
Current Use of Fenton Reaction in Drugs and Food



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Fenton Reaction
3. Fenton Reaction in Body
3.1. Iron as a Nutrient
3.2. Fenton Reaction under Biological Environment
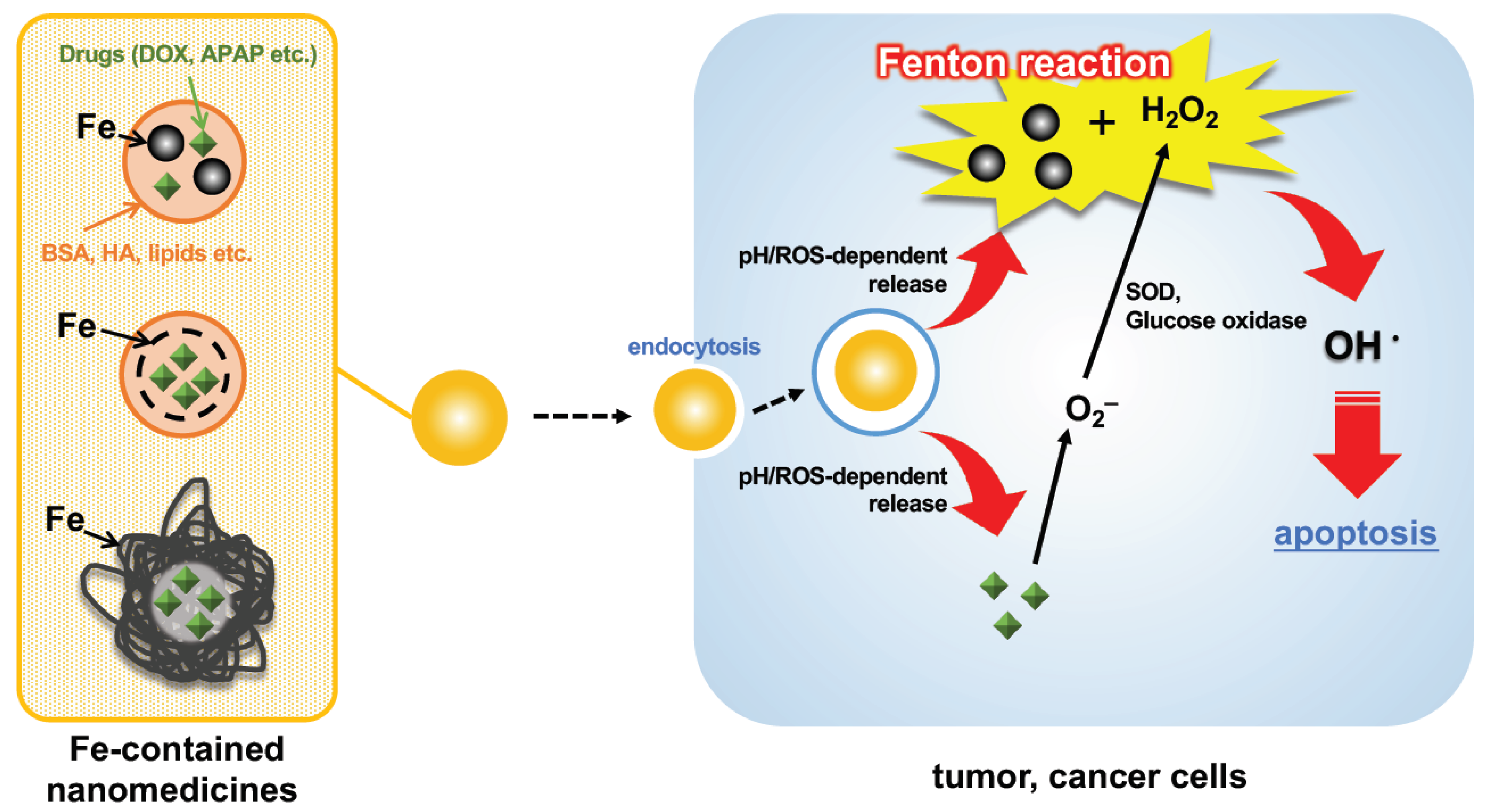
4. Use of the Fenton Reaction for Drugs
5. Fenton Reaction in Food
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ozawa, H.; Miyazawa, T.; Burdeos, G.C.; Miyazawa, T. Biological Functions of Antioxidant Dipeptides. J. Nutr. Sci. Vitaminol. 2022, 68, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, S.; Kakinuma, G.; Kawashima, K.; Tanikawa, K.; Ochiai, S.; Miyata, R.; Fujisaki, K.; Kanao, R.; Noguchi, K.; Aoki, K.; et al. The Etiology of Beriberi. J. Am. Med. Assoc. 1929, 92, 1325–1327. [Google Scholar] [CrossRef]
- Miyazawa, T.; Burdeos, G.C.; Itaya, M.; Nakagawa, K.; Miyazawa, T. Vitamin E: Regulatory Redox Interactions. IUBMB Life 2019, 71, 430–441. [Google Scholar] [CrossRef]
- Poskitt, E.M.E. Early History of Iron Deficiency. Br. J. Haematol. 2003, 122, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Biswas, J.P.; Paul, S.; Paik, A.; Maiti, D. Organic Synthesis with the Most Abundant Transition Metal–Iron: From Rust to Multitasking Catalysts. Chem. Soc. Rev. 2021, 50, 243–472. [Google Scholar] [CrossRef]
- Andrews, N.C. Iron Metabolism: Iron Deficiency and Iron Overload. Annu. Rev. Genomics Hum. Genet. 2000, 1, 75–98. [Google Scholar] [CrossRef]
- Diaz-Ochoa, V.E.; Jellbauer, S.; Klaus, S.; Raffatellu, M. Transition Metal Ions at the Crossroads of Mucosal Immunity and Microbial Pathogenesis. Front. Cell. Infect. Microbiol. 2014, 4, 2. [Google Scholar] [CrossRef]
- Sheftel, A.D.; Mason, A.B.; Ponka, P. The Long History of Iron in the Universe and in Health and Disease. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 161–187. [Google Scholar] [CrossRef]
- Kell, D.B.; Heyden, E.L.; Pretorius, E. The Biology of Lactoferrin, an Iron-Binding Protein That Can Help Defend Against Viruses and Bacteria. Front. Immunol. 2020, 11, 1221. [Google Scholar] [CrossRef]
- Jensen, F.B. The Dual Roles of Red Blood Cells in Tissue Oxygen Delivery: Oxygen Carriers and Regulators of Local Blood Flow. J. Exp. Biol. 2009, 212, 3387–3393. [Google Scholar] [CrossRef] [Green Version]
- Puig, S.; Ramos-Alonso, L.; Romero, A.M.; Martínez-Pastor, M.T. The Elemental Role of Iron in DNA Synthesis and Repair. Metallomics 2017, 9, 1483–1500. [Google Scholar] [CrossRef] [PubMed]
- Fenton, H.J.H. LXXIII.—Oxidation of Tartaric Acid in Presence of Iron. J. Chem. Soc. Trans. 1894, 65, 899–910. [Google Scholar] [CrossRef]
- Ruan, L.; Wang, M.; Zhou, M.; Lu, H.; Zhang, J.; Gao, J.; Chen, J.; Hu, Y. Doxorubicin–Metal Coordinated Micellar Nanoparticles for Intracellular Codelivery and Chemo/Chemodynamic Therapy In Vitro. ACS Appl. Bio Mater. 2019, 2, 4703–4707. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, M.; Cheng, J.; Yin, J.; Huang, C.; Cui, H.; Zhang, X.; Zhao, G. Acidity-Triggered Tumor-Targeted Nanosystem for Synergistic Therapy via a Cascade of ROS Generation and NO Release. ACS Appl. Mater. Interfaces 2020, 12, 28975–28984. [Google Scholar] [CrossRef]
- Yang, N.; Zhang, T.; Cao, C.; Mao, G.; Shao, J.; Song, X.; Wang, W.; Mou, X.; Dong, X. BSA Stabilized Photothermal-Fenton Reactor with Cisplatin for Chemo/Chemodynamic Cascade Oncotherapy. Nano Res. 2022, 15, 2235–2243. [Google Scholar] [CrossRef]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current Evidence on the Effect of Dietary Polyphenols Intake on Chronic Diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef]
