
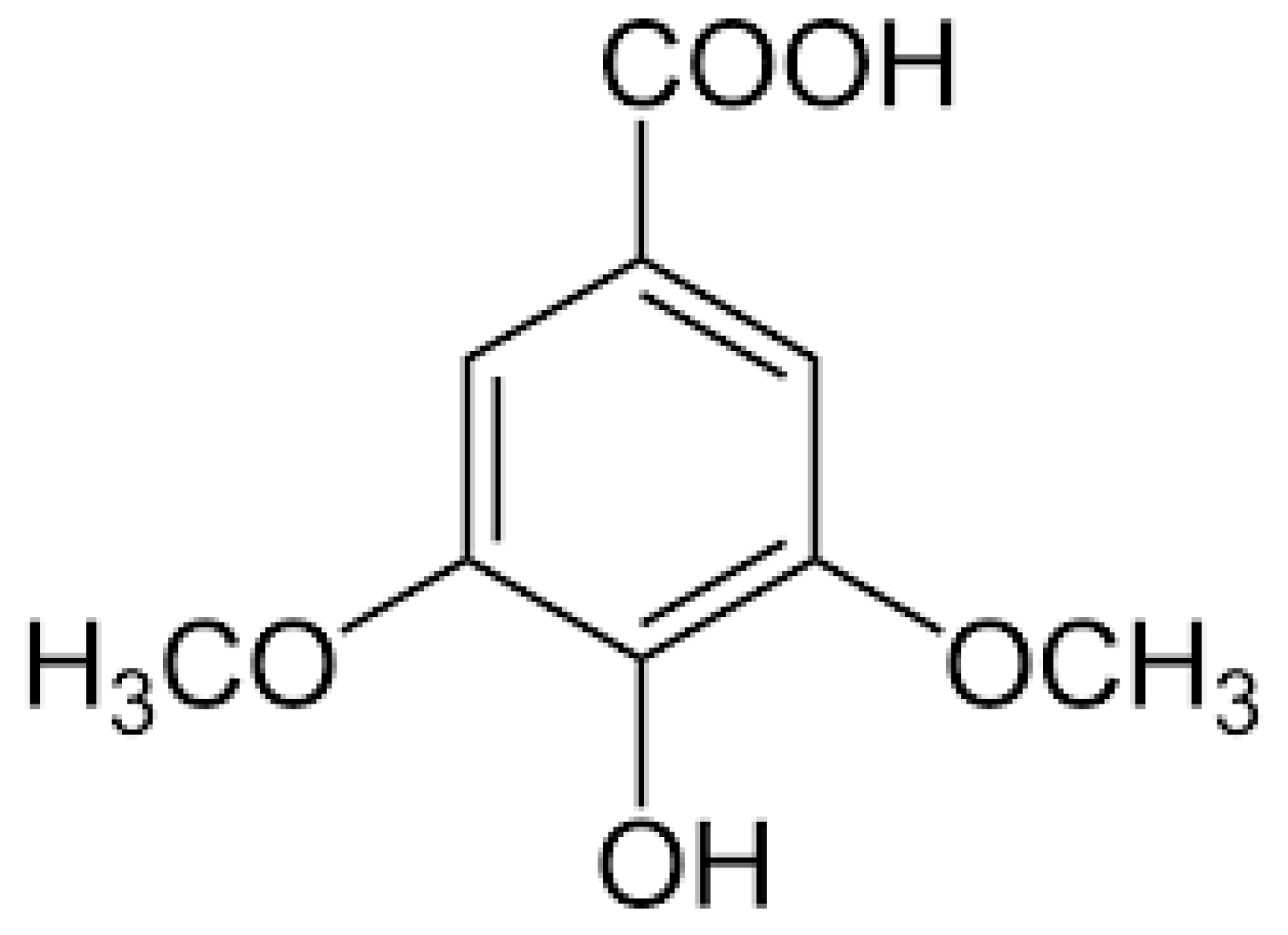
Syringic Acid Ameliorates Cardiac, Hepatic, Renal and Neuronal Damage Induced by Chronic Hyperglycaemia in Wistar Rats: A Behavioural, Biochemical and Histological Analysis
Abstract
:1. Introduction
2. Material and Methods
2.1. Agents and Chemicals
2.2. Animals and Induction of T2DM-Associated Complications
