


A Novel 5-Chloro-N-phenyl-1H-indole-2-carboxamide Derivative as Brain-Type Glycogen Phosphorylase Inhibitor: Potential Therapeutic Effect on Cerebral Ischemia

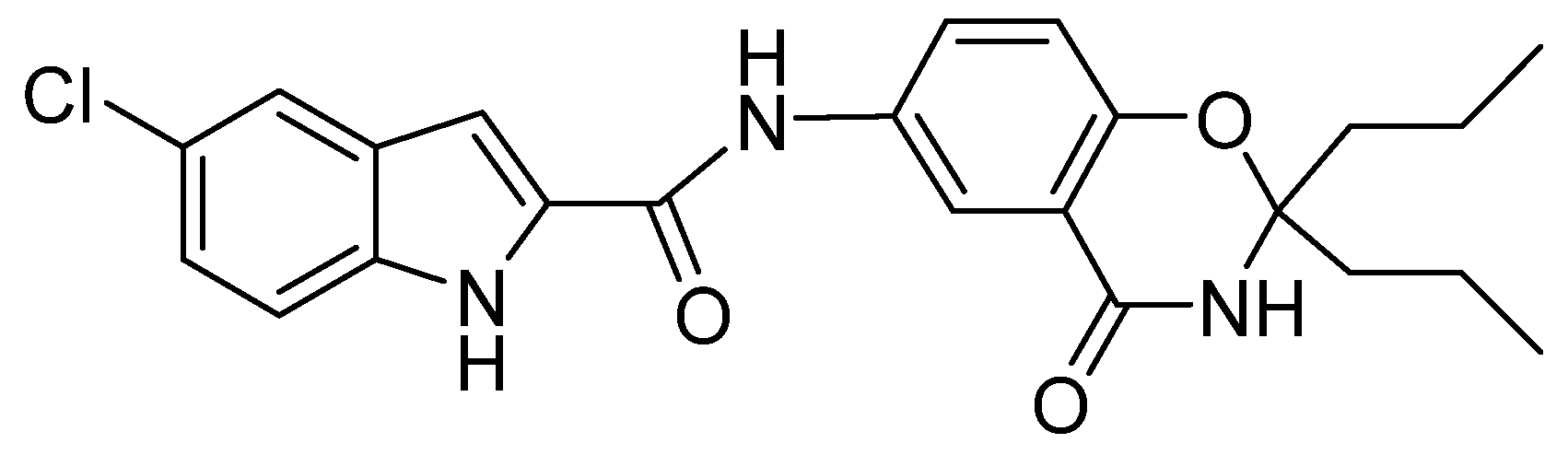{kind=link}
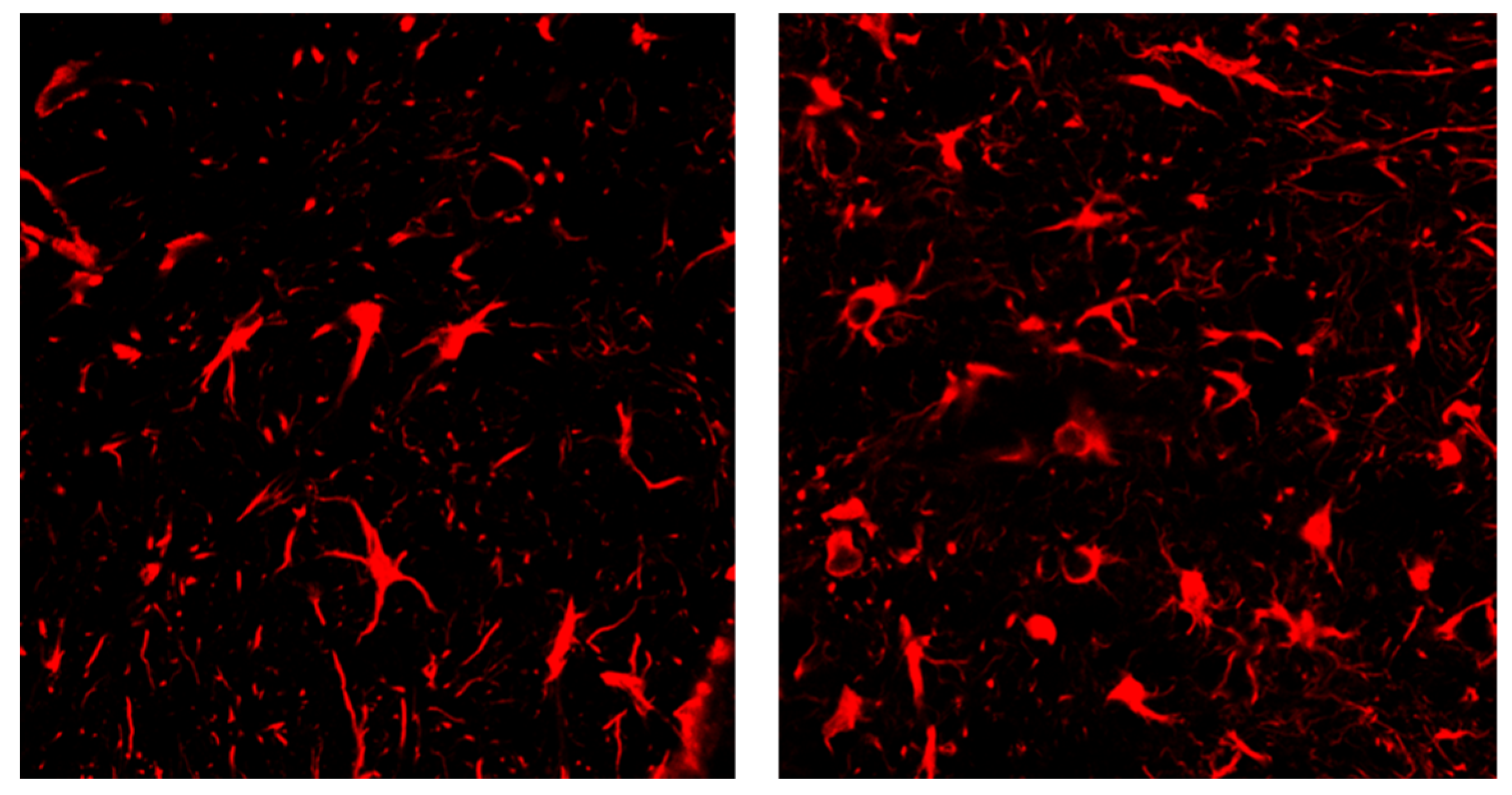{kind=link}
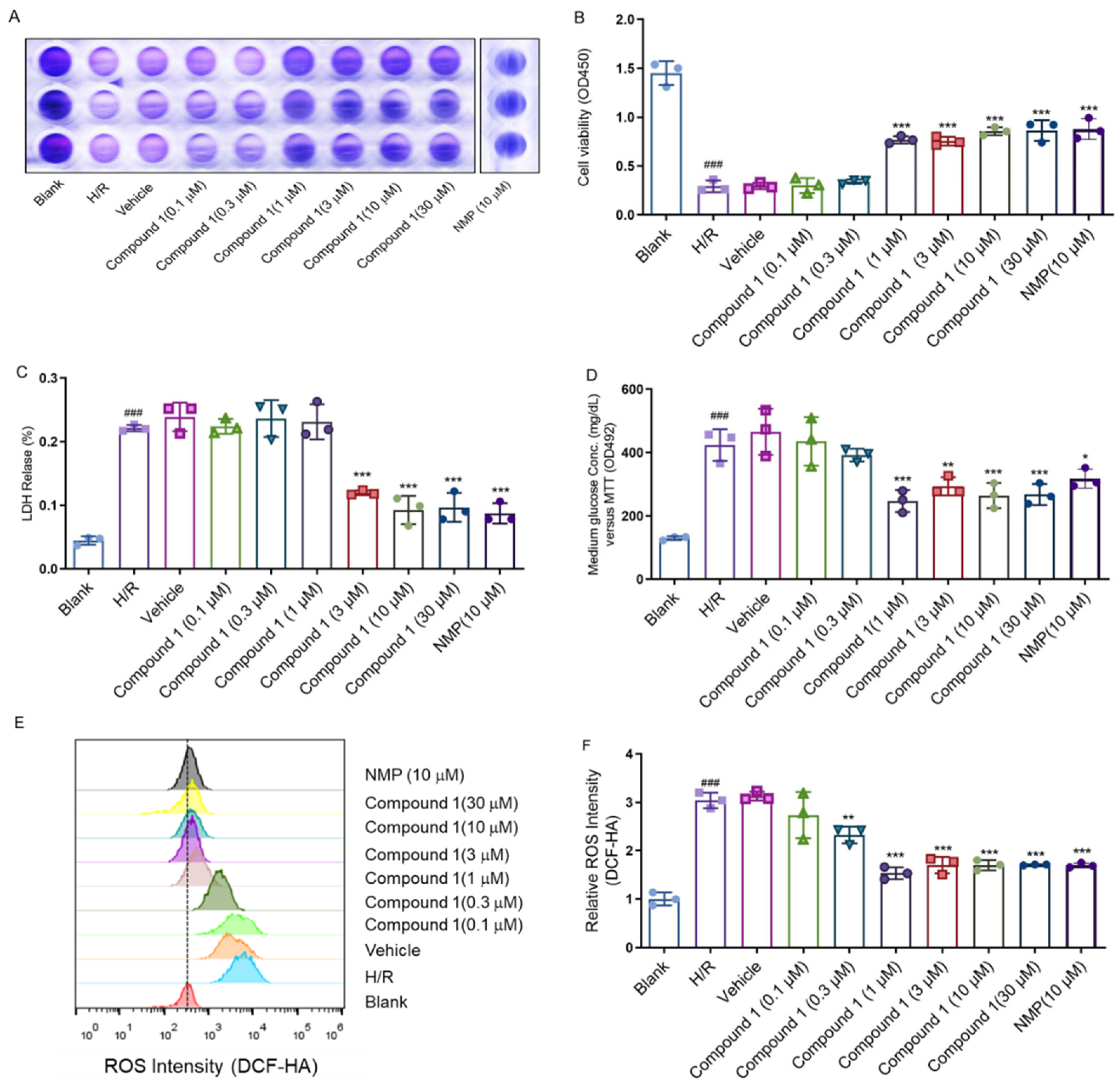{kind=link}
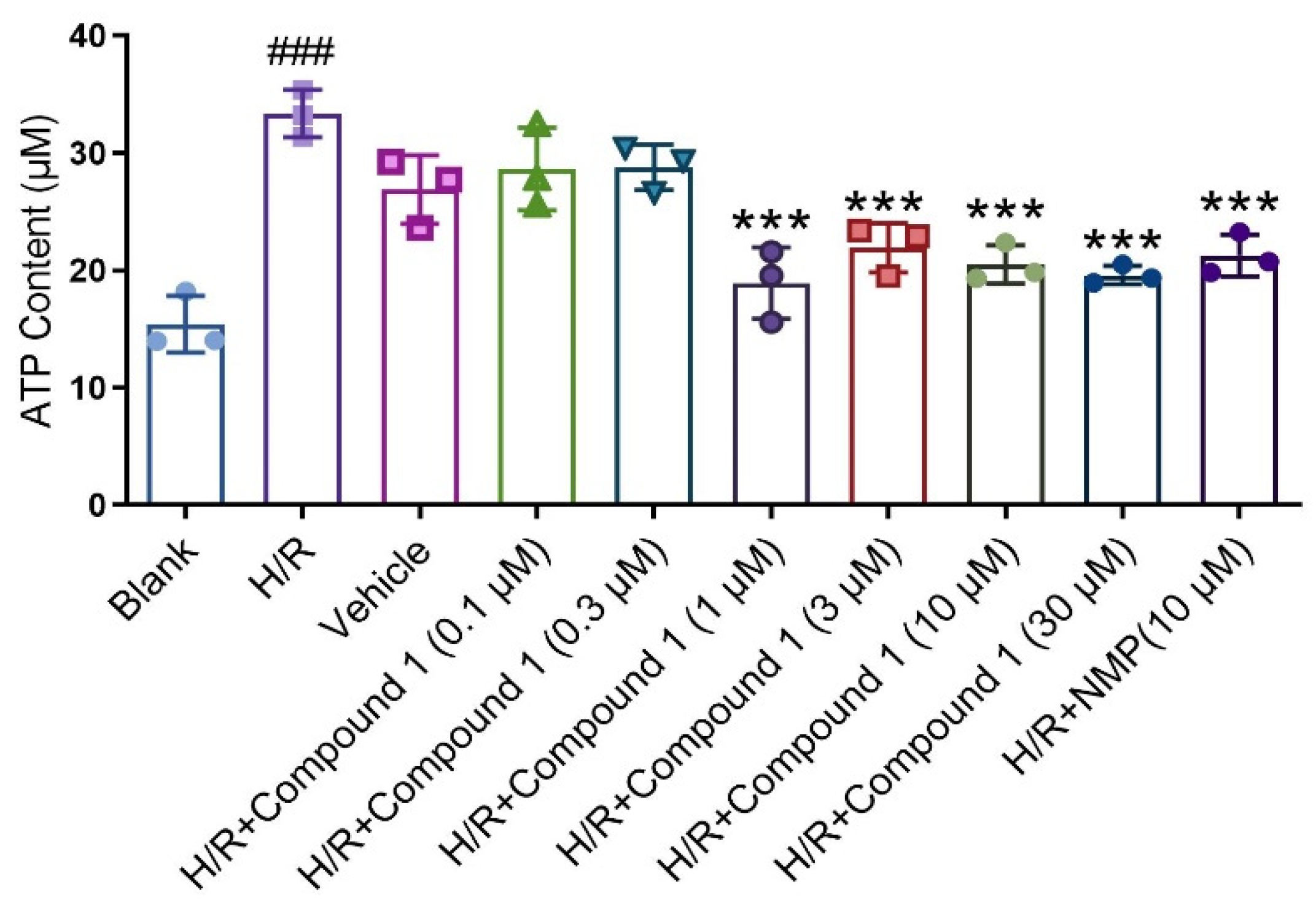{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Protective Effect of Compound 1 on H/R Injury of Brain Astrocytes
2.2. Compound 1 Improves Mouse Astrocyte Energy Metabolism
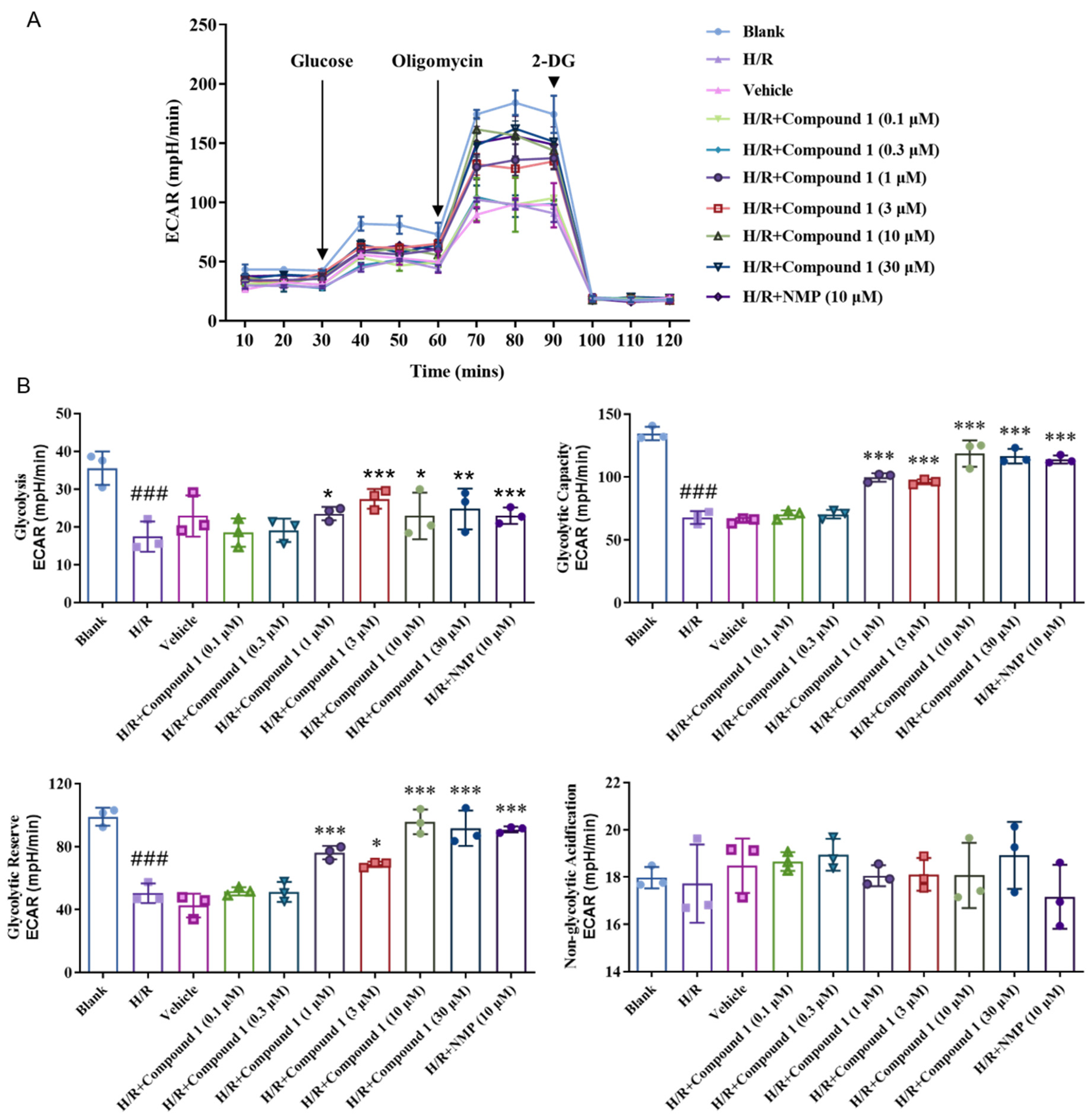
2.3. Compound 1 Significantly Downregulates the Degree of Extracellular Acidification and Ameliorates Metabolic Acidosis
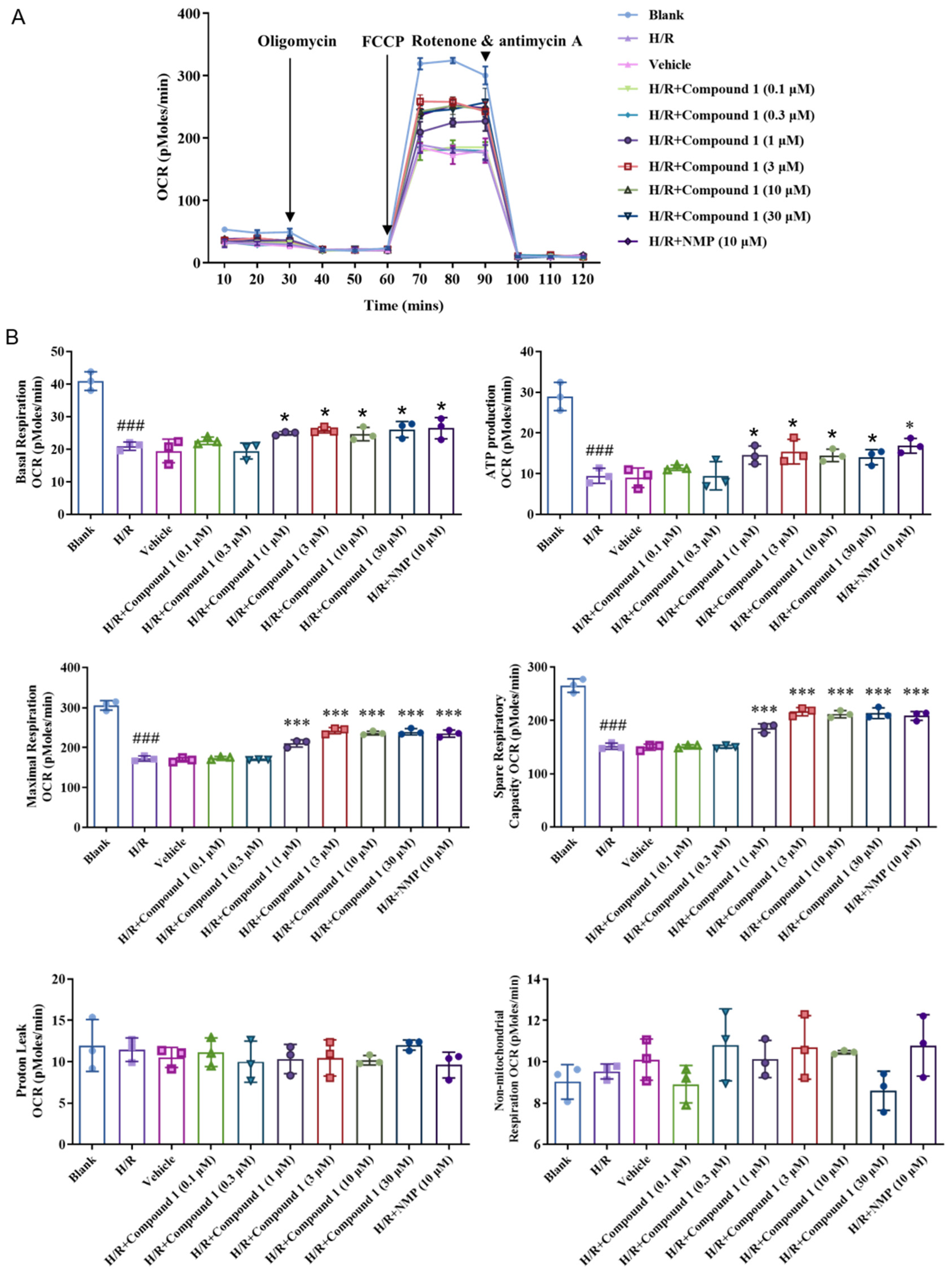
2.4. Compound 1 Significantly Increases Mitochondrial Aerobic Energy Metabolism and Decreases Anaerobic Glycolysis
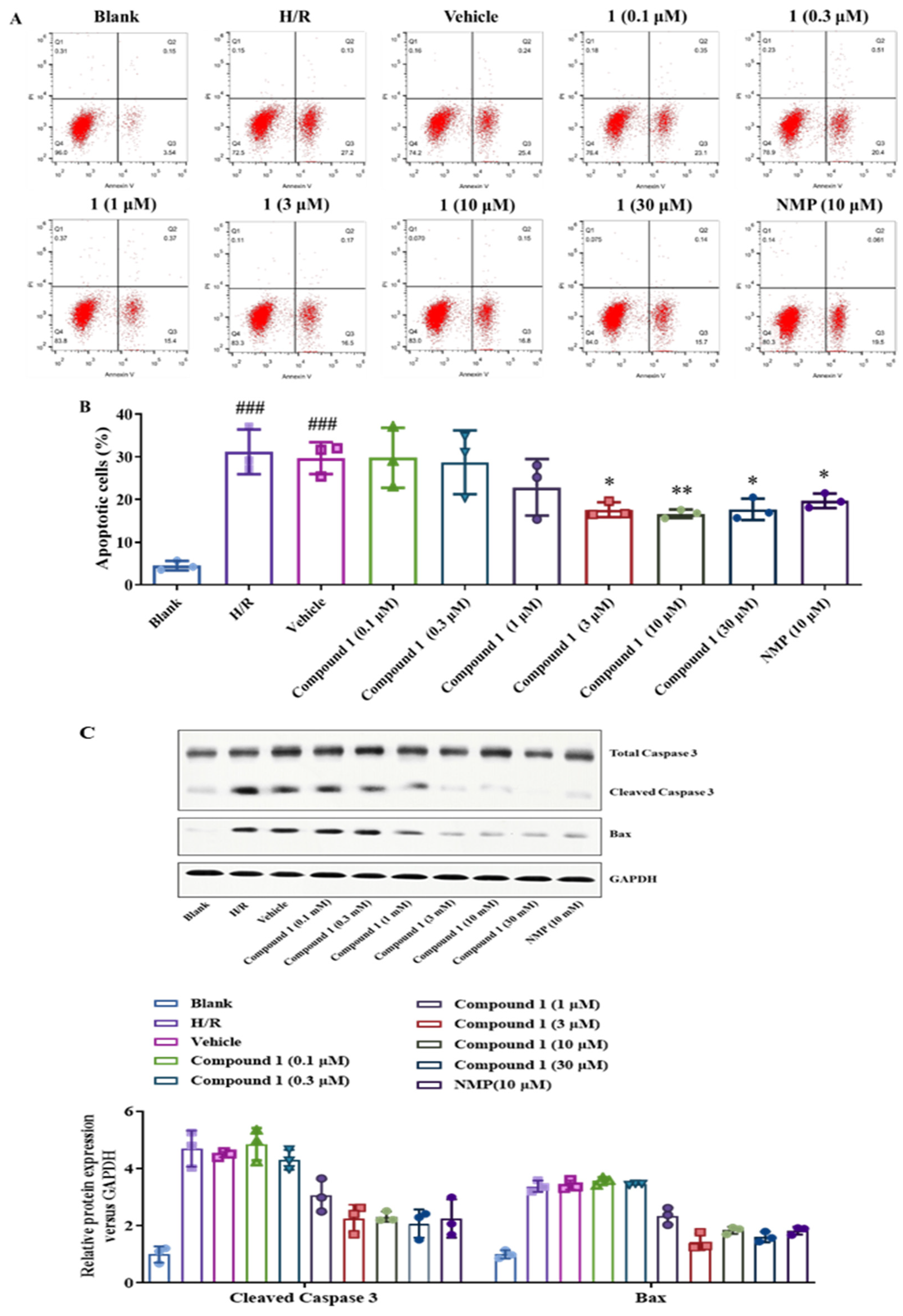
2.5. Proportion of Compound 1 Inhibits Apoptosis and the Expression of Apoptosis-Related Proteins
3. Materials and Methods
3.1. Animals
3.2. Cell Culture
3.3. Experiment Grouping
3.4. CCK-8 Assay
3.5. LDH Release
3.6. Medium Glucose Content
3.7. ROS Release
3.8. ATP Content
3.9. Mitochondrial Oxidative Respiratory Chain Function
3.10. Apoptosis Analysis
3.11. Western Blotting
3.12. Materials