- Duffuler, P.; Bhullar, K.S.; de Campos Zani, S.C.; Wu, J. Bioactive Peptides: From Basic Research to Clinical Trials and Commercialization. J. Agric. Food Chem. 2022, 70, 3585–3595. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.H.; Parihar, A.; Doseff, A.I. Dietary Flavonoids for Immunoregulation and Cancer: Food Design for Targeting Disease. Antioxidants 2019, 8, 202. [Google Scholar] [CrossRef]
- Schoenfeld, J.D.; Sibenaller, Z.A.; Mapuskar, K.A.; Wagner, B.A.; Cramer-Morales, K.L.; Furqan, M.; Sandhu, S.; Carlisle, T.L.; Smith, M.C.; Abu Hejleh, T.; et al. O2− and H2O2-Mediated Disruption of Fe Metabolism Causes the Differential Susceptibility of NSCLC and GBM Cancer Cells to Pharmacological Ascorbate. Cancer Cell 2017, 31, 487–500. [Google Scholar] [CrossRef]
- Puppo, A. Effect of Flavonoids on Hydroxyl Radical Formation by Fenton-Type Reactions; Influence of the Iron Chelator. Phytochemistry 1992, 31, 85–88. [Google Scholar] [CrossRef]
- Xu, G.; Wang, H.; Li, X.; Huang, R.; Luo, L. Recent Progress on Targeting Ferroptosis for Cancer Therapy. Biochem. Pharmacol. 2021, 190, 114584. [Google Scholar] [CrossRef] [PubMed]
- Haber, F.; Willstätter, R. Unpaarigkeit Und Radikalketten Im Reaktionsmechanismus Organischer Und Enzymatischer Vorgänge. Ber. Dtsch. Chem. Ges. 1931, 64, 2844–2856. [Google Scholar] [CrossRef]
- Haber, F.; Weiss, J.; Pope, W.J. The Catalytic Decomposition of Hydrogen Peroxide by Iron Salts. Proc. R. Soc. London. Ser. A Math. Phys. Sci. 1934, 147, 332–351. [Google Scholar] [CrossRef]
- Stanbury, D.M. The Principle of Detailed Balancing, the Iron-Catalyzed Disproportionation of Hydrogen Peroxide, and the Fenton Reaction. Dalton Trans. 2022, 51, 2135–2157. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-F.; Chen, H.-F.; Kao, C.-L.; Chao, I.; Chen, H.-Y. A Computational Study of the Fenton Reaction in Different PH Ranges. Phys. Chem. Chem. Phys. 2018, 20, 22890–22901. [Google Scholar] [CrossRef] [PubMed]
- Kremer, M.L. The Fenton Reaction. Dependence of the Rate on PH. J. Phys. Chem. A 2003, 107, 1734–1741. [Google Scholar] [CrossRef]
- Chen, H.-Y. Why the Reactive Oxygen Species of the Fenton Reaction Switches from Oxoiron(IV) Species to Hydroxyl Radical in Phosphate Buffer Solutions? A Computational Rationale. ACS Omega 2019, 4, 14105–14113. [Google Scholar] [CrossRef]
- Shinichi, E.; Yosuke, S.; Colussi, A.J. Fenton Chemistry at Aqueous Interfaces. Proc. Natl. Acad. Sci. USA 2014, 111, 623–628. [Google Scholar] [CrossRef]
- Deguillaume, L.; Leriche, M.; Chaumerliac, N. Impact of Radical versus Non-Radical Pathway in the Fenton Chemistry on the Iron Redox Cycle in Clouds. Chemosphere 2005, 60, 718–724. [Google Scholar] [CrossRef]
- Prasad, R.; Majumdar, K.; Shivay, Y.; Kapil, U. Minerals in Plant and Human Nutrition and Health; International Plant Nutrition Institute (IPNI) Peachtree Corners: Peachtree Corners, GA, USA, 2016. [Google Scholar]
- Weed, R.I.; Reed, C.F.; Berg, G. Is Hemoglobin an Essential Structural Component of Human Erythrocyte Membranes? J. Clin. Investig. 1963, 42, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Lumb, A.B. Physiology of Haemoglobin. Contin. Educ. Anaesth. Crit. Care Pain 2012, 12, 251–256. [Google Scholar] [CrossRef]
- Pasricha, S.-R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron Deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef]