2.3. Experimental Design and Protocol
2.4. Sample Collection and Tissue Preparation
2.5. Estimation of Biochemical and Behavioural Parameters
2.6. Body Weight, Urine Output, and Food and Water Intake
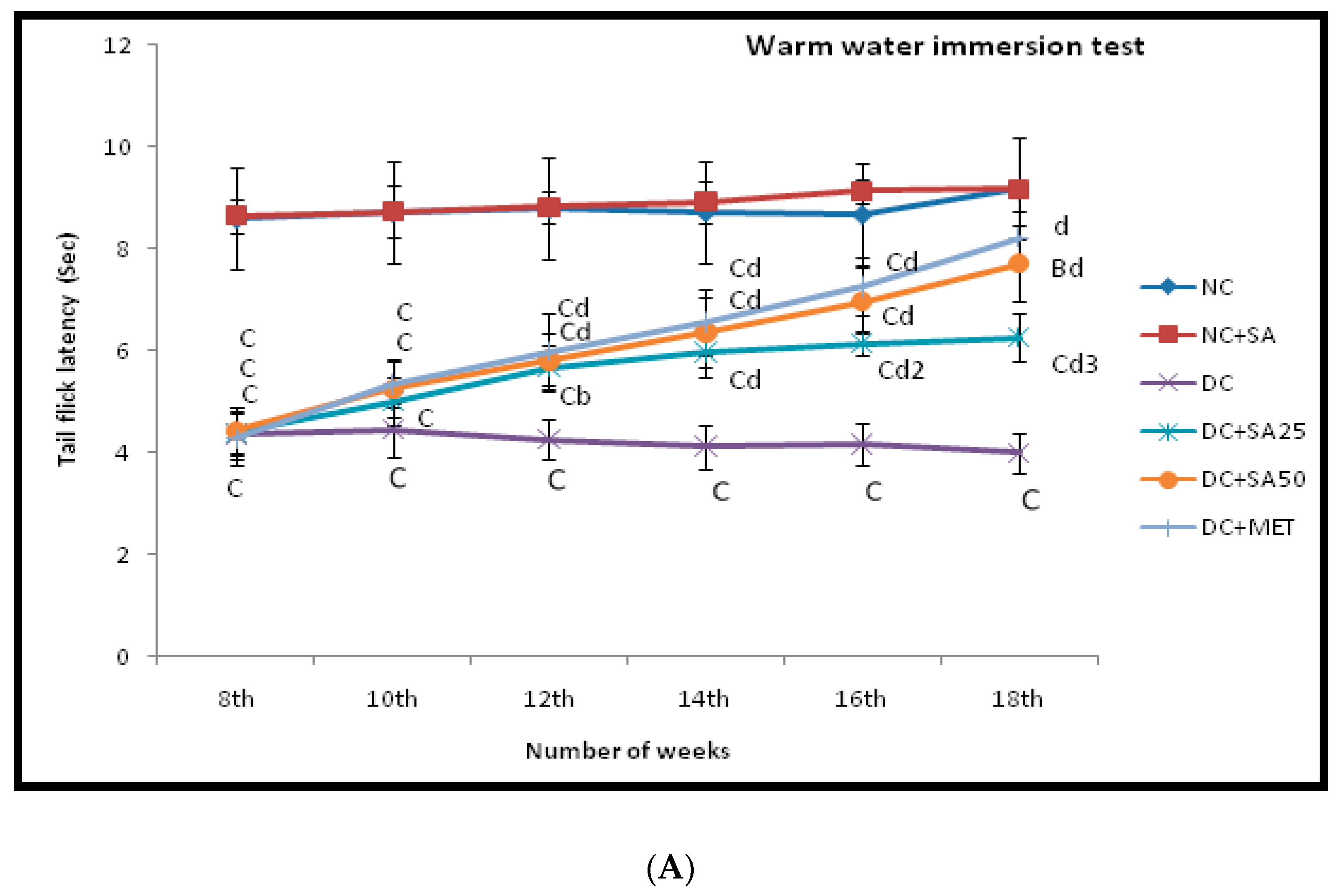
2.7. Warm and Cold Water Tail-Immersion Test
2.8. Measurement of Blood and Urine Biochemical Parameters
