

Evolution of Interferon-Gamma Aptamer with Good Affinity and Analytical Utility by a Rational In Silico Base Mutagenesis Post-SELEX Strategy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Production of Evolutionary Aptamer against IFN-γ by In Silico Base Mutagenesis Post-SELEX
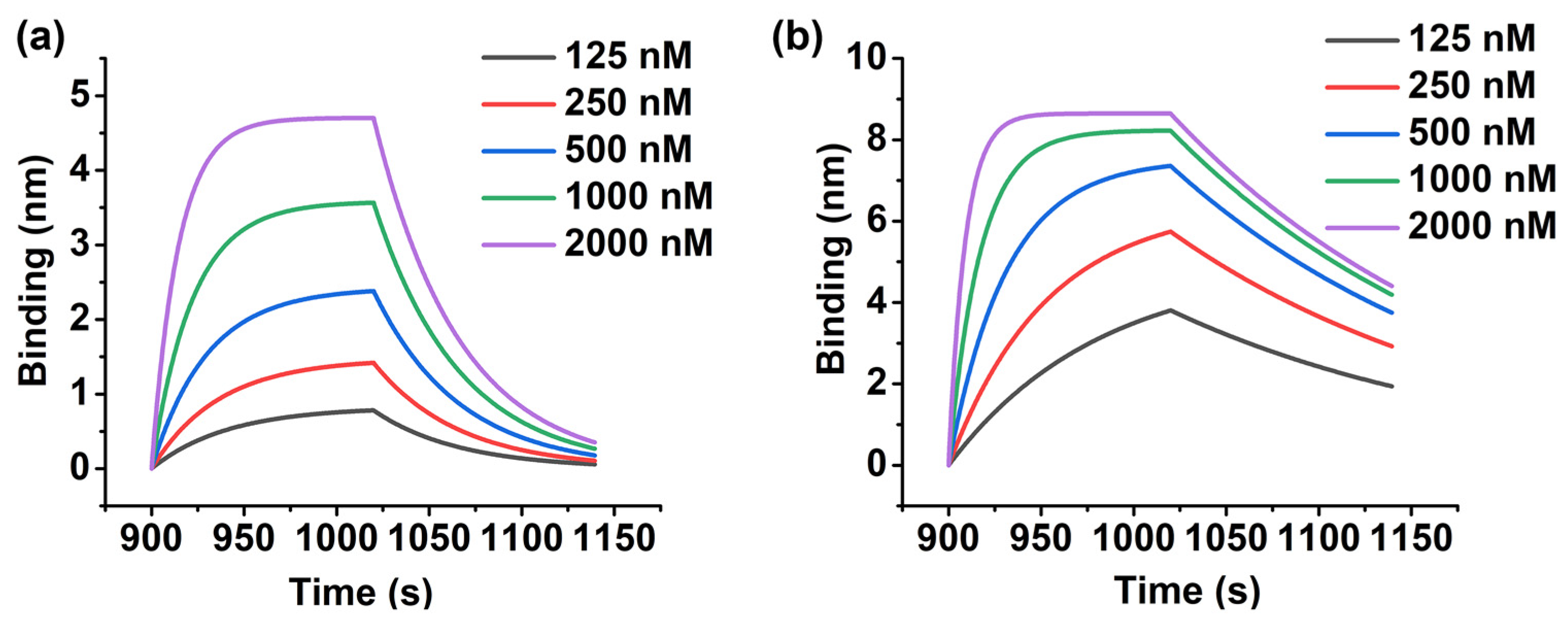
2.2. Generation of Evolutionary Aptamer with High Affinity
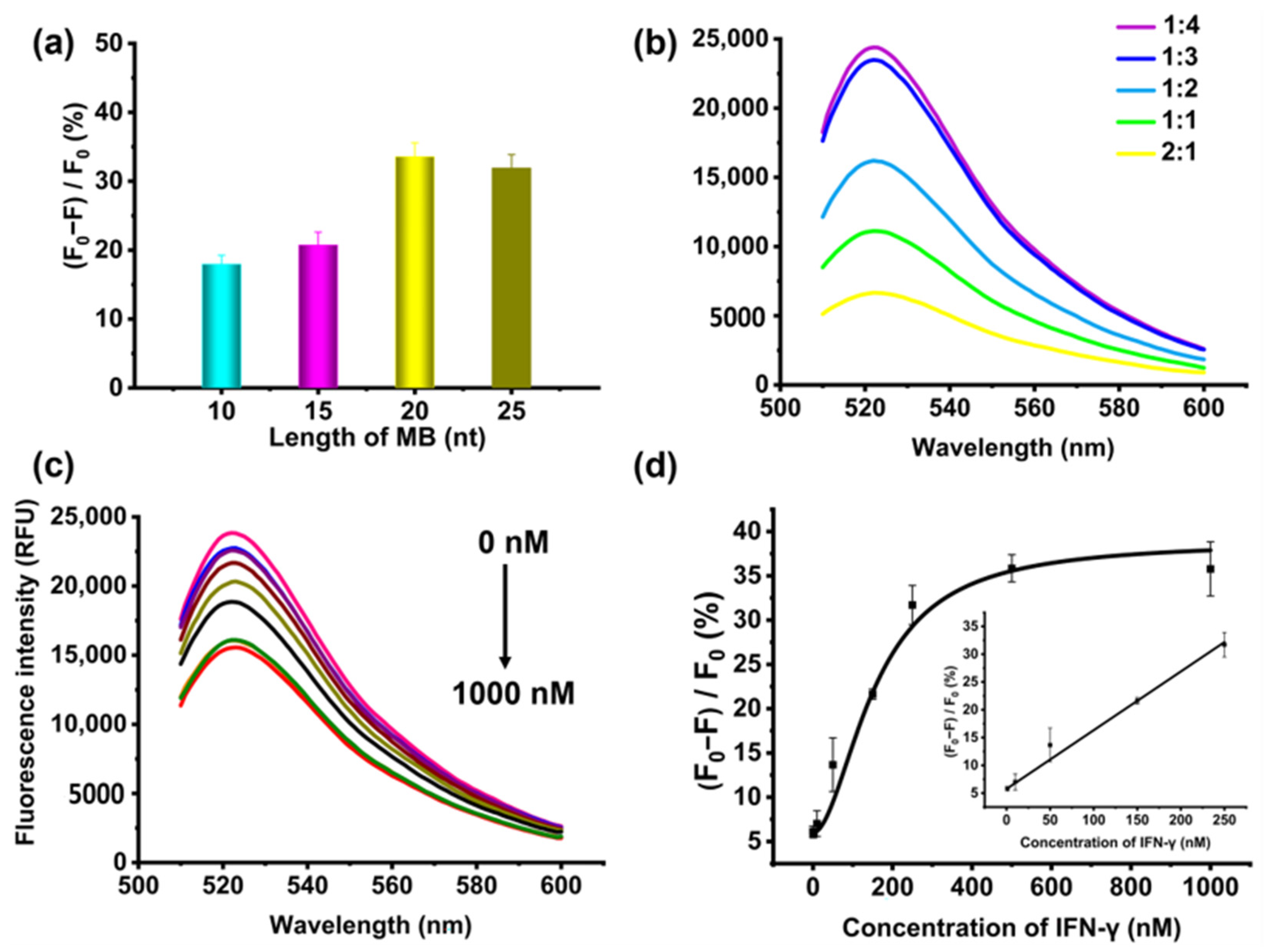
2.3. Detection of IFN-γ Using Evolutionary Aptamer
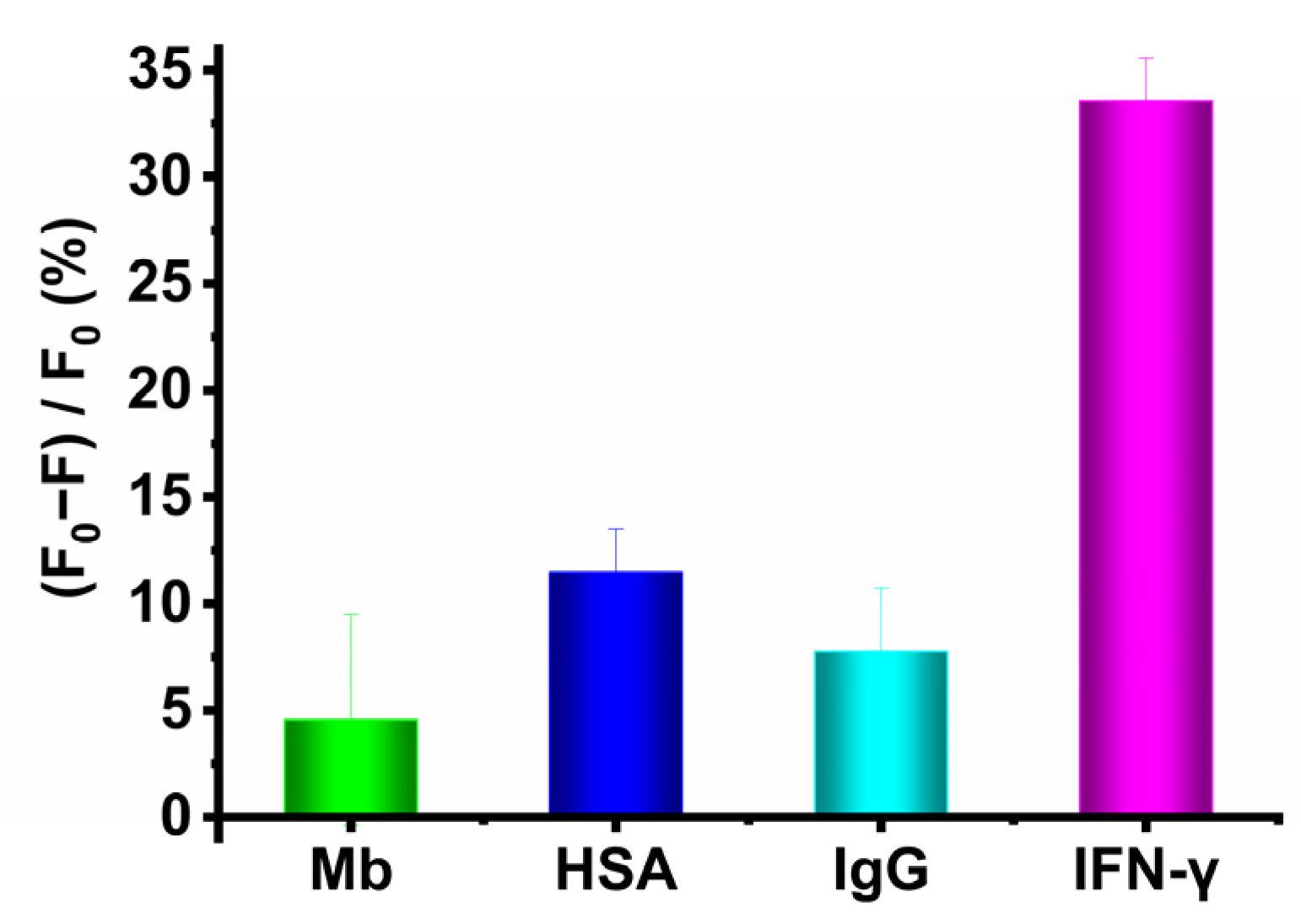
2.4. High Selectivity of the Assay Based on Evolutionary Aptamer
3. Materials and Methods
3.1. Materials and Instruments
3.2. Production of Aptamer Mutants by In Silico Mutagenesis
3.3. Model Simulation of Aptamer/IFN-γ
3.4. Biolayer Interferometry Assay
3.5. Circular Dichroism Measurement
3.6. Fluorescence Detection of IFN-γ Using Evolutionary Aptamer
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Klug, S.J.; Famulok, M. All you wanted to know about SELEX. Mol. Biol. Rep. 1994, 20, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.X.; Zheng, X.; Jiao, B.H.; Wang, L.H. Post-SELEX optimization of aptamers. Anal. Bioanal. Chem. 2016, 408, 4567–4573. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.A.; Zulkifli, R.M.; Hussin, H.; Nadri, M.H. In silico approach for Post-SELEX DNA aptamers: A mini-review. J. Mol. Graph. Model. 2021, 105, 107872. [Google Scholar] [CrossRef] [PubMed]
- Navien, T.N.; Thevendran, R.; Hamdani, H.Y.; Tang, T.H.; Citartan, M. In silico molecular docking in DNA aptamer development. Biochimie 2021, 180, 54–67. [Google Scholar] [CrossRef]