- Laftah, A.H.; Latunde-Dada, G.O.; Fakih, S.; Hider, R.C.; Simpson, R.J.; McKie, A.T. Haem and Folate Transport by Proton-Coupled Folate Transporter/Haem Carrier Protein 1 (SLC46A1). Br. J. Nutr. 2008, 101, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and Characterization of a Mammalian Proton-Coupled Metal-Ion Transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The Iron Exporter Ferroportin/Slc40a1 Is Essential for Iron Homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef]
- Billesbølle, C.B.; Azumaya, C.M.; Kretsch, R.C.; Powers, A.S.; Gonen, S.; Schneider, S.; Arvedson, T.; Dror, R.O.; Cheng, Y.; Manglik, A. Structure of Hepcidin-Bound Ferroportin Reveals Iron Homeostatic Mechanisms. Nature 2020, 586, 807–811. [Google Scholar] [CrossRef]
- Camaschella, C. Iron Deficiency. Blood 2019, 133, 30–39. [Google Scholar] [CrossRef]
- Yiannikourides, A.; Latunde-Dada, G.O. A Short Review of Iron Metabolism and Pathophysiology of Iron Disorders. Medicines 2019, 6, 85. [Google Scholar] [CrossRef]
- Lesjak, M.; Srai, S.K.S. Role of Dietary Flavonoids in Iron Homeostasis. Pharmaceuticals 2019, 12, 119. [Google Scholar] [CrossRef]
- Lesjak, M.; Hoque, R.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.S.; Sharp, P.A. Quercetin Inhibits Intestinal Iron Absorption and Ferroportin Transporter Expression In Vivo and In Vitro. PLoS ONE 2014, 9, e102900. [Google Scholar] [CrossRef] [Green Version]
- Lazrak, M.; El Kari, K.; Stoffel, N.U.; Elammari, L.; Al-Jawaldeh, A.; Loechl, C.U.; Yahyane, A.; Barkat, A.; Zimmermann, M.B.; Aguenaou, H. Tea Consumption Reduces Iron Bioavailability from NaFeEDTA in Nonanemic Women and Women with Iron Deficiency Anemia: Stable Iron Isotope Studies in Morocco. J. Nutr. 2021, 151, 2714–2720. [Google Scholar] [CrossRef] [PubMed]
- Shubham, K.; Anukiruthika, T.; Dutta, S.; Kashyap, A.V.; Moses, J.A.; Anandharamakrishnan, C. Iron Deficiency Anemia: A Comprehensive Review on Iron Absorption, Bioavailability and Emerging Food Fortification Approaches. Trends Food Sci. Technol. 2020, 99, 58–75. [Google Scholar] [CrossRef]
- Sonnweber, T.; Ress, C.; Nairz, M.; Theurl, I.; Schroll, A.; Murphy, A.T.; Wroblewski, V.; Witcher, D.R.; Moser, P.; Ebenbichler, C.F.; et al. High-Fat Diet Causes Iron Deficiency via Hepcidin-Independent Reduction of Duodenal Iron Absorption. J. Nutr. Biochem. 2012, 23, 1600–1608. [Google Scholar] [CrossRef]
- Cepeda-Lopez, A.C.; Melse-Boonstra, A.; Zimmermann, M.B.; Herter-Aeberli, I. In Overweight and Obese Women, Dietary Iron Absorption Is Reduced and the Enhancement of Iron Absorption by Ascorbic Acid Is One-Half That in Normal-Weight Women. Am. J. Clin. Nutr. 2015, 102, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Villaño, D.; Vilaplana, C.; Medina, S.; Algaba-Chueca, F.; Cejuela-Anta, R.; Martínez-Sanz, J.M.; Ferreres, F.; Gil-Izquierdo, A. Relationship between the Ingestion of a Polyphenol-Rich Drink, Hepcidin Hormone, and Long-Term Training. Molecules 2016, 21, 1333. [Google Scholar] [CrossRef]
- Mu, M.; An, P.; Wu, Q.; Shen, X.; Shao, D.; Wang, H.; Zhang, Y.; Zhang, S.; Yao, H.; Min, J.; et al. The Dietary Flavonoid Myricetin Regulates Iron Homeostasis by Suppressing Hepcidin Expression. J. Nutr. Biochem. 2016, 30, 53–61. [Google Scholar] [CrossRef]