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ou-Yang, L.; Liu, Y.; Wang, B.-Y.; Cao, P.; Zhang, J.-J.; Huang, Y.-Y.; Shen, Y.; Lyu, J.-X. Carnosine suppresses oxygen-glucose deprivation/recovery-induced proliferation and migration of reactive astrocytes of rats in vitro. Acta Pharmacol. Sin. 2018, 39, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Bastian, C.; Quinn, J.; Doherty, C.; Franke, C.; Faris, A.; Brunet, S.; Baltan, S. Role of brain glycogen during ischemia, aging and cell-to-cell interactions. Brain Glycogen Metab. 2019, 23, 347–361. [Google Scholar]
- Zhang, H.Y.; Zhao, Y.N.; Wang, Z.L.; Huang, Y.F. Chronic corticosterone exposure reduces hippocampal glycogen level and induces depression-like behavior in mice. J. Zhejiang Univ. 2015, 16, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.I.; Roulston, C.L.; Stapleton, D.I. Molecular basis of impaired glycogen metabolism during ischemic stroke and hypoxia. PLoS ONE 2014, 9, e97570. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Guo, H.; Fan, Z.; Zhang, X.; Wu, D.; Tang, W.; Gu, T.; Wang, S.; Yin, A.; Tao, L.; et al. Glycogenolysis is crucial for astrocytic glycogen accumulation and brain damage after reperfusion in ischemic stroke. IScience 2020, 23, 101136. [Google Scholar] [CrossRef]
