocrinol. 2014, 199, 46–55. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Chen, Y.; Wu, J.; Li, M.; Yang, H.; Xu, W.; Wei, L. EGCG Induces Pro-Inflammatory Response in Macrophages to Prevent Bacterial Infection through the 67LR/P38/JNK Signaling Pathway. J. Agric. Food Chem. 2021, 69, 5638–5651. [Google Scholar] [CrossRef] [PubMed]
- Byun, E.B.; Choi, H.G.; Sung, N.Y.; Byun, E.H. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits TLR4 Signaling through the 67-KDa Laminin Receptor on Lipopolysaccharide-Stimulated Dendritic Cells. Biochem. Biophys. Res. Commun. 2012, 426, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ye, L.; Wang, X.; Liu, J.; Wang, Y.; Zhou, Y.; Ho, W. (−)-Epigallocatechin Gallate Inhibits Endotoxin-Induced Expression of Inflammatory Cytokines in Human Cerebral Microvascular Endothelial Cells. J. Neuroinflamm. 2012, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Byun, E.B.; Kim, W.S.; Sung, N.Y.; Byun, E.H. Epigallocatechin-3-Gallate Regulates Anti-Inflammatory Action Through 67-KDa Laminin Receptor-Mediated Tollip Signaling Induction in Lipopolysaccharide-Stimulated Human Intestinal Epithelial Cells. Cell. Physiol. Biochem. 2018, 46, 2072–2081. [Google Scholar] [CrossRef]
- Luo, D.; Xu, J.; Chen, X.; Zhu, X.; Liu, S.; Li, J.; Xu, X.; Ma, X.; Zhao, J.; Ji, X. (−)-Epigallocatechin-3-Gallate (EGCG) Attenuates Salt-Induced Hypertension and Renal Injury in Dahl Salt-Sensitive Rats. Sci. Rep. 2020, 10, 4783. [Google Scholar] [CrossRef]
- Kim, J.E.; Park, H.; Lee, J.E.; Kang, T.C. Blockade of 67-KDa Laminin Receptor Facilitates AQP4 Down-Regulation and BBB Disruption via ERK1/2-and P38 MAPK-Mediated PI3K/AKT Activations. Cells 2020, 9, 1670. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Park, H.; Jeong, M.J.; Kang, T.C. Epigallocatechin-3-gallate and PEDF 335 Peptide, 67LR Activators, Attenuate Vasogenic Edema, and Astroglial Degeneration Following Status Epilepticus. Antioxidants 2020, 9, 854. [Google Scholar] [CrossRef] [PubMed]
- Dorchies, O.M.; Wagner, S.; Buetler, T.M.; Ruegg, U.T. Protection of Dystrophic Muscle Cells with Polyphenols from Green Tea Correlates with Improved Glutathione Balance and Increased Expression of 67LR, a Receptor for (2)-Epigallocatechin Gallate. BioFactors 2009, 35, 279–294. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Guo, Y.; Li, Y.; Jia, Q.; Han, X.; Liu, B.; Chen, J.; Cheng, J.C.; Sun, Y.P. Epigallocatechin-3-Gallate Stimulates StAR Expression and Progesterone Production in Human Granulosa Cells through the 67-KDa Laminin Receptor-Mediated CREB Signaling Pathway. J. Cell. Physiol. 2022, 237, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Bátori, R.; Bécsi, B.; Nagy, D.; Kónya, Z.; Hegedũs, C.; Bordán, Z.; Verin, A.; Lontay, B.; Erdõdi, F. Interplay of Myosin Phosphatase and Protein Phosphatase-2A in the Regulation of Endothelial Nitric-Oxide Synthase Phosphorylation and Nitric Oxide Production. Sci. Rep. 2017, 7, 44698. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.R.; Hsu, C.S.; Lu, C.H.; Chen, W.C.; Chiu, C.H.; Liou, Y.M. Epigallocatechin-3-Gallate-Mediated Cardioprotection by Akt/GSK-3β/Caveolin Signalling in H9c2 Rat Cardiomyoblasts. J. Biomed. Sci. 2013, 20, 86. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.M.; Gao, W.; Wang, H.; Zhao, D.; Nie, Z.L.; Shi, J.Q.; Zhao, S.; Lu, X.; Wang, L.S.; Yang, Z.J. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits TNF-Induced Production of Monocyte Chemoattractant Protein-1 in Human Umbilical Vein Endothelial Cells. Cell. Physiol. Biochem. 2014, 33, 1349–1358. [Google Scholar] [CrossRef]
- Gundimeda, U.; McNeill, T.H.; Elhiani, A.A.; Schiffman, J.E.; Hinton, D.R.; Gopalakrishna, R. Green Tea Polyphenols Precondition against Cell Death Induced by Oxygen-Glucose Deprivation via Stimulation of Laminin Receptor, Generation of Reactive Oxygen Species, and Activation of Protein Kinase Cε. J. Biol. Chem. 2012, 287, 34694–34708. [Google Scholar] [CrossRef]
- Gundimeda, U.; McNeill, T.H.; Fan, T.K.; Deng, R.; Rayudu, D.; Chen, Z.; Cadenas, E.; Gopalakrishna, R. Green Tea Catechins Potentiate the Neuritogenic Action of Brain-Derived Neurotrophic Factor: Role of 67-KDa Laminin Receptor and Hydrogen Peroxide. Biochem. Biophys. Res. Commun. 2014, 445, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Gundimeda, U.; McNeill, T.H.; Barseghian, B.A.; Tzeng, W.S.; Rayudu, D.V.; Cadenas, E.; Gopalakrishna, R. Polyphenols from Green Tea Prevent Antineuritogenic Action of Nogo-A via 67-KDa Laminin Receptor and Hydrogen Peroxide. J. Neurochem. 2015, 132, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Won, Y.S.; Yang, X.; Kumazoe, M.; Yamashita, S.; Hara, A.; Takagaki, A.; Goto, K.; Nanjo, F.; Tachibana, H. Green Tea Catechin Metabolites Exert Immunoregulatory Effects on CD4+ T Cell and Natural Killer Cell Activities. J. Agric. Food Chem. 2016, 64, 3591–3597. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujimura, Y.; Kumazoe, M.; Tachibana, H. 67-kDa Laminin Receptor-Mediated Cellular Sensing System of Green Tea Polyphenol EGCG and Functional Food Pairing. Molecules 2022, 27, 5130. https://doi.org/10.3390/molecules27165130
Fujimura Y, Kumazoe M, Tachibana H. 67-kDa Laminin Receptor-Mediated Cellular Sensing System of Green Tea Polyphenol EGCG and Functional Food Pairing. Molecules. 2022; 27(16):5130. https://doi.org/10.3390/molecules27165130
Chicago/Turabian StyleFujimura, Yoshinori, Motofumi Kumazoe, and Hirofumi Tachibana. 2022. "67-kDa Laminin Receptor-Mediated Cellular Sensing System of Green Tea Polyphenol EGCG and Functional Food Pairing" Molecules 27, no. 16: 5130. https://doi.org/10.3390/molecules27165130
APA StyleFujimura, Y., Kumazoe, M., & Tachibana, H. (2022). 67-kDa Laminin Receptor-Mediated Cellular Sensing System of Green Tea Polyphenol EGCG and Functional Food Pairing. Molecules, 27(16), 5130. https://doi.org/10.3390/molecules27165130