- Zaritsky, J.; Young, B.; Wang, H.-J.; Westerman, M.; Olbina, G.; Nemeth, E.; Ganz, T.; Rivera, S.; Nissenson, A.R.; Salusky, I.B. Hepcidin—A Potential Novel Biomarker for Iron Status in Chronic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Jian, J.; Katz, S.; Abramson, S.B.; Huang, X. 17β-Estradiol Inhibits Iron Hormone Hepcidin Through an Estrogen Responsive Element Half-Site. Endocrinology 2012, 153, 3170–3178. [Google Scholar] [CrossRef]
- Dasa, F.; Abera, T. Factors Affecting Iron Absorption and Mitigation Mechanisms: A Review. Int. J. Agric. Sci. Food Technol. 2018, 4, 24–30. [Google Scholar] [CrossRef]
- Diebold, L.; Chandel, N.S. Mitochondrial ROS Regulation of Proliferating Cells. Free Radic. Biol. Med. 2016, 100, 86–93. [Google Scholar] [CrossRef]
- Locato, V.; Cimini, S.; De Gara, L. ROS and Redox Balance as Multifaceted Players of Cross-Tolerance: Epigenetic and Retrograde Control of Gene Expression. J. Exp. Bot. 2018, 69, 3373–3391. [Google Scholar] [CrossRef] [PubMed]
- Qing, L.; Utta, B.-P.; Ulrike, M.; Martina, B.; Christoph, W.; Helmut, A.; Kurt, J.; Thomas, K. A Fenton Reaction at the Endoplasmic Reticulum Is Involved in the Redox Control of Hypoxia-Inducible Gene Expression. Proc. Natl. Acad. Sci. USA 2004, 101, 4302–4307. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, H.; Chai, Z.; Shi, W.; Li, X.; Ma, H. An Endoplasmic Reticulum-Targeting Fluorescent Probe for Imaging ˙OH in Living Cells. Chem. Commun. 2020, 56, 6344–6347. [Google Scholar] [CrossRef]
- Anelli, T.; Sitia, R. Protein Quality Control in the Early Secretory Pathway. EMBO J. 2008, 27, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Sung, G.; Kim, G.; Park, J.; Jin, B.; Kim, S.K. Effect of Various Intercalators on the Fenton-Type Oxidative Cleavage of Double-Stranded DNA. Chem. Asian J. 2014, 9, 1341–1348. [Google Scholar] [CrossRef]
- Imlay, J.A.; Chin, S.M.; Linn, S. Toxic DNA Damage by Hydrogen Peroxide Through the Fenton Reaction in Vivo and in Vitro. Science 1988, 240, 640–642. [Google Scholar] [CrossRef]
- Gutteridge, J.M.C. Iron Promoters of the Fenton Reaction and Lipid Peroxidation Can Be Released from Haemoglobin by Peroxides. FEBS Lett. 1986, 201, 291–295. [Google Scholar] [CrossRef]
- Miyazawa, T. Lipid Hydroperoxides in Nutrition, Health, and Diseases. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2021, 97, 161–196. [Google Scholar] [CrossRef]
- Thomas, D.D.; Michael, E.G.; Vitek, M.P.; Miranda, K.M.; Wink, D.A. Protein Nitration Is Mediated by Heme and Free Metals through Fenton-Type Chemistry: An Alternative to the NO/O2− Reaction. Proc. Natl. Acad. Sci. USA 2002, 99, 12691–12696. [Google Scholar] [CrossRef]
- Murakami, K.; Shimizu, T.; Irie, K. Formation of the 42-Mer and Amyloid β Radical and the Therapeutic Role of Superoxide Dismutase in Alzheimer’s Disease. J. Amino Acids 2011, 2011, 654207. [Google Scholar] [CrossRef] [Green Version]
- Hachimoto, S.; Tatsuno, Y.; Kitagawa, T. Resonance Raman Evidence for the Presence of the FeIV=O Bond in Horseradish Peroxidase Compound II. Proc. Jpn. Acad. Ser. B 1984, 60, 345–348. [Google Scholar] [CrossRef]
- Carlomagno, F. Ferritinophagy and Cell Cycle Control. Blood 2016, 128, SCI-20. [Google Scholar] [CrossRef]
- Ma, S.; Fu, X.; Liu, L.; Liu, Y.; Feng, H.; Jiang, H.; Liu, X.; Liu, R.; Liang, Z.; Li, M.; et al. Iron-Dependent Autophagic Cell Death Induced by Radiation in MDA-MB-231 Breast Cancer Cells. Front. Cell Dev. Biol. 2021, 9, 723801. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Stockwell, B.R. Synthetic Lethal Screening Identifies Compounds Activating Iron-Dependent, Nonapoptotic Cell Death in Oncogenic-RAS-Harboring Cancer Cells. Chem. Biol. 2008, 15, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Chung, S.W. ROS-Mediated Autophagy Increases Intracellular Iron Levels and Ferroptosis by Ferritin and Transferrin Receptor Regulation. Cell Death Dis. 2019, 10, 822. [Google Scholar] [CrossRef]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and Function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef]