- Dienel, G.A.; Hertz, L.J.G. Astrocytic contributions to bioenergetics of cerebral ischemia. Glia 2005, 50, 362–388. [Google Scholar] [CrossRef]
- Prebil, M.; Jensen, J.; Zorec, R.; Kreft, M. Astrocytes and energy metabolism. Arch. Physiol. Biochem. 2011, 117, 64–69. [Google Scholar] [CrossRef]
- Brown, A.M.; Rich, L.R.; Ransom, B.R. Metabolism of glycogen in brain white matter. Brain Glycogen Metab. 2019, 23, 187–207. [Google Scholar]
- Dienel, G.A.; Cruz, N.F. Astrocyte activation in working brain: Energy supplied by minor substrates. Neurochem. Int. 2006, 48, 586–595. [Google Scholar] [CrossRef]
- Mathieu, C.; de la Sierra-Gallay, I.L.; Duval, R.; Xu, X.; Cocaign, A.; Léger, T.; Woffendin, G.; Camadro, J.M.; Etchebest, C.; Haouz, A.; et al. Insights into brain glycogen metabolism: The structure of human brain glycogen phosphorylase. J. Biol. Chem. 2016, 291, 18072–18083. [Google Scholar] [CrossRef]
- Yan, Z.; Li, S.; Wang, Y.; Li, J.; Ma, C.; Guo, Y.; Zhang, L. Discovery of novel heterocyclic derivatives as potential glycogen phosphorylase inhibitors with a cardioprotective effect. Bioorg. Chem. 2022, 129, 106120. [Google Scholar] [CrossRef] [PubMed]
- Haile, M.; Limson, F.; Gingrich, K.; Li, Y.-S.; Quartermain, D.; Blanck, T.; Bekker, A. Nimodipine prevents transient cognitive dysfunction after moderate hypoxia in adult mice. J. Neurosurg. Anesthesiol. 