- Song, M.H.; Li, G.; Zhang, Q.; Liu, J.P.; Huang, Q. De novo post-SELEX optimization of a G-quadruplex DNA aptamer binding to marine toxin gonyautoxin 1/4. Comput. Struct. Biotechnol. J. 2020, 10, 3425–3433. [Google Scholar] [CrossRef]
- Hsieh, P.C.; Lin, H.T.; Chen, W.Y.; Tsai, J.J.P.; Hu, W.P. The combination of computational and biosensing technologies for selecting aptamer against prostate specific antigen. Biomed. Res. Int. 2017, 2017, 5041683. [Google Scholar] [CrossRef]
- Wang, Q.L.; Cui, H.F.; Du, J.F.; Lv, Q.Y.; Song, X.J. In silico post-SELEX screening and experimental characterizations for acquisition of high affinity DNA aptamers against carcinoembryonic antigen. RSC. Adv. 2019, 9, 6328–6334. [Google Scholar] [CrossRef]
- Mousivand, M.; Anfossi, L.; Bagherzadeh, K.; Barbero, N.; Mirzadi-Gohari, A.; Javan-Nikkhah, M. In silico maturation of affinity and selectivity of DNA aptamers against aflatoxin B1 for biosensor development. Anal. Chim. Acta 2020, 1105, 178–186. [Google Scholar] [CrossRef]
- Setlem, K.; Mondal, B.; Ramlal, S.; Kingston, J. Immuno affinity SELEX for simple, rapid, and cost-effective aptamer enrichment and identification against aflatoxin B1. Front. Microbiol. 2016, 1, 1909. [Google Scholar] [CrossRef] [PubMed]
- Thiel, W.H.; Bair, T.; Wyatt Thiel, K.; Dassie, J.P.; Rockey, W.M.; Howell, C.A.; Liu, X.Y.; Dupuy, A.J.; Huang, L.; Owczarzy, R.; et al. Nucleotide Cbserved with a short SELEX RNA aptamer library. Nucleic Acid Ther. 2011, 21, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Chen, J.; Chen, X.; Wang, X.; Zhou, N. Systematic screening and optimization of single-stranded DNA aptamer specific for N-acetylneuraminic acid: A comparative study. Sens. Actuators B—Chem. 2021, 344, 130270. [Google Scholar] [CrossRef]