2.9. Measurement of Physical Parameters
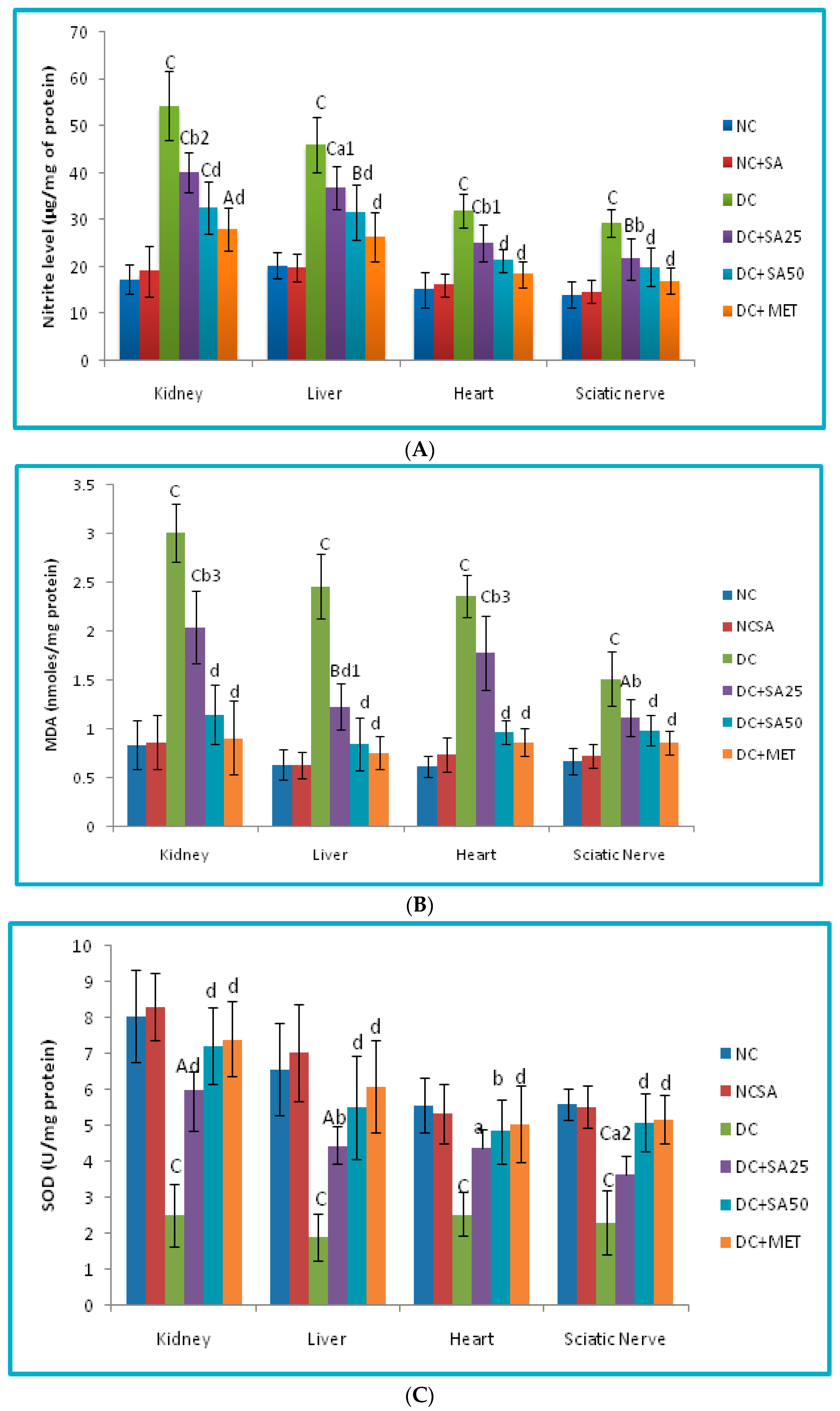
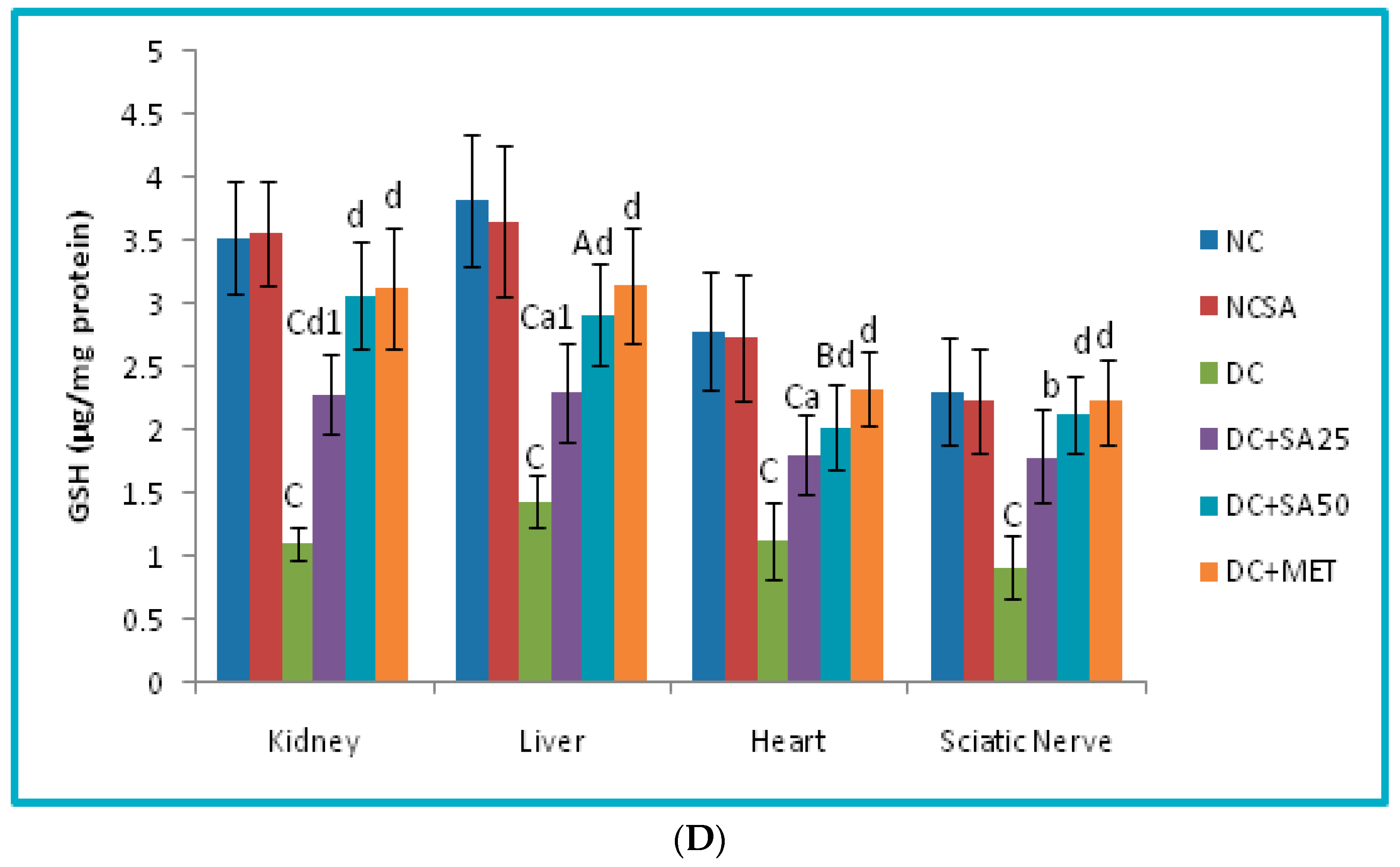
2.10. Evaluation of Oxidative Stress Level in Renal, Cardiac, Hepatic and Neuronal Tissues
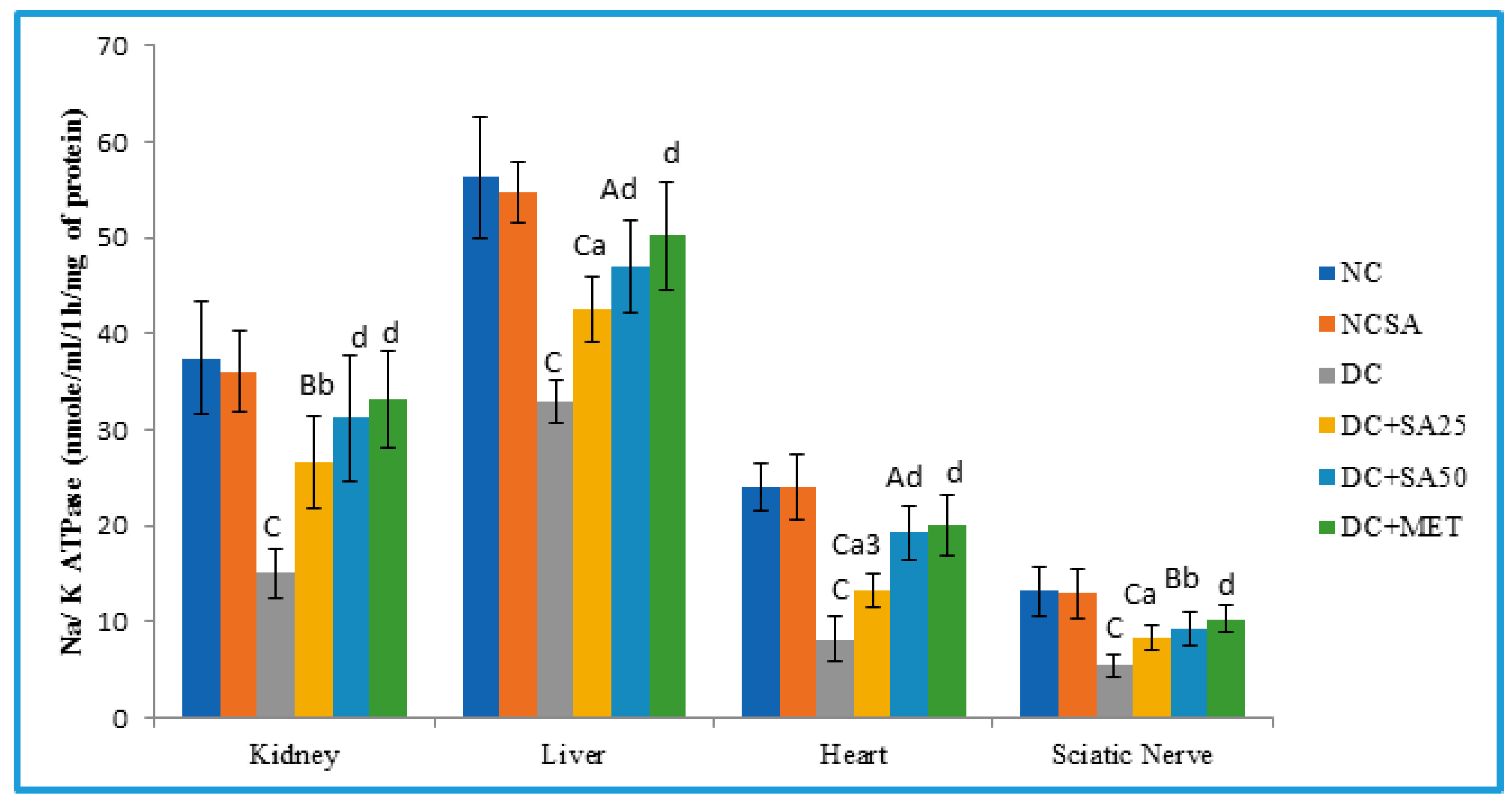
2.11. Determination of Membrane-Bound Inorganic Phosphate Determination (Na-K ATPase)