2009, 21, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Biala, G.; Kruk-Slomka, M.; Jozwiak, K. Influence of acute or chronic calcium channel antagonists on the acquisition and consolidation of memory and nicotine-induced cognitive effects in mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2013, 386, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.B. Intracellular brain pH and ischemic vasoconstriction in the white New Zealand rabbit. Stroke 1990, 21, IV117–IV119. [Google Scholar]
- Yang, J.; Qi, J.; Xiu, B.; Yang, B.; Niu, C.; Yang, H. Reactive oxygen species play a biphasic role in brain ischemia. J. Investig. Surg. 2018, 32, 97–102. [Google Scholar] [CrossRef]
- Letteria, M.; Domenico, P.; Mariagrazia, R.; Natasha, I.; Herbert, M.; Vincenzo, A.; Alessandra, B.; Giovanni, C.; Antonina, P.; Francesco, S.; et al. Cellular longevity, ROS-mediated NLRP3 inflammasome activation in brain, heart, kidney, and testis ischemia/reperfusion injury. Oxid. Med. Cell. Longev. 2016, 2016, 2183026. [Google Scholar]
- Vannucci, R.C.; Brucklacher, R.M.; Vannucci, S.J. The effect of hyperglycemia on cerebral metabolism during hypoxia-ischemia in the immature rat. J. Cereb. Blood Flow Metab. 1996, 16, 1026–1033. [Google Scholar] [CrossRef]