- Chen, X.; Li, J.; Kang, R.; Klionsky, D.J.; Tang, D. Ferroptosis: Machinery and Regulation. Autophagy 2021, 17, 2054–2081. [Google Scholar] [CrossRef]
- Bebber, C.M.; Müller, F.; Prieto Clemente, L.; Weber, J.; von Karstedt, S. Ferroptosis in Cancer Cell Biology. Cancers 2020, 12, 164. [Google Scholar] [CrossRef]
- Young, I.S.; Woodside, J.V. Antioxidants in Health and Disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Zhang, X.; Liu, M.; Cai, B.; He, N.; Wang, Z. Fenton Reaction-Based Nanomedicine in Cancer Chemodynamic and Synergistic Therapy. Appl. Mater. Today 2020, 21, 100864. [Google Scholar] [CrossRef]
- Ranji-Burachaloo, H.; Gurr, P.A.; Dunstan, D.E.; Qiao, G.G. Cancer Treatment through Nanoparticle-Facilitated Fenton Reaction. ACS Nano 2018, 12, 11819–11837. [Google Scholar] [CrossRef]
- Miyazawa, T.; Itaya, M.; Burdeos, G.C.; Nakagawa, K.; Miyazawa, T. A Critical Review of the Use of Surfactant-Coated Nanoparticles in Nanomedicine and Food Nanotechnology. Int. J. Nanomed. 2021, 16, 3937–3999. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Liu, X.-Y.; Zhou, T.-J.; Wan, X.; Wang, Y.; Jiang, H.-L. Photothermal Nanozyme-Ignited Fenton Reaction-Independent Ferroptosis for Breast Cancer Therapy. J. Control. Release 2021, 339, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; An, L.; Tian, Q.; Lin, J.; Yang, S. Ellagic Acid-Fe@BSA Nanoparticles for Endogenous H2S Accelerated Fe(III)/Fe(II) Conversion and Photothermal Synergistically Enhanced Chemodynamic Therapy. Theranostics 2020, 10, 4101–4115. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Cao, F.; Li, W.; Zhang, L.; You, Y.; Deng, Q.; Dong, K.; Ren, J.; Qu, X. Bioinspired Construction of a Nanozyme-Based H2O2 Homeostasis Disruptor for Intensive Chemodynamic Therapy. J. Am. Chem. Soc. 2020, 142, 5177–5183. [Google Scholar] [CrossRef]
- Gao, F.; Wang, F.; Nie, X.; Zhang, Z.; Chen, G.; Xia, L.; Wang, L.-H.; Wang, C.-H.; Hao, Z.-Y.; Zhang, W.-J.; et al. Mitochondria-Targeted Delivery and Light Controlled Release of Iron Prodrug and CO to Enhance Cancer Therapy by Ferroptosis. New J. Chem. 2020, 44, 3478–3486. [Google Scholar] [CrossRef]
- You, C.; Wu, H.; Gao, Z.; Chen, F.; Ning, L.; Zhang, Y.; Dong, Y.; Sun, B.; Wang, F. Enhanced Reactive Oxygen Species Levels by an Active Benzothiazole Complex-Mediated Fenton Reaction for Highly Effective Antitumor Therapy. Mol. Pharm. 2019, 16, 4929–4939. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, Q.; Zhang, M.; Sun, B.; She, Z.; Ge, M.; Lu, T.; Chu, X.; Wang, Y.; Wang, J.; et al. Hollow Porous Carbon Coated FeS2-Based Nanocatalysts for Multimodal Imaging-Guided Photothermal, Starvation, and Triple-Enhanced Chemodynamic Therapy of Cancer. ACS Appl. Mater. Interfaces 2020, 12, 10142–10155. [Google Scholar] [CrossRef]
- Fu, X.; Zhang, Y.; Zhang, G.; Li, X.; Ni, S.; Cui, J. Targeted Delivery of Fenton Reaction Packages and Drugs for Cancer Theranostics. Appl. Mater. Today 2022, 26, 101353. [Google Scholar] [CrossRef]