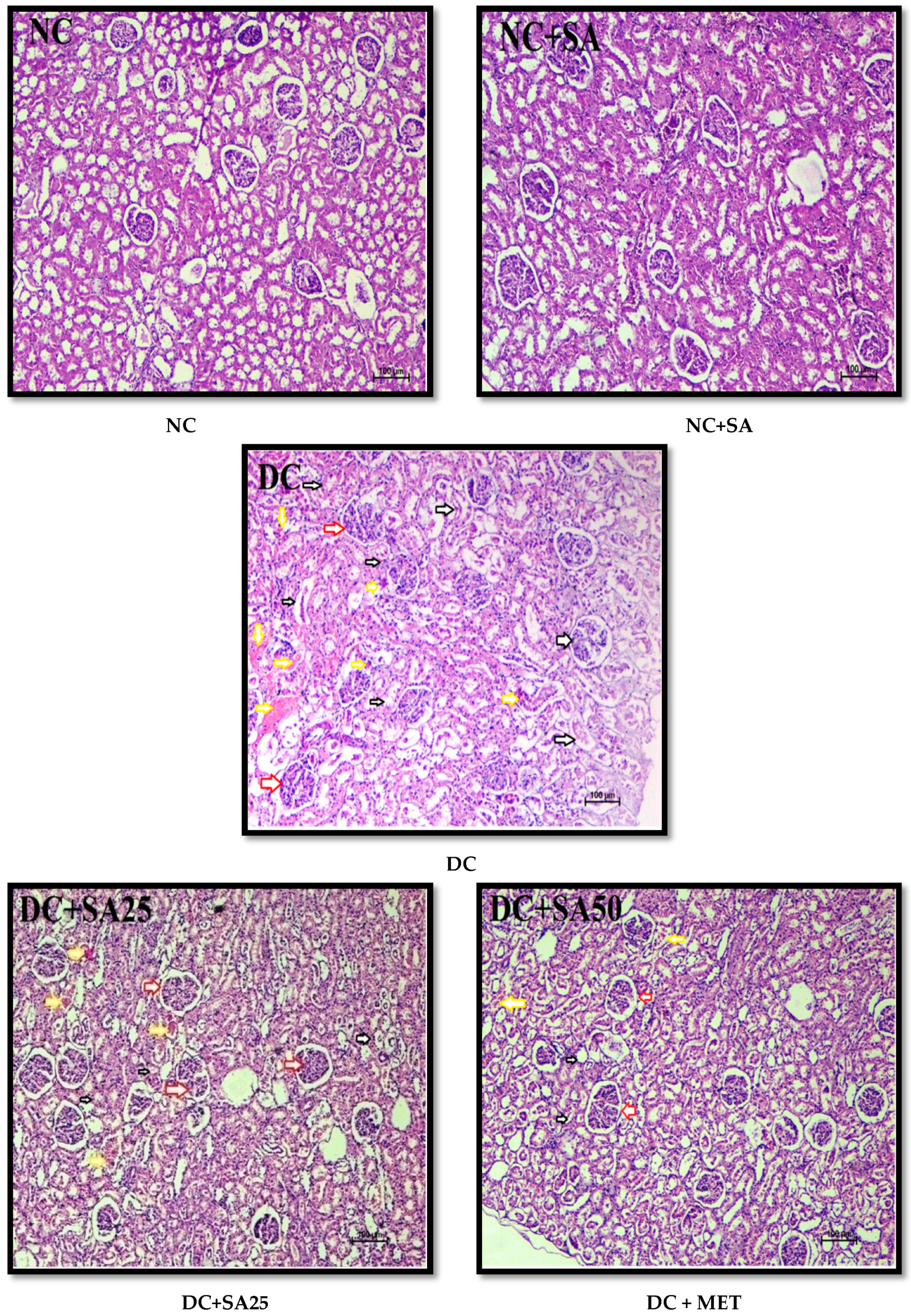
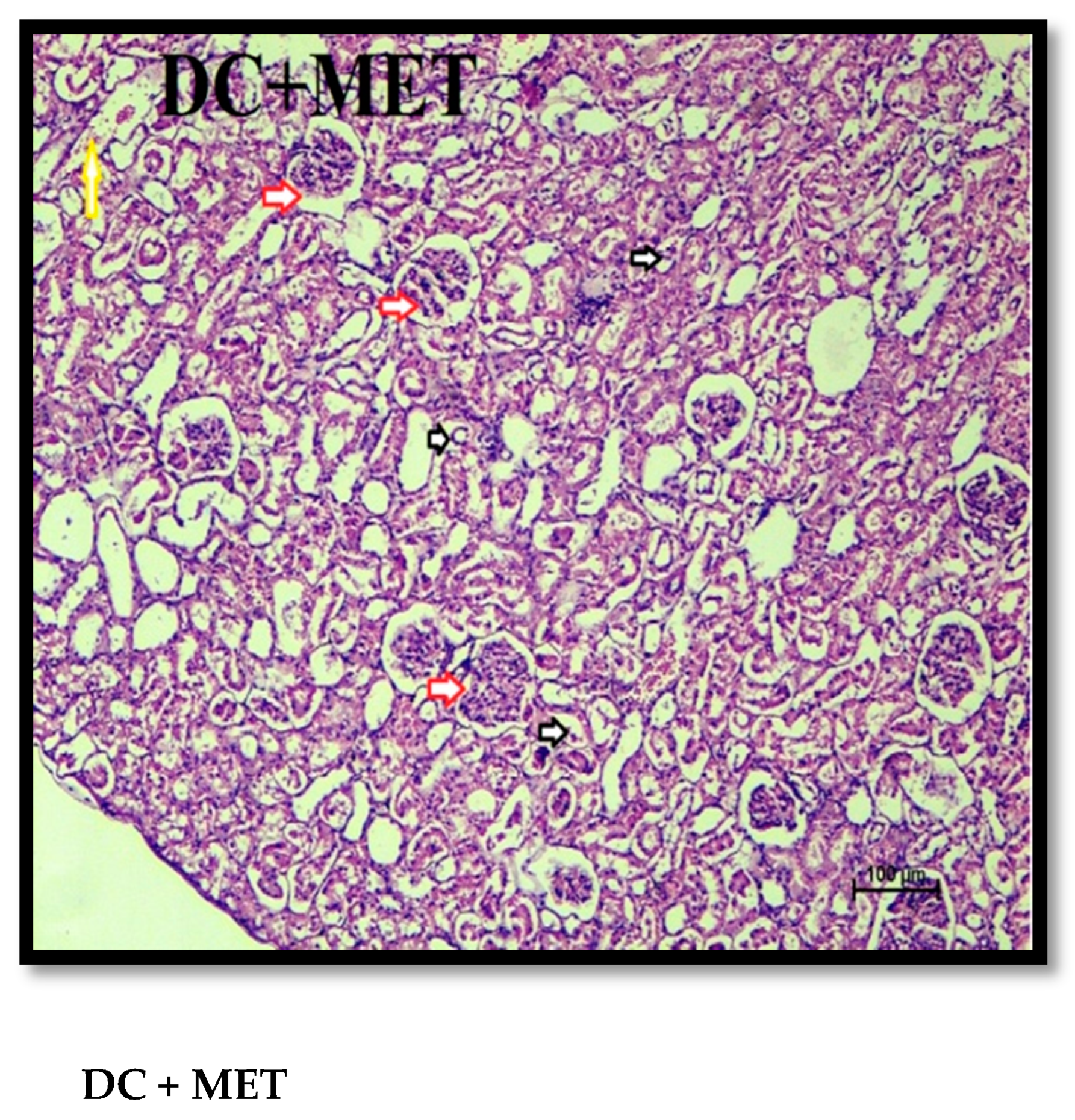
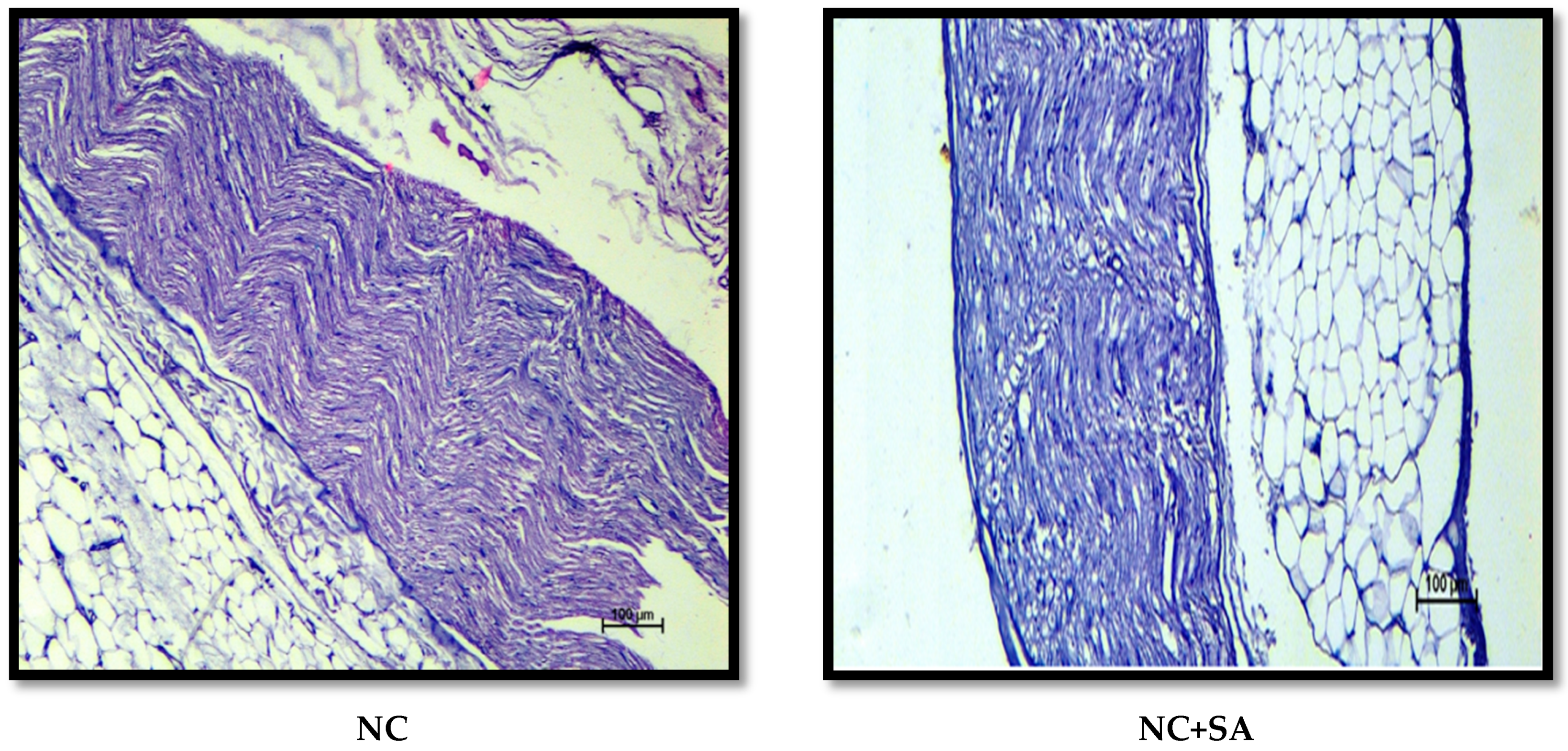
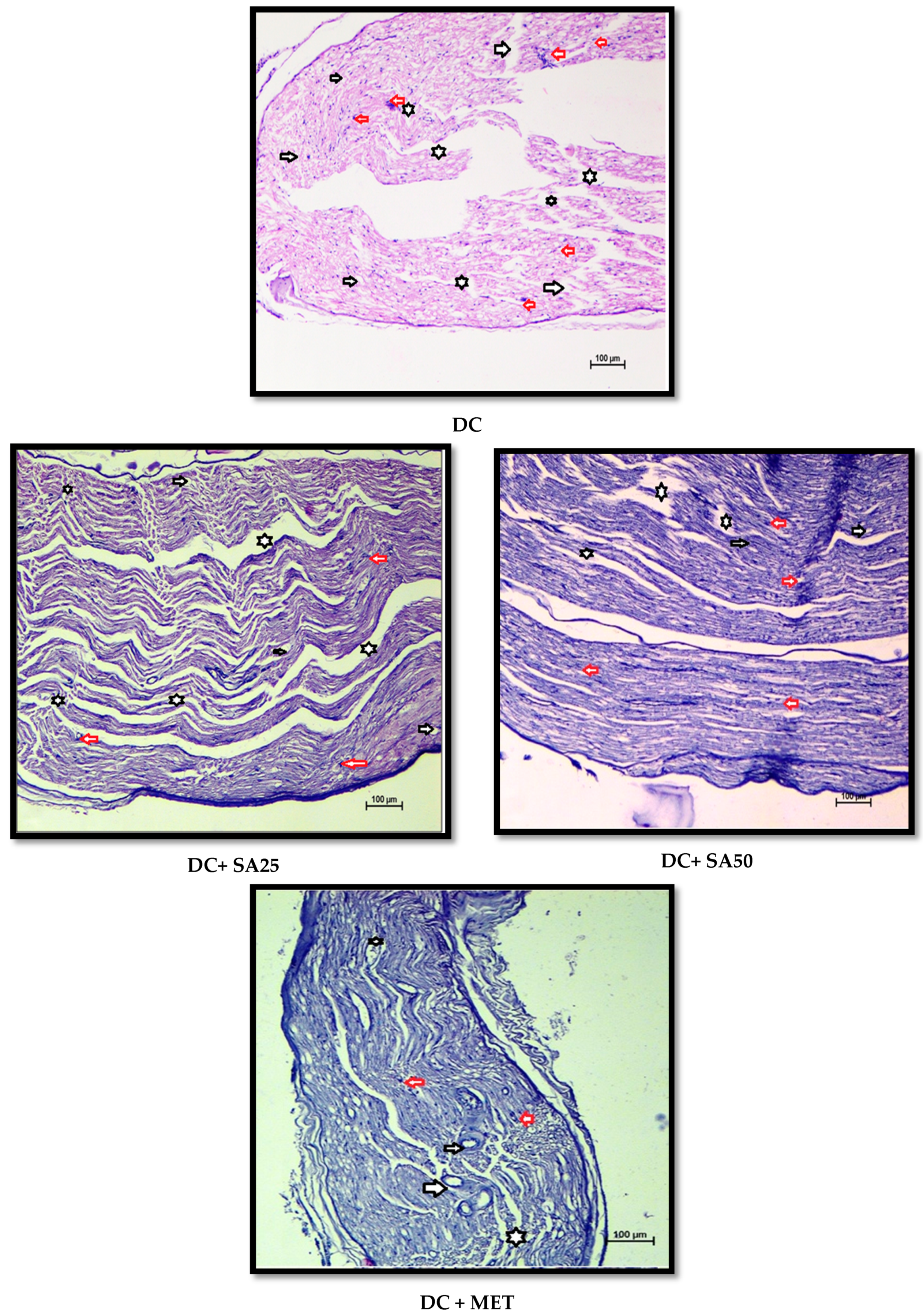
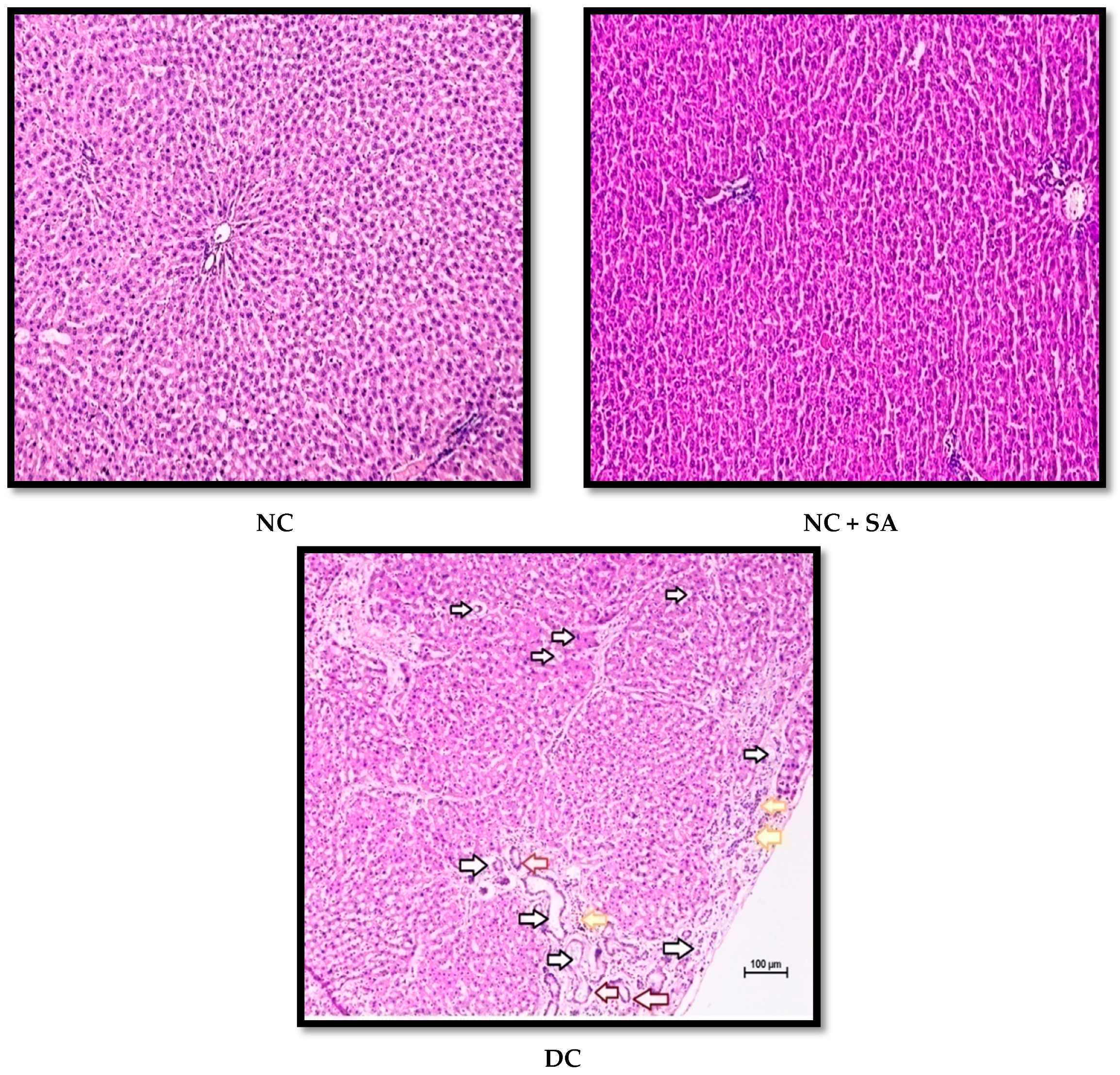
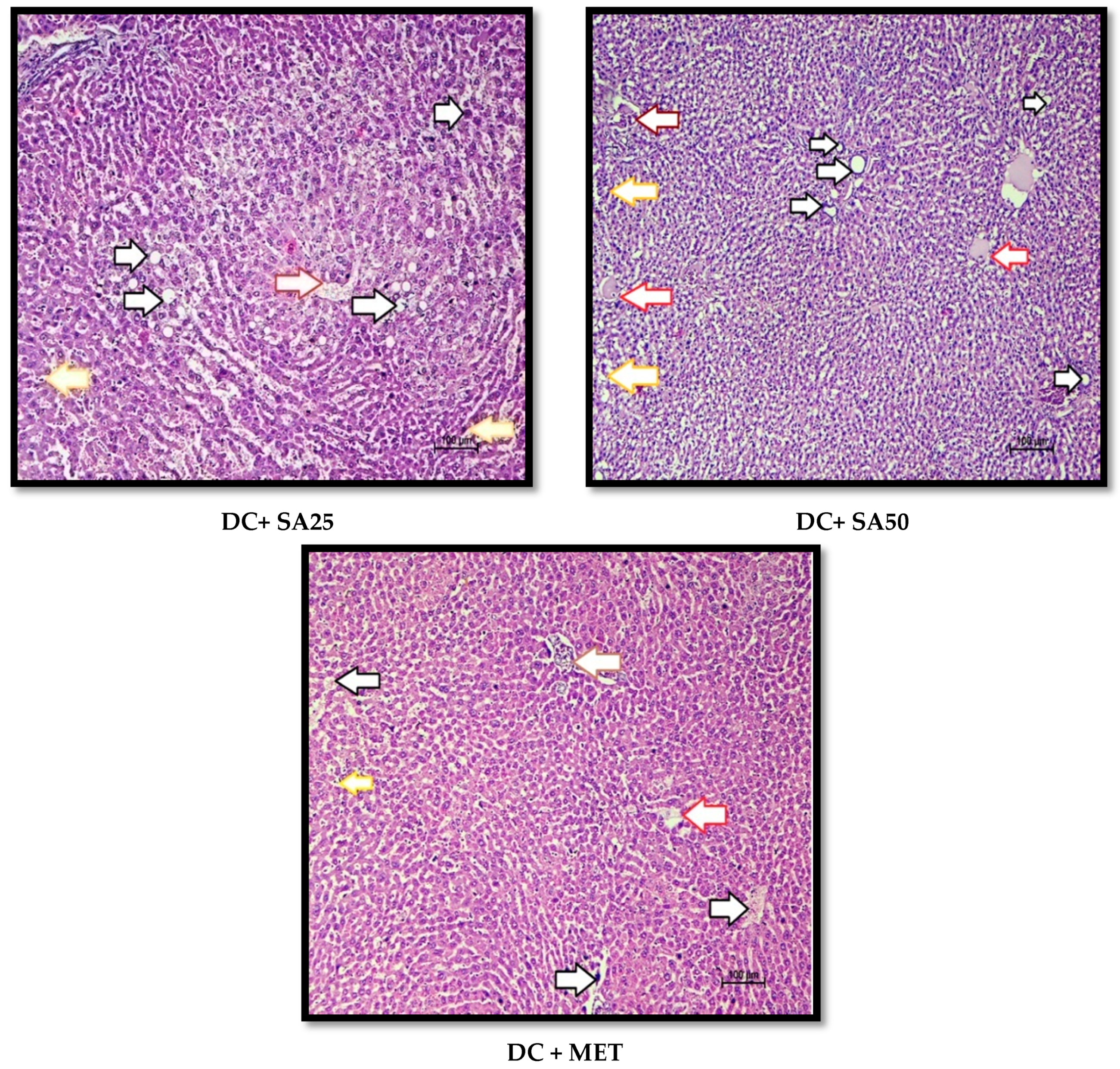
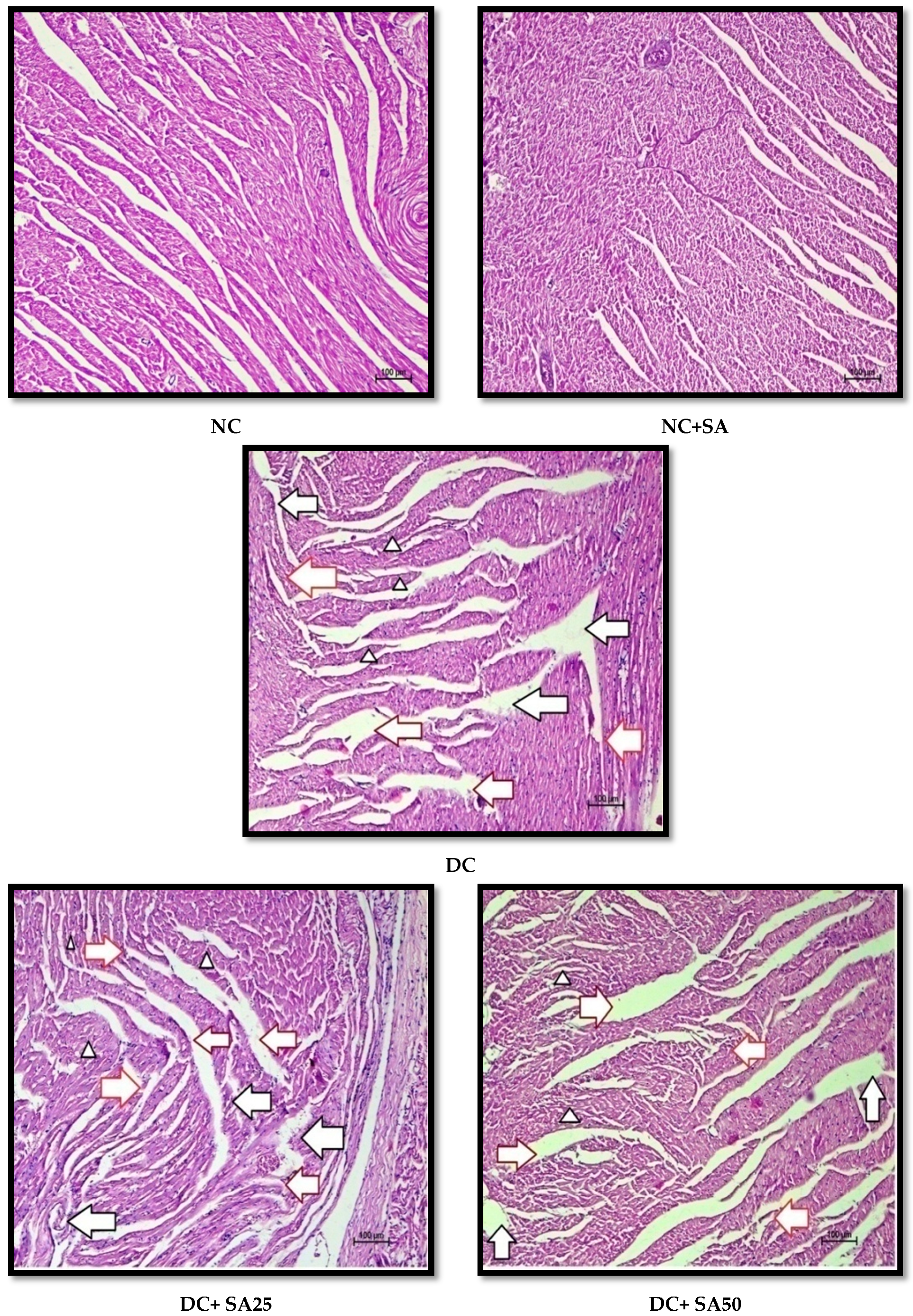
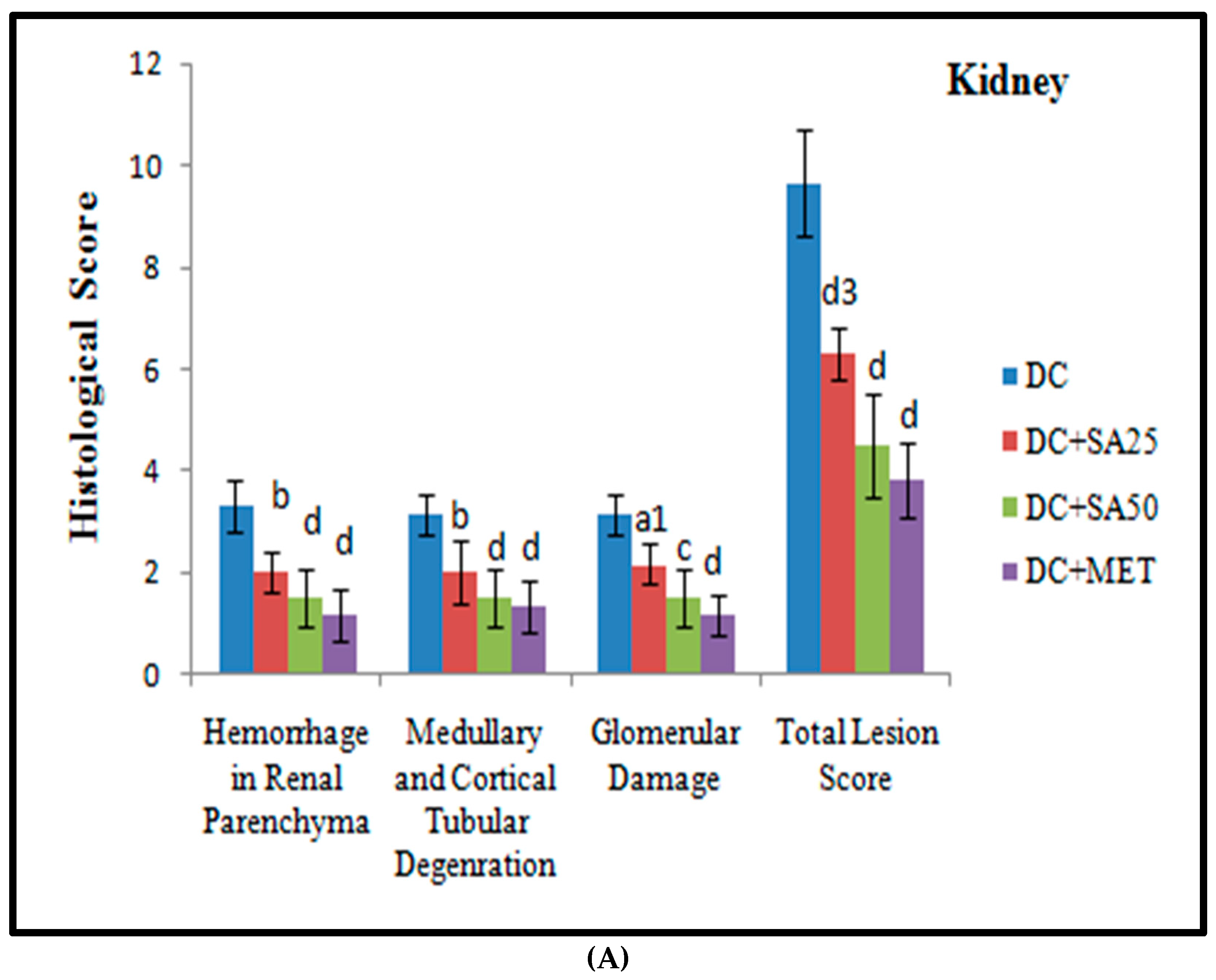
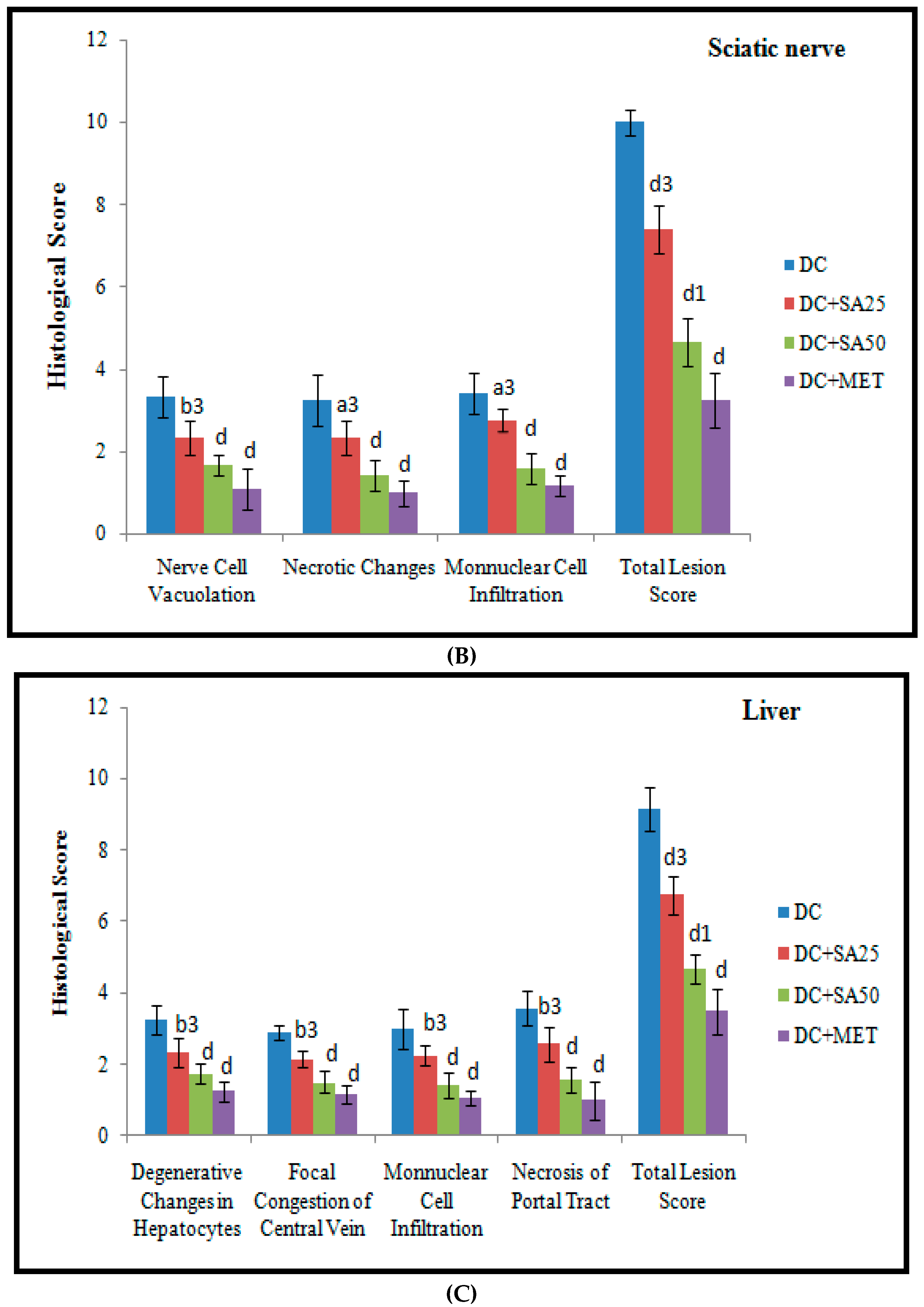