- Blanco, M.M.B.; Prashant, G.N.; Vespa, P.M. Cerebral metabolism and the role of glucose control in acute traumatic brain injury. Neurosurg. Clin. N. Am. 2016, 27, 453–463. [Google Scholar] [CrossRef]
- Yu, S.; Xu, H.; Chi, X.; Wei, L.; Cheng, Q.; Yang, Y.; Zhou, C.; Ding, F.J.N. 2-(4-methoxyphenyl)ethyl-2-acetamido-2-deoxy-β-d-pyranoside (a salidroside analog) confers neuroprotection with a wide therapeutic window by regulating local glucose metabolism in a rat model of cerebral ischemic injury. Neuroscience 2018, 391, 60–72. [Google Scholar] [CrossRef]
- Zhen, X.; Cao, B.Q.; Tao, W.; Qi, L.; Kang, T.; Ge, C.Y.; Gao, W.J.; Hao, H.J. LanCL1 attenuates ischemia-induced oxidative stress by Sirt3-mediated preservation of mitochondrial function. Brain Res. Bull. 2018, 142, 216–223. [Google Scholar]
- Rao, J.; Oz, G.; Seaquist, E.R. Regulation of cerebral glucose metabolism. Minerva Endocrinol. 2006, 31, 149. [Google Scholar] [PubMed]
- Veluthakal, R.; Kaetzel, D.; Kowluru, A. Biochemistry, Nm23-H1 regulates glucose-stimulated insulin secretion in pancreatic β-cells via Arf6-Rac1 signaling axis. Cell. Physiol. Biochem. 2013, 32, 533–541. [Google Scholar] [CrossRef] [PubMed]