- Kuai, H.L.; Zhao, Z.L.; Mo, L.T.; Liu, H.; Hu, X.X.; Fu, T.; Zhang, X.B.; Tan, W.H. Circular bivalent aptamers enable in vivo stability and recognition. J. Am. Chem. Soc. 2017, 139, 9128–9131. [Google Scholar] [CrossRef]
- Zhao, L.H.; Qi, X.Y.; Yan, X.C.; Huang, Y.F.; Liang, X.G.; Zhang, L.Q.; Wang, S.; Tan, W.H. Engineering Aptamer with enhanced affinity by triple helix-based terminal fixation. J. Am. Chem. Soc. 2019, 141, 17493–17497. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.Y.; Wang, S.; Zhao, L.H.; Yang, J.B.; Xu, Y. Some frontier technologies for aptamers in medical applications. In Aptamers for Medical Applications: From Diagnosis to Therapeutics; Dong, Y., Ed.; Springer: Singapore, 2021; pp. 375–403. [Google Scholar]
- Boehm, U.; Klamp, T.; Groot, A.M.; Howard, J.C. Cellular responses to interferon-gamma. Annu. Rev. Immunol. 1997, 15, 749–795. [Google Scholar] [CrossRef]
- Dhenadhayalan, N.; Sriram, M.I.; Lin, K.C. Aptamer-based fluorogenic sensing of interferon-gamma probed with ReS2 and TiS2 nanosheets. Sens. Actuators B—Chem. 2017, 258, 929–936. [Google Scholar] [CrossRef]
- Januarie, K.C.; Uhuo, O.V.; Iwuoha, E.; Feleni, U. Recent advances in the detection of interferon-gamma as a TB biomarker. Anal. Bioanal. Chem. 2022, 414, 907–921. [Google Scholar] [CrossRef]
- Kulabhusan, P.K.; Hussain, B.; Yüce, M. Current perspectives on aptamers as diagnostic tools and therapeutic agents. Pharmaceutics 2020, 12, 646. [Google Scholar] [CrossRef]
- Cao, B.B.; Hu, Y.; Duan, J.H.; Ma, J.; Xu, D.K.; Yang, X.D. Selection of a novel DNA aptamer for assay of intracellular interferon-gamma. PLoS ONE 2014, 9, e98214. [Google Scholar] [CrossRef]