- Chen, Q.; Zhou, J.; Chen, Z.; Luo, Q.; Xu, J.; Song, G. Tumor-Specific Expansion of Oxidative Stress by Glutathione Depletion and Use of a Fenton Nanoagent for Enhanced Chemodynamic Therapy. ACS Appl. Mater. Interfaces 2019, 11, 30551–30565. [Google Scholar] [CrossRef]
- Sun, F.; Peng, Y.; Li, Y.; Xu, M.; Cai, T. Fenton-Reaction-Triggered Metabolism of Acetaminophen for Enhanced Cancer Therapy. Chin. Chem. Lett. 2022, in press. [Google Scholar] [CrossRef]
- Zhong, X.; Bao, X.; Zhong, H.; Zhou, Y.; Zhang, Z.; Lu, Y.; Dai, Q.; Yang, Q.; Ke, P.; Xia, Y.; et al. Mitochondrial Targeted Drug Delivery Combined with Manganese Catalyzed Fenton Reaction for the Treatment of Breast Cancer. Int. J. Pharm. 2022, 622, 121810. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Chen, L.; Lv, R.; Liu, M.; He, N.; Wang, Z. A Metal–Phenolic Network-Based Multifunctional Nanocomposite with PH-Responsive ROS Generation and Drug Release for Synergistic Chemodynamic/Photothermal/Chemo-Therapy. J. Mater. Chem. B 2020, 8, 2177–2188. [Google Scholar] [CrossRef]
- Lei, M.; Chen, G.; Zhang, M.; Lei, J.; Li, T.; Li, D.; Zheng, H. A PH-Sensitive Drug Delivery System Based on Hyaluronic Acid Co-Deliver Doxorubicin and Aminoferrocene for the Combined Application of Chemotherapy and Chemodynamic Therapy. Colloids Surf. B Biointerfaces 2021, 203, 111750. [Google Scholar] [CrossRef]
- Chi, H.; Zhu, G.; Yin, Y.; Diao, H.; Liu, Z.; Sun, S.; Guo, Z.; Xu, W.; Xu, J.; Cui, C.; et al. Dual-Responsive Multifunctional “Core–Shell” Magnetic Nanoparticles Promoting Fenton Reaction for Tumor Ferroptosis Therapy. Int. J. Pharm. 2022, 622, 121898. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yang, J.; Zhou, L.; Hu, X.; Wang, C.; Chai, K.; Li, R.; Feng, L.; Sun, Y.; Dong, C.; et al. Dual-Responsive and ROS-Augmented Nanoplatform for Chemo/Photodynamic/Chemodynamic Combination Therapy of Triple Negative Breast Cancer. ACS Appl. Mater. Interfaces 2022, 14, 57–68. [Google Scholar] [CrossRef]
- Jia, C.; Wu, H.; Luo, K.; Hao, W.; Wang, S.; Huang, M. Magnetic Silica Nanosystems With NIR-Responsive and Redox Reaction Capacity for Drug Delivery and Tumor Therapy. Front. Chem. 2020, 8, 567652. [Google Scholar] [CrossRef]
- Pal, S.; Jana, N.R. Pharmacologic Vitamin C-Based Cell Therapy via Iron Oxide Nanoparticle-Induced Intracellular Fenton Reaction. ACS Appl. Nano Mater. 2020, 3, 1683–1692. [Google Scholar] [CrossRef]
- Wang, X.; Xu, S.; Zhang, L.; Cheng, X.; Yu, H.; Bao, J.; Lu, R. Vitamin C Induces Ferroptosis in Anaplastic Thyroid Cancer Cells by Ferritinophagy Activation. Biochem. Biophys. Res. Commun. 2021, 551, 46–53. [Google Scholar] [CrossRef]
- Ouyang, Q.; Kou, F.; Zhang, N.; Lian, J.; Tu, G.; Fang, Z. Tea Polyphenols Promote Fenton-like Reaction: PH Self-Driving Chelation and Reduction Mechanism. Chem. Eng. J. 2019, 366, 514–522. [Google Scholar] [CrossRef]
- Cheng, R.; Li, G.; Fan, L.; Jiang, J.; Zhao, Y. Therapeutic Iminoboronate-Based Polymersomes with a Cu(II)-Mediated Fenton Reaction-Enhanced ROS-Response. Chem. Commun. 2020, 56, 12246–12249. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, H.; Liu, H.; Yuan, Q.; Ren, F.; Han, Y.; Sun, Q.; Li, Z.; Gao, M. Boosting H2O2-Guided Chemodynamic Therapy of Cancer by Enhancing Reaction Kinetics through Versatile Biomimetic Fenton Nanocatalysts and the Second Near-Infrared Light Irradiation. Adv. Funct. Mater. 2020, 30, 1906128. [Google Scholar] [CrossRef]
- Sun, M.; Yang, D.; Sun, Q.; Jia, T.; Kuang, Y.; Gai, S.; He, F.; Zhang, F.; Yang, P. A Porous Material Excited by Near-Infrared Light for Photo/Chemodynamic and Photothermal Dual-Mode Combination Therapy. J. Mater. Chem. B 2020, 8, 10559–10576. [Google Scholar] [CrossRef]