- De Cristóbal, J.; Cárdenas, A.; Lizasoain, I.; Leza, J.C.; Fernández-Tomé, P.; Lorenzo, P.; Moro, M. Inhibition of glutamate release via recovery of ATP levels accounts for a neuroprotective effect of aspirin in rat cortical neurons exposed to oxygen-glucose deprivation. Stroke 2002, 33, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Olah, J.; Klivenyi, P.; Gardian, G.; Vécsei, L.; Orosz, F.; Kovacs, G.G.; Westerhoff, H.V.; Ovadi, J. Increased glucose metabolism and ATP level in brain tissue of Huntington’s disease transgenic mice. FEBS J. 2008, 275, 4740–4755. [Google Scholar] [CrossRef]
- Nielsen, T.H.; Bindslev, T.T.; Pedersen, S.M.; Toft, P.; Olsen, N.V.; Nordström, C.H. Cerebral energy metabolism during induced mitochondrial dysfunction. Acta Anaesthesiol. Scand. 2013, 57, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.H.; Lee, D.Y.; Basith, S.; Manavalan, B.; Paik, M.J.; Rybinnik, I.; Mouradian, M.M.; Ahn, J.H.; Lee, G. Metabolome changes in cerebral ischemia. Cells 2020, 9, 1630. [Google Scholar] [CrossRef]
- Golias, T.; Papandreou, I.; Sun, R.; Kumar, B.; Brown, N.V.; Swanson, B.J.; Pai, R.; Jaitin, D.; Le, Q.T.; Teknos, T.N.; et al. Hypoxic repression of pyruvate dehydrogenase activity is necessary for metabolic reprogramming and growth of model tumours. Sci. Rep. 2016, 6, 31146. [Google Scholar] [CrossRef]