- Li, Y.Y.; Wen, H.; Shi, X.B.; Li, H.T. Determination of protein-DNA (ZMYND11-DNA) interaction by a label-free biolayer interferometry assay. Bio-Protocol 2015, 5, e1402. [Google Scholar] [CrossRef]
- Tuleuova, N.; Jones, C.N.; Yan, J.; Ramanculov, E.; Yokobayashi, Y.; Revzin, A. Development of an aptamer beacon for detection of interferon-gamma. Anal. Chem. 2010, 82, 1851–1857. [Google Scholar] [CrossRef]
- Abnous, K.; Danesh, N.M.; Ramezani, M.; Alibolandi, M.; Hassanabad, K.Y.; Emrani, A.S.; Bahreyni, A.; Taghdisi, S.M. A triple-helix molecular switch-based electrochemical aptasensor for interferon-gamma using a gold electrode and Methylene Blue as a redox probe. Microchim. Acta 2017, 184, 4151–4157. [Google Scholar] [CrossRef]
- Jin, H.; Gui, R.J.; Gao, X.H.; Sun, Y.J. An amplified label-free electrochemical aptasensor of γ-interferon based on target-induced DNA strand transform of hairpin-to-linear conformation enabling simultaneous capture of redox probe and target. Biosens. Bioelectron. 2019, 145, 111732. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- SantaLucia, J., Jr. A unified view of polymer, dumbbell, and oligonucleotide DNA nearest-neighbor thermodynamics. Proc. Natl. Acad. Sci. USA 1998, 95, 1460–1465. [Google Scholar] [CrossRef]
- Maciej, A.; Mariusz, P.; Tomasz, Z.; Joanna, S.; Tomasz, R.; Katarzyna, T.; Adamiak, R.W.; Marta, S. New functionality of RNAComposer: An application to shape the axis of MiR160 precursor structure. Acta Biochim. Pol. 2016, 63, 737–744. [Google Scholar]
- Mariusz, P.; Marta, S.; Maciej, A.; Purzycka, K.J.; Piotr, L.; Natalia, B.; Jacek, B.; Adamiak, R.W. Automated 3D structure composition for large RNAs. Nucleic Acids Res. 2012, 40, e112. [Google Scholar]