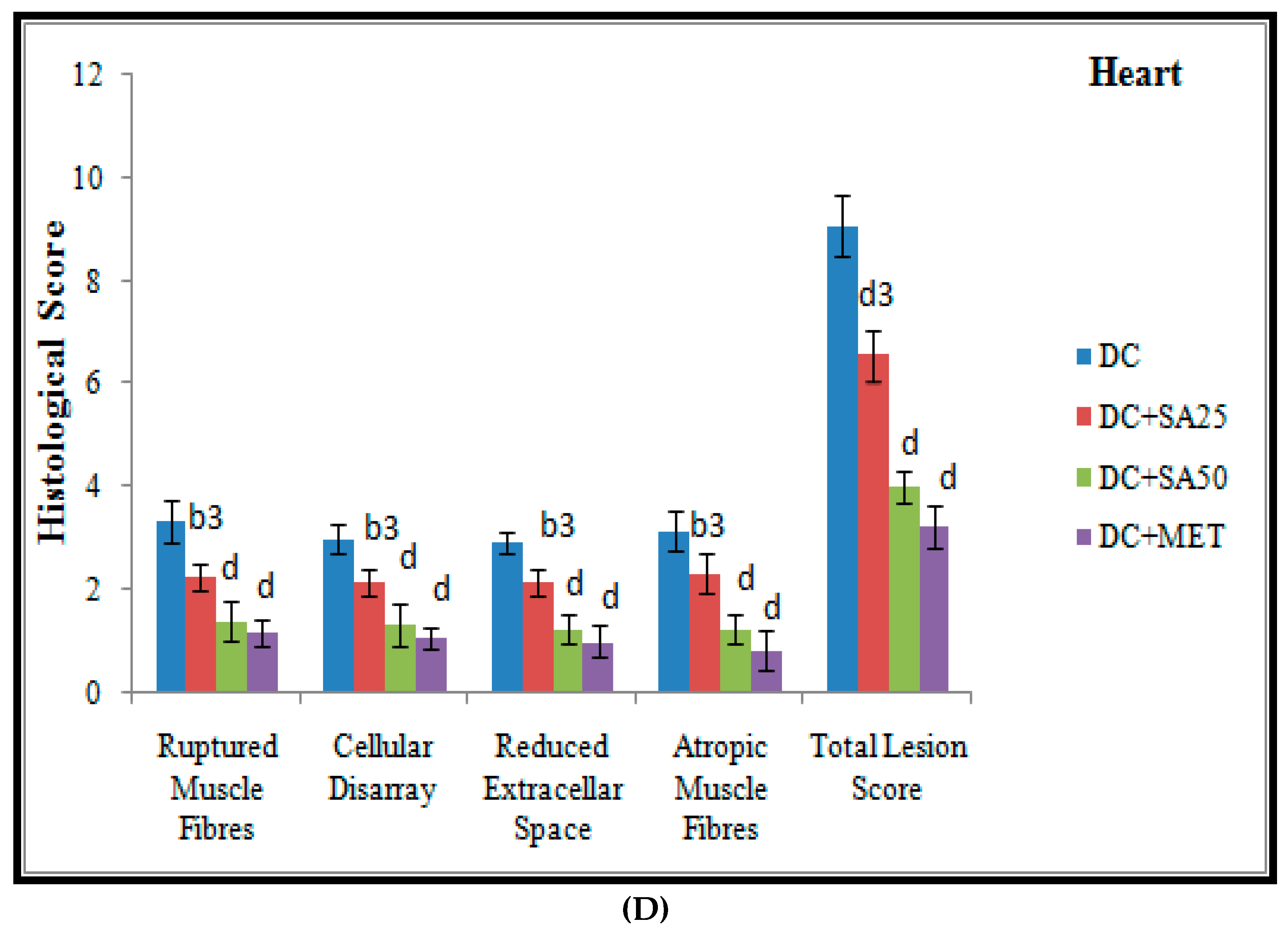
2.12. Histopathological Examination
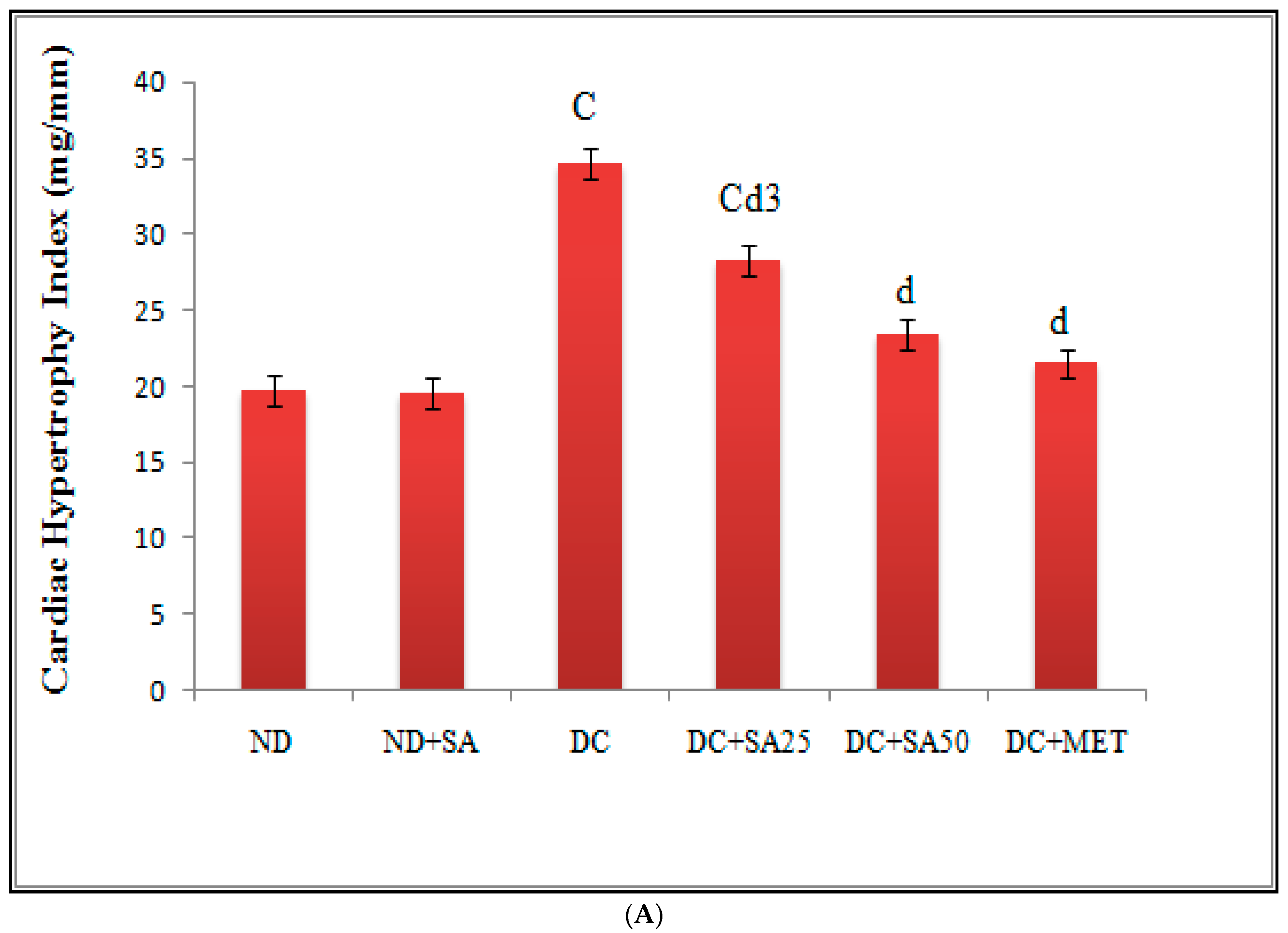
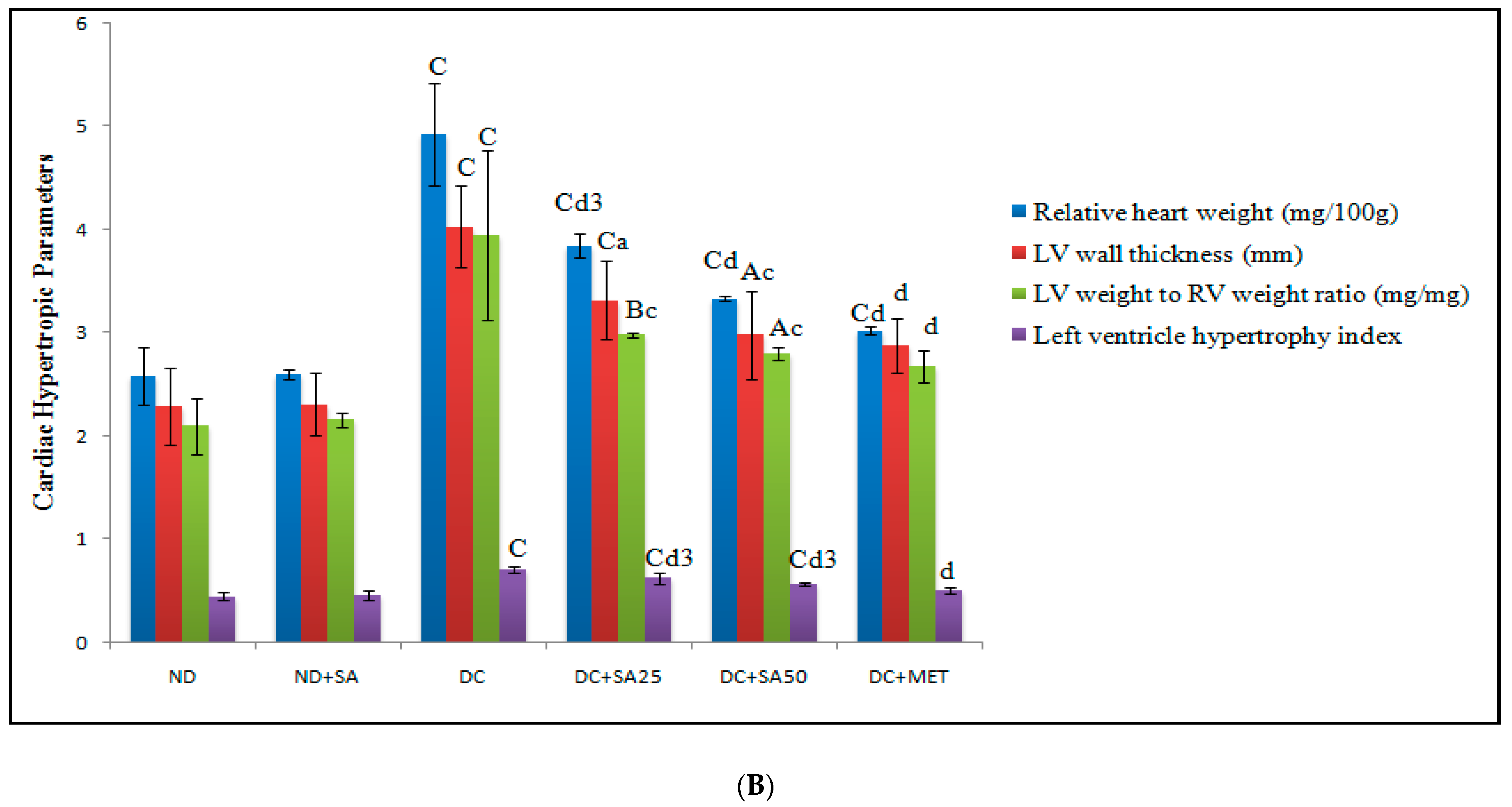
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation: Fact Figures. 2022. Available online: https://idf.org/aboutdiabetes/what-is-diabetes/facts-figures.html (accessed on 5 March 2022).