- Hoole, S.; Jaworski, C.; Brown, A.; Mccormick, L.; Agrawal, B.; Clarke, S.; West, N.E.J. Serial assessment of the index of microcirculatory resistance during primary percutaneous coronary intervention comparing manual aspiration catheter thrombectomy with balloon angioplasty (IMPACT study): A randomised controlled pilot study. Open Heart 2015, 2, e000238. [Google Scholar] [CrossRef]
- Chen, Q.; Vazquez, E.J.; Moghaddas, S.; Hoppel, C.L.; Lesnefsky, E.J. Production of reactive oxygen species by mitochondria: Central role of complex III. J. Biol. Chem. 2003, 278, 36027–36031. [Google Scholar] [CrossRef]
- Yan, S.; Du, F.; Wu, L.; Zhang, Z.; Zhong, C.; Yu, Q.; Wang, Y.; Lue, L.-F.; Walker, D.G.; Douglas, J.T.; et al. F1F0 ATP synthase–cyclophilin D interaction contributes to diabetes-induced synaptic dysfunction and cognitive decline. Diabetes 2016, 65, 3482–3494. [Google Scholar] [CrossRef]
- Brito, M.D.; Siena, A.; Chipara, M.; Sarkar, S.; Rosenstock, T.R. Oxygen Consumption Evaluation: An important indicator of metabolic state, cellular function, and cell fate along neural deregulation. In Toxicity Assessment; Springer: Berlin/Heidelberg, Germany, 2021; pp. 207–230. [Google Scholar]
- Liu, Y.J.; Wang, D.Y.; Yang, Y.J.; Lei, W.-F. Effects and mechanism of dexmedetomidine on neuronal cell injury induced by hypoxia-ischemia. BMC Anesthesiol. 2017, 17, 117. [Google Scholar] [CrossRef] [PubMed]