- Jomova, K.; Hudecova, L.; Lauro, P.; Simunková, M.; Barbierikova, Z.; Malcek, M.; Alwasel, S.H.; Alhazza, I.M.; Rhodes, C.J.; Valko, M. The Effect of Luteolin on DNA Damage Mediated by a Copper Catalyzed Fenton Reaction. J. Inorg. Biochem. 2022, 226, 111635. [Google Scholar] [CrossRef]
- Simunkova, M.; Barbierikova, Z.; Jomova, K.; Hudecova, L.; Lauro, P.; Alwasel, S.H.; Alhazza, I.; Rhodes, C.J.; Valko, M. Antioxidant vs. Prooxidant Properties of the Flavonoid, Kaempferol, in the Presence of Cu(II) Ions: A ROS-Scavenging Activity, Fenton Reaction and DNA Damage Study. Int. J. Mol. Sci. 2021, 22, 1619. [Google Scholar] [CrossRef]
- Perron, N.R.; Wang, H.C.; DeGuire, S.N.; Jenkins, M.; Lawson, M.; Brumaghim, J.L. Kinetics of Iron Oxidation upon Polyphenol Binding. Dalt. Trans. 2010, 39, 9982–9987. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Edwards, A.M.; Ruiz, M.; Silva, E.; Lissi, E. Lysozyme Modification by the Fenton Reaction and Gamma Radiation. Free Radic. Res. 2002, 36, 277–284. [Google Scholar] [CrossRef]
- Bochi, G.V.; Torbitz, V.D.; Santos, R.C.V.; Cubillos-Rojas, M.; López, J.L.R.; Siebel, A.M.; Gomes, P.; de Oliveira, J.R.; Moresco, R.N. Fenton Reaction-Generated Advanced Oxidation Protein Products Induces Inflammation in Human Embryonic Kidney Cells. Inflammation 2016, 39, 1285–1290. [Google Scholar] [CrossRef]
- Ishikawa, S.; Yano, Y.; Arihara, K.; Itoh, M. Egg Yolk Phosvitin Inhibits Hydroxyl Radical Formation from the Fenton Reaction. Biosci. Biotechnol. Biochem. 2004, 68, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Voltea, S.; Karabagias, I.K.; Roussis, I.G. Use of Fe (II) and H2O2 along with Heating for the Estimation of the Browning Susceptibility of White Wine. Appl. Sci. 2022, 12, 4422. [Google Scholar] [CrossRef]
- Gharib-Bibalan, S.; Keramat, J.; Hamdami, N. Better Lime Purification of Raw Sugar Beet Juice by Advanced Fenton Oxidation Process. Ozone Sci. Eng. 2018, 40, 54–63. [Google Scholar] [CrossRef]
- Blank, I.; Pascual, E.C.; Devaud, S.; Fay, L.B.; Stadler, R.H.; Yeretzian, C.; Goodman, B.A. Degradation of the Coffee Flavor Compound Furfuryl Mercaptan in Model Fenton-Type Reaction Systems. J. Agric. Food Chem. 2002, 50, 2356–2364. [Google Scholar] [CrossRef] [PubMed]
- Yeung, Y.K.; Kang, Y.-R.; So, B.R.; Jung, S.K.; Chang, Y.H. Structural, Antioxidant, Prebiotic and Anti-Inflammatory Properties of Pectic Oligosaccharides Hydrolyzed from Okra Pectin by Fenton Reaction. Food Hydrocoll. 2021, 118, 106779. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, J.J.; Chen, L.; Peng, Z.; Zeng, Q.-H.; Zhu, Y.; Zhao, Y. Fenton Reaction-Assisted Photodynamic Inactivation of Calcined Melamine Sponge against Salmonella and Its Application. Food Res. Int. 2022, 151, 110847. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, C.K. A New Green Approach to Fenton’s Chemistry Using Tea Dregs and Coffee Grounds as Raw Material. Green Process. Synth. 2014, 3, 117–125. [Google Scholar] [CrossRef]
- Calhoun, L.N.; Kwon, Y.M. The Ferritin-like Protein Dps Protects Salmonella Enterica Serotype Enteritidis from the Fenton-Mediated Killing Mechanism of Bactericidal Antibiotics. Int. J. Antimicrob. Agents 2011, 37, 261–265. [Google Scholar] [CrossRef]
- Abbas, M.E.; Luo, W.; Zhu, L.; Zou, J.; Tang, H. Fluorometric Determination of Hydrogen Peroxide in Milk by Using a Fenton Reaction System. Food Chem. 2010, 120, 327–331. [Google Scholar] [CrossRef]