- Laakso, M. Heart in diabetes: A microvascular disease. Diabetes Care 2011, 34 (Suppl. 2), S145–S149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Okawa, H.; Doi, K. Neoplastic lesions in streptozotocin-treated rats. Exp. Anim. 1983, 32, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Ghule, A.E.; Jadhav, S.S.; Bodhankar, S.L. Trigonelline ameliorates diabetic hypertensive nephropathy by suppression of oxidative stress in kidney and reduction in renal cell apoptosis and fibrosis in streptozotocin induced neonatal diabetic (nSTZ) rats. Int. Immunopharmacol. 2012, 14, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Shang, G.; Gao, P.; Zhao, Z.; Chen, Q.; Jiang, T.; Zhang, N.; Li, H. 3,5-Diiodo-l-thyronine ameliorates diabetic nephropathy in streptozotocin-induced diabetic rats. Biochim. Biophys. Acta-Mol. Basis Dis. 2013, 1832, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Albers, J.W.; Pop-Busui, R. Diabetic neuropathy: Mechanisms, emerging treatments, and subtypes. Curr. Neurol. Neurosci. Rep. 2014, 14, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herlitz, J.; Malmberg, K.; Karlson, B.W.; Rydén, L.; Hjalmarson, Å. Mortality and morbidity during a five-year follow-up of diabetics with myocardial infarction. Acta Neurol. Scand. 1988, 224, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.A.; Patel, B.; Khattar, R.S.; Malik, R.A. Diabetic cardiomyopathy: Mechanisms, diagnosis and treatment. Clin. Sci. 2004, 107, 539–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, K.; Das, J.; Sil, P.C. Taurine ameliorate alloxan induced oxidative stress and intrinsic apoptotic pathway in the hepatic tissue of diabetic rats. Food Chem. Toxicol. 2013, 51, 317–329. [Google Scholar] [CrossRef]
- Harrison, S.A. Liver Disease in Patients with Diabetes Mellitus. J. Clin. Gastroenterol. 2006, 40, 68–76. [Google Scholar] [CrossRef]
- Tolman, K.G.; Fonseca, V.; Tan, M.H.; Dalpiaz, A. Narrative Review: Hepatobiliary Disease in Type 2 Diabetes Mellitus. Ann. Intern. Med. 2004, 141, 946. [Google Scholar] [CrossRef] [PubMed]
- Ling, W.; Huang, Y.; Huang, Y.M.; Fan, R.R.; Sui, Y.; Zhao, H.L. Global trend of diabetes mortality attributed to vascular complications, 2000-2016. Cardiovasc. Diabetol. 2020, 19, 182. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.Y.; Qian, K.; Morris-Natschke, S.L.; Hsu, C.S.; Lee, K.H. Recent Discovery of Plant-Derived Anti-Diabetic Natural Products. Nat. Prod. Rep. 2012, 29, 580. [Google Scholar] [CrossRef]
- Inzucchi, S.E. Oral Antihyperglycemic Therapy for Type 2 Diabetes. JAMA 2002, 287, 360. [Google Scholar] [CrossRef] [Green Version]
- Kahn, S.E.; Haffner, S.M.; Heise, M.A.; Herman, W.H.; Holman, R.R.; Jones, N.P.; Kravitz, B.G.; Lachin, J.M.; O’Neill, M.C.; Zinman, B.; et al. Glycemic Durability of Rosiglitazone, Metformin, or Glyburide Monotherapy. N. Engl. J. Med. 2006, 355, 2427–2443. [Google Scholar] [CrossRef] [Green Version]
- Saxena, A.; Vikram, N.K. Role of Selected Indian Plants in Management of Type 2 Diabetes: A Review. J. Altern. Complement. Med. 2004, 10, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Suresh Kumar, C. Syringic acid (SA)–A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Muthukumaran, J.; Srinivasan, S.; Venkatesan, R.S.; Ramachandran, V.; Muruganathan, U. Syringic acid, a novel natural phenolic acid, normalizes hyperglycemia with special reference to glycoprotein components in experimental diabetic rats. J. Acute Dis. 2013, 2, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Muthukumaran, J.; Muruganathan, U.; Venkatesan, R.S.; Jalaludeen, A.M. Antihyperglycemic effect of syringic acid on attenuating the key enzymes of carbohydrate metabolism in experimental diabetic rats. Biomed. Prev. Nutr. 2014, 4, 595–602. [Google Scholar] [CrossRef]
- Pawar, S.; Upaganlawar, A.; Upasani, C. Evaluation of Some Phenolic Acids in Diabetic Neuropathy. Indian J. Pharm. Educ. Res. 2021, 55, S176–S183. [Google Scholar] [CrossRef]
- Bhattacherjee, A.; Datta, A. Mechanism of antiglycating properties of syringic and chlorogenic acids in in vitro glycation system. Int. Food Res. J. 2015, 77, 540–548. [Google Scholar] [CrossRef]
- Ham, J.R.; Lee, H.I.; Choi, R.Y.; Sim, M.O.; Seo, K.I.; Lee, M.K. Anti-steatotic and anti-inflammatory roles of syringic acid in high-fat diet-induced obese mice. Food Funct. 2016, 7, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, L.; Wang, X.; Wu, W.; Qin, R. Effect of Syringic acid on antioxidant biomarkers and associated inflammatory markers in mice model of asthma. Drug Dev Res. 2019, 80, 253–261. [Google Scholar] [CrossRef]
- Fernandez, M.A.; Saenz, M.T.; Garcia, M.D. Anti-inflammatory activity in rats and mice of phenolic acids isolated from Scrophularia frutescens. J. Pharm. Pharmacol. 1998, 50, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Prahalathan, P.; Raja, B. Syringic acid ameliorates (L)-NAME-induced hypertension by reducing oxidative stress. Naunyn. Schmiedebergs Arch. Pharmacol. 2012, 385, 1175–1184. [Google Scholar] [CrossRef]
- Cikman, O.; Soylemez, O.; Ozkan, O.F.; Kiraz, H.A.; Sayar, I.; Ademoglu, S.; Taysi, S.; Karaayvaz, M. Antioxidant Activity of Syringic Acid Prevents Oxidative Stress in l-arginine-Induced Acute Pancreatitis: An Experimental Study on Rats. Int. Surg. 2015, 100, 891–896. [Google Scholar] [CrossRef] [Green Version]
- Aziz, N.H.; Farag, S.E.; Mousa, L.A.; Abo-Zaid, M.A. Comparative antibacterial and antifungal effects of some phenolic compounds. Microbios 1998, 93, 43–54. [Google Scholar]
- Shi, C.; Sun, Y.; Zheng, Z.; Zhang, X.; Song, K.; Jia, Z.; Chen, Y.; Yang, M.; Liu, X.; Dong, R.; et al. Antimicrobial activity of syringic acid against Cronobacter sakazakii and its effect on cell membrane. Food Chem. 2016, 197 Pt A, 100–106. [Google Scholar] [CrossRef]
- Tokmak, M.; Yuksel, Y.; Sehitoglu, M.H.; Guven, M.; Akman, T.; Aras, A.B.; Cosar, M.; Abbed, K.M. The Neuroprotective Effect of Syringic Acid on Spinal Cord Ischemia/Reperfusion Injury in Rats. Inflammation 2015, 38, 1969–1978. [Google Scholar] [CrossRef]
- Tokmak, M.; Sehitoglu, M.H.; Yuksel, Y.; Guven, M.; Akman, T.; Aras, A.B.; Yaka, U.; Gomleksiz, C.; Albayrak, S.B.; Cosar, M. The Axon Protective Effects of Syringic Acid on Ischemia/Reperfusion Injury in a Rat Sciatic Nerve Model. Turk Neurosurg. 2017, 27, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Güven, M.; Aras, A.B.; Topaloğlu, N.; Özkan, A.; Şen, H.M.; Kalkan, Y.; Okuyucu, A.; Akbal, A.; Gökmen, F.; Coşar, M. The protective effect of syringic acid on ischemia injury in rat brain. Turk J. Med. Sci. 2015, 45, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Rashedinia, M.; Alimohammadi, M.; Shalfroushan, N.; Khoshnoud, M.J.; Mansourian, M.; Azarpira, N.; Sabahi, Z. Neuroprotective Effect of Syringic Acid by Modulation of Oxidative Stress and Mitochondrial Mass in Diabetic Rats. Biomed. Res. Int. 2020, 2020, 8297984. [Google Scholar] [CrossRef] [PubMed]
- Dalmagro, A.P.; Camargo, A.; Zeni, A.L.B. Morus nigra and its major phenolic, syringic acid, have antidepressant-like and neuroprotective effects in mice. Metab. Brain Dis. 2017, 32, 1963–1973. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, L.; Sun, S.; Yi, Z.; Jiang, X.; Jia, D. Neuroprotective effects of syringic acid against OGD/R-induced injury in cultured hippocampal neuronal cells. Int. J. Mol. Med. 2016, 38, 567–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Dang, M.; Zhang, W.; Lei, Y.; Ramesh, T.; Veeraraghavan, V.P.; Hou, X. Neuroprotective effects of Syringic acid against aluminium chloride induced oxidative stress mediated neuroinflammation in rat model of Alzheimer’s disease. J. Funct. Foods. 2020, 71, 104009. [Google Scholar] [CrossRef]
- Ramachandran, V.; Raja, B. Protective effects of syringic acid against acetaminophen-induced hepatic damage in albino rats. J. Basic Clin. Physiol. Pharmacol. 2010, 21, 369–385. [Google Scholar] [CrossRef]
- Itoh, A.; Isoda, K.; Kondoh, M.; Kawase, M.; Watari, A.; Kobayashi, M.; Tamesada, M.; Yagi, K. Hepatoprotective effect of syringic acid and vanillic acid on CCl4-induced liver injury. Biol. Pharm Bull. 2010, 33, 983–987. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Chen, D.; Yi, Y.; Qi, H.; Gao, X.; Fang, H.; Gu, Q.; Wang, L.; Gu, L. Syringic Acid Extracted from Herba dendrobii Prevents Diabetic Cataract Pathogenesis by Inhibiting Aldose Reductase Activity. Evid.-Based Complement. Altern. Med. 2012, 2012, 426537. [Google Scholar] [CrossRef] [Green Version]
- Orabi, K.Y.; Abaza, M.S.; El Sayed, K.A.; Elnagar, A.Y.; Al-Attiyah, R.; Guleri, R.P. Selective growth inhibition of human malignant melanoma cells by syringic acid-derived proteasome inhibitors. Cancer Cell Int. 2013, 13, 82. [Google Scholar] [CrossRef] [Green Version]
- Abaza, M.S.; Al-Attiyah, R.; Bhardwaj, R.; Abbadi, G.; Koyippally, M.; Afzal, M. Syringic acid from Tamarix aucheriana possesses antimitogenic and chemo-sensitizing activities in human colorectal cancer cells. Pharm Biol. 2013, 51, 1110–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauer, G.M.; Stansbury, J.W. Cements containing syringic acid esters-o-ethoxybenzoic acid and zinc oxide. J. Dent. Res. 1984, 63, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Kim, S. Mechanisms of attenuation of clot formation and acute thromboembolism by syringic acid in mice. J. Funct. Foods 2018, 43, 112–122. [Google Scholar] [CrossRef]
- Liszt, K.I.; Eder, R.; Wendelin, S.; Somoza, V. Identification of Catechin, Syringic Acid, and Procyanidin B2 in Wine as Stimulants of Gastric Acid Secretion. J. Agric. Food Chem. 2015, 63, 7775–7783. [Google Scholar] [CrossRef] [PubMed]
- Rasheeda, K.; Bharathy, H.; Nishad Fathima, N. Vanillic acid and syringic acid: Exceptionally robust aromatic moieties for inhibiting in vitro self-assembly of type I collagen. Int. J. Biol. Macromol. 2018, 113, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.C.; Panchal, S.S. Safety evaluation of syringic acid: Subacute oral toxicity studies in Wistar rats. Heliyon 2019, 5, e02129. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.A.; Panchal, S.S. Streptozotocin-Induced Diabetes Mellitus in Neonatal Rats: An Insight into its Applications to Induce Diabetic Complications. Curr. Diabetes Rev. 2019, 16, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Bonner-Weir, S.; Trent, D.F.; Honey, R.N.; Weir, G.C. Responses of neonatal rat islets to streptozotocin: Limited B-cell regeneration and hyperglycemia. Diabetes 1981, 30, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Marathe, P.A.; Parekar, R.R.; Shinde, S.P.; Rege, N.N. A split dose regimen of streptozotocin to induce diabetes in a neonatal rat model. Indian, J. Pharmacol. 2006, 38, 432. [Google Scholar] [CrossRef]
- Salman, I.M.; Inamdar, M.N. Effect of gliclazide on cardiovascular risk factors involved in split-dose streptozotocin induced neonatal rat model: A chronic study. Int. J. Basic Clin. Pharmacol. 2012, 1, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, C.P.; Bodhankar, S.L.; Ghule, A.E.; Mohan, V.; Thakurdesai, P.A. Antidiabetic activity of Trigonella foenumgraecum L. seeds extract (IND01) in neonatal streptozotocin-induced (n-STZ) rats. Diabetol. Croat. 2012, 41, 29–40. [Google Scholar]
- Sharma, R.; Kumar, A.; Srinivasan, B.P.; Chauhan, A.; Dubey, K. Cardioprotective effects of Ficus religiosa in neonatal streptozotocin-induced diabetic cardiomyopathy in rats. Biomed. Aging Pathol. 2014, 4, 53–58. [Google Scholar] [CrossRef]
- Kandhare, A.D.; Raygude, K.S.; Ghosh, P.; Ghule, A.E.; Bodhankar, S.L. Neuroprotective effect of naringin by modulation of endogenous biomarkers in streptozotocin induced painful diabetic neuropathy. Fitoterapia. 2012, 83, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.M.; Bhadada, S.V. Type 2 diabetes-induced cardiovascular complications: Comparative evaluation of spironolactone, atenolol, metoprolol, ramipril and perindopril. Clin. Exp. Hypertens. 2014, 36, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Sabahi, Z.; Khoshnoud, M.J.; Khalvati, B.; Hashemi, S.S.; Farsani, Z.G.; Gerashi, H.M.; Rashedinia, M. Syringic acid improves oxidative stress and mitochondrial biogenesis in the liver of streptozotocin-induced diabetic rats. Asian Pac. J. Trop. Biomed. 2020, 10, 111. [Google Scholar]