- Nicotera, P.; Bano, D.J.C. The enemy at the gates. Ca2+ entry through TRPM7 channels and anoxic neuronal death. Cell 2004, 115, 768–770. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Li, S.; Wang, Y.; Yan, Z.; Guo, Y.; Zhang, L. A Novel 5-Chloro-N-phenyl-1H-indole-2-carboxamide Derivative as Brain-Type Glycogen Phosphorylase Inhibitor: Potential Therapeutic Effect on Cerebral Ischemia. Molecules 2022, 27, 6333. https://doi.org/10.3390/molecules27196333
Huang Y, Li S, Wang Y, Yan Z, Guo Y, Zhang L. A Novel 5-Chloro-N-phenyl-1H-indole-2-carboxamide Derivative as Brain-Type Glycogen Phosphorylase Inhibitor: Potential Therapeutic Effect on Cerebral Ischemia. Molecules. 2022; 27(19):6333. https://doi.org/10.3390/molecules27196333
Chicago/Turabian StyleHuang, Yatao, Shuai Li, Youde Wang, Zhiwei Yan, Yachun Guo, and Liying Zhang. 2022. "A Novel 5-Chloro-N-phenyl-1H-indole-2-carboxamide Derivative as Brain-Type Glycogen Phosphorylase Inhibitor: Potential Therapeutic Effect on Cerebral Ischemia" Molecules 27, no. 19: 6333. https://doi.org/10.3390/molecules27196333
APA StyleHuang, Y., Li, S., Wang, Y., Yan, Z., Guo, Y., & Zhang, L. (2022). A Novel 5-Chloro-N-phenyl-1H-indole-2-carboxamide Derivative as Brain-Type Glycogen Phosphorylase Inhibitor: Potential Therapeutic Effect on Cerebral Ischemia. Molecules, 27(19), 6333. https://doi.org/10.3390/molecules27196333