- Thiel, D.J.; Du, M.; Walter, R.L.; Arcy, A.D.; Chène, C.; Fountoulakis, M.; Garotta, G.; Winkler, F.K.; Ealick, S.E. Observation of an unexpected third receptor molecule in the crystal structure of human interferon-γ receptor complex. Structure 2000, 8, 927–936. [Google Scholar] [CrossRef]
- Pierce, B.G.; Kevin, W.; Howook, H.; Bong-Hyun, K.; Thom, V.; Weng, Z. ZDOCK server: Interactive docking prediction of protein–protein complexes and symmetric multimers. Bioinformatics 2014, 30, 1771–1773. [Google Scholar] [CrossRef]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Wang, Q.; Yin, Y.; Yang, Y.; Cui, H.; Dong, Y. Evolution of Interferon-Gamma Aptamer with Good Affinity and Analytical Utility by a Rational In Silico Base Mutagenesis Post-SELEX Strategy. Molecules 2022, 27, 5725. https://doi.org/10.3390/molecules27175725
Zhao L, Wang Q, Yin Y, Yang Y, Cui H, Dong Y. Evolution of Interferon-Gamma Aptamer with Good Affinity and Analytical Utility by a Rational In Silico Base Mutagenesis Post-SELEX Strategy. Molecules. 2022; 27(17):5725. https://doi.org/10.3390/molecules27175725
Chicago/Turabian StyleZhao, Lianhui, Qionglin Wang, Yingai Yin, Yan Yang, Huifang Cui, and Yiyang Dong. 2022. "Evolution of Interferon-Gamma Aptamer with Good Affinity and Analytical Utility by a Rational In Silico Base Mutagenesis Post-SELEX Strategy" Molecules 27, no. 17: 5725. https://doi.org/10.3390/molecules27175725
APA StyleZhao, L., Wang, Q., Yin, Y., Yang, Y., Cui, H., & Dong, Y. (2022). Evolution of Interferon-Gamma Aptamer with Good Affinity and Analytical Utility by a Rational In Silico Base Mutagenesis Post-SELEX Strategy. Molecules, 27(17), 5725. https://doi.org/10.3390/molecules27175725