- Sammeturi, M.; Shaik, A.H.; Prasad, E.M.; Mohammad, A.; Kodidhela, L.D. Cardioprotective molecular mechanism of syringic acid against isoproterenol induced post-myocardial toxicity in male albino wistar rats. J. King Saud Univ. Sci. 2020, 32, 1375–1381. [Google Scholar] [CrossRef]
- Samatha, P.; Venkateswarlu, M.; Prabodh, S. Role of biochemical markers in the prediction of microvascular complications in type-2 diabetes mellitus. J. Clin. Diagnostic Res. 2011, 5, 1154–1157. [Google Scholar]
- Singh, M.; Anwer, E.; Kumar, V. Assessment of biochemical parameters in the patients of coronary artery disease with type 2 diabetes mellitus. Int. J. Pharm. Sci. 2017, 8, 1420. [Google Scholar]
- Sethi, S.K.; Wong, M.S. The Clinical Biochemistry of Diabetes Mellitus in Singapore. EJIFCC. 2002, 13, 125–128. [Google Scholar]
- Mohamed, J.; Nazratun Nafizah, A.H.; Zariyantey, A.H.; Budin, S.B. Mechanisms of Diabetes-Induced Liver Damage: The role of oxidative stress and inflammation. Sultan Qaboos Univ. Med. J. 2016, 16, e132–e141. [Google Scholar] [CrossRef] [PubMed]
- Balaha, M.; Kandeel, S.; Kabel, A. Phloretin either alone or in combination with duloxetine alleviates the STZ-induced diabetic neuropathy in rats. Biomed. Pharmacother. 2018, 101, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Patel, U.; Kulkarni, M.; Undale, V.; Bhosale, A. Evaluation of diuretic activity of aqueous and methanol extracts of Lepidium sativum garden cress (Cruciferae) in rats. Trop. J. Pharm. Res. 2009, 8, 215–219. [Google Scholar] [CrossRef]
- Baig, M.A.; Gawali, V.B.; Patil, R.R.; Naik, S.R. Protective effect of herbomineral formulation (Dolabi) on early diabetic nephropathy in streptozotocin-induced diabetic rats. J. Nat. Med. 2012, 66, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Necker, R.; Hellon, R.F. Noxious thermal input from the rat tail: Modulation by descending inhibitory influences. Pain 1978, 4, 231–242. [Google Scholar] [CrossRef]
- Anjaneyulu, M.; Chopra, K. Quercetin attenuates thermal hyperalgesia and cold allodynia in STZ-induced diabetic rats. Indian J. Exp. Biol. 2004, 42, 766–769. [Google Scholar] [PubMed]
- Attal, N.; Jazat, F.; Kayser, V.; Guilbaud, G. Further evidence for ‘pain-related’ behaviours in a model of unilateral peripheral mononeuropathy. Pain 1990, 41, 235–251. [Google Scholar] [CrossRef]
- Goyal, B.R.; Mesariya, P.; Goyal, R.K.; Mehta, A.A. Effect of telmisartan on cardiovascular complications associated with streptozotocin diabetic rats. Mol. Cell Biochem. 2008, 314, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Goyal, B.R.; Parmar, K.; Goyal, R.K.; Mehta, A.A. Beneficial role of telmisartan on cardiovascular complications associated with STZ-induced type 2 diabetes in rats. Pharmacol. Rep. 2011, 63, 956–966. [Google Scholar] [CrossRef]
- Hon, J.K.; Steendijk, P.; Petrou, M.; Wong, K.; Yacoub, M.H. Influence of clenbuterol treatment during six weeks of chronic right ventricular pressure overload as studied with pressure-volume analysis. J. Thorac. Cardiovasc. Surg. 2001, 122, 767–774. [Google Scholar] [CrossRef] [Green Version]
- Qiu, W.C.; Wang, Z.G.; Lv, R.; Wang, W.G.; Han, X.D.; Yan, J.; Wang, Y.; Zheng, Q.; Ai, K.X. Ghrelin improves delayed gastrointestinal transit in alloxan-induced diabetic mice. World J. Gastroenterol. 2008, 14, 2572–2577. [Google Scholar] [CrossRef]
- Zheng, Q.; Qiu, W.C.; Yan, J.; Wang, W.G.; Yu, S.; Wang, Z.G.; Ai, K.X. Prokinetic effects of a ghrelin receptor agonist GHRP-6 in diabetic mice. World J. Gastroenterol. 2008, 14, 4795–4799. [Google Scholar] [CrossRef]
- Janseen, P.A.S.; Jagenerous, A.H. New series of potent analysis. J. Pharm. Pharmacol. 1957, 6, 38–40. [Google Scholar]
- Peddireddy, M.K.R. In vivo methods for evaluation of drugs for the treatment of gastrointestinal motility disorders. Indian J. Pharm. Educ. Res. 2010, 44, 42–48. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Acta 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Griess, J.P.; Bemerkungen, Z.A.H.H. Ueber einige azoverbindungen. Ber. Deutch Chem. Ges. 1879, 12, 426–428. [Google Scholar] [CrossRef] [Green Version]
- Svoboda, P.; Mosinger, B. Catecholamines and the brain microsomal Na, K-adenosinetriphosphatase-I. Protection against lipoperoxidative damage. Biochem Pharmacol. 1981, 30, 427–432. [Google Scholar] [CrossRef]
- Rao, B.S.; Deshpande, V. Experimental Biochemistry-A Student Companion; I. K. International Pvt. Ltd.: New Delhi, India, 2005; pp. 28–31. [Google Scholar]
- Babu, C.S.; Ramanathan, M. Post-ischemic administration of nimodipine following focal cerebral ischemic-reperfusion injury in rat alleviated excitotoxicity, neurobehavioural alterations and partially the bioenergetics. Int. J. Dev. Neurosci. 2011, 29, 93–105. [Google Scholar] [CrossRef]
- Cancelas, J.; Sancho, V.; Villanueva-Peñacarrillo, M.L.; Courtois, P.; Scott, F.W.; Valverde, I.; Malaisse, W.J. Glucagon-like peptide 1 content of intestinal tract in adult rats injected with streptozotocin either during neonatal period or 7 d before sacrifice. Endocrine 2002, 19, 279–286. [Google Scholar] [CrossRef]
- Hinnen, D. Glucagon-Like Peptide 1 Receptor Agonists for Type 2 Diabetes. Diabetes Spectr. 2017, 30, 202–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, R.A.; Rauscher, F.M.; Watkins, J.B., 3rd. Effects of quercetin on antioxidant defense in streptozotocin-induced diabetic rats. J. Biochem Mol. Toxicol. 2001, 15, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Abdollah, M.; Zuki, A.B.Z.; Goh, Y.; Rezaeizadeh Rezaeizadeh, A.; Noordin, M.M. The effects of Momordica charantia on the liver in streptozotocin-induced diabetes in neonatal rats. Afr. J. Biotechnol. 2010, 9, 5004–5012. [Google Scholar]
- Shinde, U.A.; Goyal, R.K. Effect of chromium picolinate on histopathological alterations in STZ and neonatal STZ diabetic rats. J. Cell Mol. Med. 2003, 7, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Hamadi, N.; Mansour, A.; Hassan, M.H.; Khalifi-Touhami, F.; Badary, O. Ameliorative effects of resveratrol on liver injury in streptozotocin-induced diabetic rats. J. Biochem Mol. Toxicol. 2012, 26, 384–392. [Google Scholar] [CrossRef]
- Chang, A.S.; Hathaway, C.K.; Smithies, O.; Kakoki, M. Transforming growth factor-β1 and diabetic nephropathy. Am. J. Physiol. Renal Physiol. 2016, 310, F689–F696. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, V.; Snehalatha, C.; Kumutha, R.; Jayaraman, M.; Ramachandran, A. Serum albumin levels in different stages of type 2 diabetic nephropathy patients. Indian J. Nephrol. 2004, 14, 89–92. [Google Scholar]
- Zhu, Y.; Cai, X.; Liu, Y.; Hu, M.; Zhou, L.; Liu, W.; Wu, J.; Zhang, R.; Gao, X.; Yang, W.; et al. Serum Albumin, but not Bilirubin, is Associated with Diabetic Chronic Vascular Complications in a Chinese Type 2 Diabetic Population. Sci. Rep. 2019, 9, 12086. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, E.N.; Reddy, P.U.; Singh, K.P. Effect of alcoholic extracts of Indian medicinal plants on the altered enzymatic activities of diabetic rats. Indian J. Pharm. Sci. 2009, 71, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.S.; Kim, K.W.; Chung, H.Y.; Yoon, S.; Moon, J.O. Effect of sinapic acid against dimethylnitrosamine-induced hepatic fibrosis in rats. Arch. Pharm Res. 2013, 36, 608–618. [Google Scholar] [CrossRef]
- Dobretsov, M.; Romanovsky, D.; Stimers, J.R. Early diabetic neuropathy: Triggers and mechanisms. World J. Gastroenterol. 2007, 13, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Barragán-Iglesias, P.; Oidor-Chan, V.H.; Loeza-Alcocer, E.; Pineda-Farias, J.B.; Velazquez-Lagunas, I.; Salinas-Abarca, A.B.; Hong, E.; Sánchez-Mendoza, A.; Delgado-Lezama, R.; Price, T.J.; et al. Evaluation of the neonatal streptozotocin model of diabetes in rats: Evidence for a model of neuropathic pain. Pharmacol. Rep. 2018, 70, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Al-Ani, F.S.; Al-Nimer, M.S.; Ali, F.S. Dyslipidemia as a contributory factor in etiopathogenesis of diabetic neuropathy. Indian J. Endocrinol. Metab. 2011, 15, 110–114. [Google Scholar]
- Punkkinen, J.; Färkkilä, M.; Mätzke, S.; Korppi-Tommola, T.; Sane, T.; Piirilä, P.; Koskenpato, J. Upper abdominal symptoms in patients with Type 1 diabetes: Unrelated to impairment in gastric emptying caused by autonomic neuropathy. Diabet. Med. 2008, 25, 570–577. [Google Scholar] [CrossRef]
- Triantafyllou, K.; Kalantzis, C.; Papadopoulos, A.A.; Apostolopoulos, P.; Rokkas, T.; Kalantzis, N.; Ladas, S.D. Video-capsule endoscopy gastric and small bowel transit time and completeness of the examination in patients with diabetes mellitus. Dig. Liver Dis. 2007, 39, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Sung, T.S.; La, J.H.; Kang, T.M.; Kim, T.W.; Yang, I.S. Visceral Hypersensitivity and Altered Colonic Motility in Type 2 Diabetic Rat. J. Neurogastroenterol. Motil. 2015, 21, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Hauschildt, A.T.; Corá, L.A.; Volpato, G.T.; Sinzato, Y.K.; Damasceno, D.C.; Américo, M.F. Mild diabetes: Long-term effects on gastric motility evaluated in rats. Int. J. Exp. Pathol. 2018, 99, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.K.; Wang, L.X.; Guo, L.S.; Luo, P.; Du, J.J.; Zhao, Z.L.; Wang, G.G. Syringic acid inhibits apoptosis pathways via downregulation of p38MAPK and JNK signaling pathways in H9c2 cardiomyocytes following hypoxia/reoxygenation injury. Mol. Med. Rep. 2017, 16, 2290–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirwaikar, A.; Rajendran, K.; Dinesh Kumar, C.; Bodla, R. Antidiabetic activity of aqueous leaf extract of Annona squamosa in streptozotocin-nicotinamide type 2 diabetic rats. J. Ethnopharmacol. 2004, 91, 171–175. [Google Scholar] [CrossRef]
- Kawanami, D.; Matoba, K.; Utsunomiya, K. Dyslipidemia in diabetic nephropathy. Ren. Replace. Ther. 2016, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kameswararao, B.; Kesavulu, M.M.; Apparao, C. Evaluation of antidiabetic effect of Momordica cymbalaria fruit in alloxan-diabetic rats. Fitoterapia 2003, 74, 7–13. [Google Scholar] [CrossRef]
- Al-Rasheed, N.M.; Al-Rasheed, N.M.; Hasan, I.H.; Al-Amin, M.A.; Al-Ajmi, H.N.; Mohamad, R.A.; Mahmoud, A.M. Simvastatin Ameliorates Diabetic Cardiomyopathy by Attenuating Oxidative Stress and Inflammation in Rats. Oxid. Med. Cell Longev. 2017, 2017, 1092015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edet, E.E.; Eno, M.A.A.; Umoh, I.B.; Itam, E.H. Effect of Gongronema latifolium crude leaf extract on some cardiac enzymes of alloxan-induced diabetic rats. Afr. J. Biochem. Res. 2009, 3, 366–369. [Google Scholar]
- Bueno-Orovio, A.; Sánchez, C.; Pueyo, E.; Rodriguez, B. Na/K pump regulation of cardiac repolarization: Insights from a systems biology approach. Pflugers Arch. 2014, 466, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Burwen, S.J.; Schmucker, D.L.; Jones, A.L. Subcellular and molecular mechanisms of bile secretion. Int. Rev. Cytol. 1992, 135, 269–313. [Google Scholar]
- Barman, S.; Pradeep, S.R.; Srinivasan, K. Zinc supplementation alleviates the progression of diabetic nephropathy by inhibiting the overexpression of oxidative-stress-mediated molecular markers in streptozotocin-induced experimental rats. J. Nutr. Biochem. 2018, 54, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Zhou, Q.Y.; Liu, D.; Yu, L.; Zhan, L.; Li, X.J.; Peng, H.Y.; Zhang, X.L.; Yuan, X.C. Advanced glycation end-products impair Na⁺/K⁺-ATPase activity in diabetic cardiomyopathy: Role of the adenosine monophosphate-activated protein kinase/sirtuin 1 pathway. Clin. Exp. Pharmacol Physiol. 2014, 41, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Babu, P.S.; Srinivasan, K. Renal lesions in streptozotocin-induced diabetic rats maintained on onion and capsaicin containing diets. J. Nutr. Biochem. 1999, 10, 477–483. [Google Scholar] [CrossRef]