- Wang, S.; Fang, B.; Yuan, M.; Wang, Z.; Peng, J.; Lai, W. Dual-Mode Immunoassay System Based on Glucose Oxidase-Triggered Fenton Reaction for Qualitative and Quantitative Detection of Danofloxacin in Milk. J. Dairy Sci. 2020, 103, 7826–7833. [Google Scholar] [CrossRef]
- Wu, K.; Ying, W.; Shi, Z.; Yang, H.; Zheng, Z.; Zhang, J.; Yang, J. Fenton Reaction-Oxidized Bamboo Lignin Surface and Structural Modification to Reduce Nonproductive Cellulase Binding and Improve Enzyme Digestion of Cellulose. ACS Sustain. Chem. Eng. 2018, 6, 3853–3861. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, M.-J. Enhancing Enzymolysis and Fermentation Efficiency of Sugarcane Bagasse by Synergistic Pretreatment of Fenton Reaction and Sodium Hydroxide Extraction. Bioresour. Technol. 2016, 214, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Midassi, S.; Bedoui, A.; Bensalah, N. Efficient Degradation of Chloroquine Drug by Electro-Fenton Oxidation: Effects of Operating Conditions and Degradation Mechanism. Chemosphere 2020, 260, 127558. [Google Scholar] [CrossRef]
- Marchetti, M.D.; Bessa Azevedo, E. Degradation of NSAIDs by Optimized Photo-Fenton Process Using UV-LEDs at near-Neutral PH. J. Water Process Eng. 2020, 35, 101171. [Google Scholar] [CrossRef]
- Barros, W.R.P.; Franco, P.C.; Steter, J.R.; Rocha, R.S.; Lanza, M.R.V. Electro-Fenton Degradation of the Food Dye Amaranth Using a Gas Diffusion Electrode Modified with Cobalt (II) Phthalocyanine. J. Electroanal. Chem. 2014, 722–723, 46–53. [Google Scholar] [CrossRef]
- Shin, J.; Bae, S.; Chon, K. Fenton Oxidation of Synthetic Food Dyes by Fe-Embedded Coffee Biochar Catalysts Prepared at Different Pyrolysis Temperatures: A Mechanism Study. Chem. Eng. J. 2021, 421, 129943. [Google Scholar] [CrossRef]
- Chen, Q.; Espey, M.G.; Sun, A.Y.; Pooput, C.; Kirk, K.L.; Krishna, M.C.; Khosh, D.B.; Drisko, J.; Levine, M. Pharmacologic Doses of Ascorbate Act as a Prooxidant and Decrease Growth of Aggressive Tumor Xenografts in Mice. Proc. Natl. Acad. Sci. USA 2008, 105, 11105–11109. [Google Scholar] [CrossRef] [PubMed]
- Ngo, B.; Van Riper, J.M.; Cantley, L.C.; Yun, J. Targeting Cancer Vulnerabilities with High-Dose Vitamin C. Nat. Rev. Cancer 2019, 19, 271–282. [Google Scholar] [CrossRef]
- Yun, J.; Mullarky, E.; Lu, C.; Bosch, K.N.; Kavalier, A.; Rivera, K.; Roper, J.; Chio, C.I.I.; Giannopoulou, E.G.; Rago, C.; et al. Vitamin C Selectively Kills KRAS and BRAF Mutant Colorectal Cancer Cells by Targeting GAPDH. Science 2015, 350, 1391–1396. [Google Scholar] [CrossRef]
- Abe, C.; Higuchi, O.; Matsumoto, A.; Miyazawa, T. Determination of Intracellular Ascorbic Acid Using Tandem Mass Spectrometry. Analyst 2022, 147, 2640–2643. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abe, C.; Miyazawa, T.; Miyazawa, T. Current Use of Fenton Reaction in Drugs and Food. Molecules 2022, 27, 5451. https://doi.org/10.3390/molecules27175451
Abe C, Miyazawa T, Miyazawa T. Current Use of Fenton Reaction in Drugs and Food. Molecules. 2022; 27(17):5451. https://doi.org/10.3390/molecules27175451
Chicago/Turabian StyleAbe, Chizumi, Taiki Miyazawa, and Teruo Miyazawa. 2022. "Current Use of Fenton Reaction in Drugs and Food" Molecules 27, no. 17: 5451. https://doi.org/10.3390/molecules27175451
APA StyleAbe, C., Miyazawa, T., & Miyazawa, T. (2022). Current Use of Fenton Reaction in Drugs and Food. Molecules, 27(17), 5451. https://doi.org/10.3390/molecules27175451