- Siddiqui, M.R.; Moorthy, K.; Taha, A.; Hussain, M.E.; Baquer, N.Z. Low doses of vanadate and Trigonella synergistically regulate Na+/K+ -ATPase activity and GLUT4 translocation in alloxan-diabetic rats. Mol. Cell Biochem. 2006, 285, 17–27. [Google Scholar] [CrossRef]
- Al-Numair, K.S.; Veeramani, C.; Alsaif, M.A.; Chandramohan, G. Influence of kaempferol, a flavonoid compound, on membrane-bound ATPases in streptozotocin-induced diabetic rats. Pharm. Biol. 2015, 53, 1372–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Tatsunami, R.; Yama, K.; Tampo, Y. Glycolaldehyde induces cytotoxicity and increases glutathione and multidrug-resistance-associated protein levels in Schwann cells. Biol. Pharm Bull. 2013, 36, 1111–1117. [Google Scholar] [CrossRef] [Green Version]
- Bell, D.S. Diabetic cardiomyopathy. Diabetes Care. 2003, 26, 2949–2951. [Google Scholar] [CrossRef] [Green Version]
- Hyogo, H.; Yamagishi, S. Advanced glycation end products (AGEs) and their involvement in liver disease. Curr. Pharm Des. 2008, 14, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Arai, K.; Maguchi, S.; Fujii, S.; Ishibashi, H.; Oikawa, K.; Taniguchi, N. Glycation and inactivation of human Cu-Zn-superoxide dismutase. Identification of the in vitro glycated sites. J. Biol. Chem. 1987, 262, 16969–16972. [Google Scholar] [CrossRef]
- Santini, S.A.; Cotroneo, P.; Marra, G.; Manto, A.; Giardina, B.; Mordente, A.; Greco, A.V.; Martorana, G.E.; Magnani, P.; Ghirlanda, G. NA+/K+ ATPase impairment and experimental glycation: The role of glucose autoxidation. Free Radic. Res. 1996, 24, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Bandeira Sde, M.; Guedes Gda, S.; da Fonseca, L.J.; Pires, A.S.; Gelain, D.P.; Moreira, J.C.; Rabelo, L.A.; Vasconcelos, S.M.; Goulart, M.O. Characterization of blood oxidative stress in type 2 diabetes mellitus patients: Increase in lipid peroxidation and SOD activity. Oxid. Med. Cell Longev. 2012, 2012, 819310. [Google Scholar] [CrossRef] [Green Version]
- Sancak, E.B.; Akbas, A.; Silan, C.; Cakir, D.U.; Turkon, H.; Ozkanli, S.S. Protective effect of syringic acid on kidney ischemia-reperfusion injury. Ren. Fail. 2016, 38, 629–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didangelos, T.; Karlafti, E.; Kotzakioulafi, E.; Kontoninas, Z.; Margaritidis, C.; Giannoulaki, P.; Kantartzis, K. Efficacy and Safety of the Combination of Superoxide Dismutase, Alpha Lipoic Acid, Vitamin B12, and Carnitine for 12 Months in Patients with Diabetic Neuropathy. Nutrients 2020, 12, 3254. [Google Scholar] [CrossRef] [PubMed]
- Coudriet, G.M.; Delmastro-Greenwood, M.M.; Previte, D.M.; Marré, M.L.; O’Connor, E.C.; Novak, E.A.; Vincent, G.; Mollen, K.P.; Lee, S.; Dong, H.H.; et al. Treatment with a Catalytic Superoxide Dismutase (SOD) Mimetic Improves Liver Steatosis, Insulin Sensitivity, and Inflammation in Obesity-Induced Type 2 Diabetes. Antioxidants 2017, 6, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowluru, R.A.; Atasi, L.; Ho, Y.S. Role of mitochondrial superoxide dismutase in the development of diabetic retinopathy. Invest. Ophthalmol Vis. Sci. 2006, 47, 1594–1599. [Google Scholar] [CrossRef] [Green Version]
- Dinçer, Y.; Akçay, T.; Alademir, Z.; Ilkova, H. Assessment of DNA base oxidation and glutathione level in patients with type 2 diabetes. Mutat. Res. 2002, 505, 75–81. [Google Scholar] [CrossRef]
- Aydin, A.; Orhan, H.; Sayal, A.; Ozata, M.; Sahin, G.; Işimer, A. Oxidative stress and nitric oxide related parameters in type II diabetes mellitus: Effects of glycemic control. Clin. Biochem. 2001, 34, 65–70. [Google Scholar] [CrossRef]
- Stevens, R.B.; Sutherland, D.E.; Ansite, J.D.; Saxena, M.; Rossini, T.J.; Levay-Young, B.K.; Hering, B.J.; Mills, C.D. Insulin down-regulates the inducible nitric oxide synthase pathway: Nitric oxide as cause and effect of diabetes? J. Immunol. 1997, 159, 5329–5335. [Google Scholar] [PubMed]
- Ii, M.; Nishimura, H.; Kusano, K.F.; Qin, G.; Yoon, Y.S.; Wecker, A.; Asahara, T.; Losordo, D.W. Neuronal nitric oxide synthase mediates statin-induced restoration of vasa nervorum and reversal of diabetic neuropathy. Circulation 2005, 112, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakar, S.S. Role of nitric oxide in diabetic nephropathy. Semin. Nephrol. 2004, 24, 333–344. [Google Scholar] [CrossRef]
- Wold, L.E.; Ceylan-Isik, A.F.; Ren, J. Oxidative stress and stress signaling: Menace of diabetic cardiomyopathy. Acta Pharmacol. Sin. 2005, 26, 908–917. [Google Scholar] [CrossRef]
- Madar, Z.; Kalet-Litman, S.; Stark, A.H. Inducible nitric oxide synthase activity and expression in liver and hepatocytes of diabetic rats. Pharmacology 2005, 73, 106–112. [Google Scholar] [CrossRef]
- Wright, K.L.; Ward, S.G. Interactions between phosphatidylinositol 3-kinase and nitric oxide: Explaining the paradox. Mol. Cell Biol. Res. Commun. 2000, 4, 137–143. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, M.; Li, J.; Chen, K.; Xia, L.; Wang, W.; Ren, P.; Huang, X. Ex vivo to in vivo extrapolation of syringic acid and ferulic acid as grape juice proxies for endothelium-dependent vasodilation: Redefining vasoprotective resveratrol of the French paradox. Food Chem. 2021, 363, 130323. [Google Scholar] [CrossRef]
- Güzelad, Ö.; Özkan, A.; Parlak, H.; Sinen, O.; Afşar, E.; Öğüt, E.; Yıldırım, F.B.; Bülbül, M.; Ağar, A.; Aslan, M. Protective mechanism of Syringic acid in an experimental model of Parkinson’s disease. Metab. Brain Dis. 2021, 36, 1003–1014. [Google Scholar] [CrossRef]
- Rutter, M.K.; Meigs, J.B.; Sullivan, L.M.; D’Agostino RBSr Wilson, P.W. C-reactive protein, the metabolic syndrome, and prediction of cardiovascular events in the Framingham Offspring Study. Circulation 2004, 110, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.K.; Srinivasan, B.P. Triple verses glimepiride plus metformin therapy on cardiovascular risk biomarkers and diabetic cardiomyopathy in insulin resistance type 2 diabetes mellitus rats. Eur. J. Pharm Sci. 2009, 38, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Frati, G.; Schirone, L.; Chimenti, I.; Yee, D.; Biondi-Zoccai, G.; Volpe, M.; Sciarretta, S. An overview of the inflammatory signalling mechanisms in the myocardium underlying the development of diabetic cardiomyopathy. Cardiovasc. Res. 2017, 113, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Zafar, M.; Naqvi, S.N.U.H. Effects of STZ-Induced diabetes on the relative weights of kidney, liver and pancreas in albino rats: A comparative study. J. Morphol. 2010, 28, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Salahshoor, M.R.; Mohammadi, M.M.; Roshankhah, S.; Najari, N.; Jalili, C. Effect of Falcaria vulgaris on oxidative damage of liver in diabetic rats. J. Diabetes Metab. Disord. 2019, 18, 15–23. [Google Scholar] [CrossRef]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.; Matsumura, T.; Kaneda, Y.; Yorek, M.A.; Beebe, D.; Oates, P.J.; Hammes, H.P.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 2000, 404, 787–790. [Google Scholar] [CrossRef]
- Alaofi, A.L. Sinapic Acid Ameliorates the Progression of Streptozotocin (STZ)-Induced Diabetic Nephropathy in Rats via NRF2/HO-1 Mediated Pathways. Front Pharmacol. 2020, 11, 1119. [Google Scholar] [CrossRef]
- Chu, J.; Yan, R.; Wang, S.; Li, G.; Kang, X.; Hu, Y.; Lin, M.; Shan, W.; Zhao, Y.; Wang, Z.; et al. Sinapic Acid Reduces Oxidative Stress and Pyroptosis via Inhibition of BRD4 in Alcoholic Liver Disease. Front Pharmacol. 2021, 12, 668708. [Google Scholar] [CrossRef]
- Siquet, C.; Paiva-Martins, F.; Lima, J.L.; Reis, S.; Borges, F. Antioxidant profile of dihydroxy- and trihydroxyphenolic acids--a structure-activity relationship study. Free Radic. Res. 2006, 40, 433–442. [Google Scholar] [CrossRef]
- Karamac, M.; Kosiñska, A.; Pegg, R.B. Comparison of radical-scavenging activities for selected phenolic acids. Polish J. Food Nutr. Sci. 2005, 14, 165–170. [Google Scholar]
- Wright, J.S.; Johnson, E.R.; DiLabio, G.A. Predicting the activity of phenolic antioxidants: Theoretical method, analysis of substituent effects, and application to major families of antioxidants. J. Am. Chem Soc. 2001, 123, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, J.; Ma, L.; Li, J.; Shahzad, N.; Kim, C.K. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci. Rep. 2020, 10, 2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, J.; Li, X.; Tian, H.; Liu, C.; Guo, Y.; Zhang, S.; Chu, Y.; Li, J.; Wang, Y.; Zhang, L. Neuroprotective effect of diosgenin in a mouse model of diabetic peripheral neuropathy involves the Nrf2/HO-1 pathway. BMC Complement Med. Ther. 2020, 20, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Eisa, R.A.; Al-Salmi, F.A.; Hamza, R.Z.; El-Shenawy, N.S. Role of L-carnitine in protection against the cardiac oxidative stress induced by aspartame in Wistar albino rats. PLoS ONE 2018, 13, e0204913. [Google Scholar] [CrossRef] [PubMed]
- Rashedinia, M.; Khoshnoud, M.J.; Fahlyan, B.K.; Hashemi, S.S.; Alimohammadi, M.; Sabahi, Z. Syringic Acid: A Potential Natural Compound for the Management of Renal Oxidative Stress and Mitochondrial Biogenesis in Diabetic Rats. Curr. Drug Discov. Technol. 2021, 18, 405–413. [Google Scholar] [CrossRef] [PubMed]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight (gm) | Food Intake (gm) | Water Intake (mL) | Urine Output (mL) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 8th Week | 18th Week | % Increase in BW | 8th Week | FI per 100 g b.w | 18th Week | FI per 100 g b.w | 8th Week | WI per 100 g b.w | 18th Week | WI per 100 g b.w | 8th Week | UOP per 100 g b.w | 18th Week | UOP per 100 g b.w | |
| NC | 198.43 ± 12.15 | 271.17 ± 2.28 | 36.86 ± 6.25 | 9.66 ± 1.10 | 4.86 ± 0.30 | 14.83 ± 1.05 | 5.47 ± 0.29 | 19.57 ± 2.48 | 9.88 ± 1.29 | 29.98 ± 3.29 | 11.06 ± 1.19 | 2.14 ± 0.53 | 1.09 ± 0.31 | 3.12 ± 1.00 | 1.15 ± 0.37 |
| NC + SA | 187.96 ± 9.65 | 267.97 ± 10.0 | 42.86 ± 8.84 | 9.77 ± 0.71 | 5.21 ± 0.54 | 14.96 ± 1.40 | 5.59 ± 0.64 | 19.91 ± 1.72 | 10.66 ± 1.45 | 30.76 ± 3.20 | 11.50 ± 1.40 | 2.35 ± 0.54 | 1.25 ± 0.26 | 3.53 ± 0.98 | 1.31 ± 0.35 |
| DC | 186.56 ± 15.66 | 212.7 ± 17.96 C | 13.07 ± 3.64 C | 12.98 ± 1.09 C | 6.13 ± 0.66 B | 18.76 ± 1.08 C | 8.88 ± 1.09 C | 24.97 ± 4.59 B | 13.31 ± 1.47 C | 36.56 ± 4.33 B | 17.18 ± 1.34 C | 4.85 ± 1.145 C | 2.57 ± 0.44 C | 7.34 ± 1.34 C | 3.43 ± 0.43 C |
| DC + SA25 | 200.84 ± 8.94 | 253.51 ± 10.96 d | 26.43 ± 7.65 a | 14.97 ± 0.89 | 5.90 ± 0.27 B | 17.86 ± 1.29 | 7.05 ± 0.57 Ad | 24.79 ± 1.77 A | 12.36 ± 1.07 A | 35.47 ± 2.91 A | 14.04 ± 1.64 Bb | 4.96 ± 0.67 C | 2.46 ± 0.22 C | 6.26 ± 0.81 | 2.47 ± 0.35 Cd |
| DC + SA50 | 195.39 ± 14.40 | 252.52 ± 12.4 d | 29.46 ± 4.98 b | 15.04 ± 0.85 | 5.96 ± 0.27 B | 16.95 ± 1.60 | 6.70 ± 0.52 Ad | 24.02 ± 1.47 A | 12.33 ± 1.03 A | 32.92 ± 2.37 | 13.03 ± 0.69 d | 4.78 ± 0.96 C | 2.47 ± 0.59 C | 5.04 ± 0.78 | 1.99 ± 0.32 Bd |
| DC + MET | 189.77 ± 12.05 | 252.22 ± 9.41 d | 33.38 ± 9.98 d | 11.04 ± 1.27 | 5.82 ± 0.66 | 16.63 ± 1.57 | 6.59 ± 0.51 Ad | 23.41 ± 1.89 | 12.35 ± 0.82 A | 32.60 ± 1.78 | 12.92 ± 0.42 d | 4.66 ± 0.69 C | 2.45 ± 0.29 C | 4.80 ± 0.90 | 1.91 ± 0.39 Bd |
| Parameters | No. of Weeks | NC | NC + SA | DC | DC + SA25 | DC + SA50 | DC + MET |
|---|---|---|---|---|---|---|---|
| FBG (mg/dL) | 8 | 95.39 ± 3.56 | 92.75 ± 4.64 | 156.38 ± 4.40 C | 157.46 ± 5.38 C | 155.97 ± 7.76 C | 156.69 ± 6.67 C |
| 13 | 91.54 ± 4.58 | 87.99 ± 5.29 | 204.32 ± 7.65 C | 183.59 ± 11.66 Cb2 | 175.52 ± 6.53 Cd | 163.79 ± 8.41 Cd | |
| 18 | 91.02 ± 6.42 | 94.33 ± 4.85 | 247.86 ± 15.0 C | 218.96 ± 12.21 Cb3 | 185.91 ± 10.34 Cd1 | 165.00 ± 11.49 Cd | |
| BUN (mg/dL) | 8 | 18.87 ± 3.54 | 19.83 ± 4.64 | 28.27 ± 4.41 B | 26.77 ± 5.58 A | 23.37 ± 4.29 | 25.68 ± 4.46 |
| 13 | 20.04 ± 3.57 | 21.87 ± 3.91 | 38.75 ± 6.11 C | 37.13 ± 6.05 C | 28.58 ± 3.71 a | 29.13 ± 4.57 a | |
| 18 | 19.03 ± 2.26 | 22.34 ± 4.70 | 50.04 ± 5.48 C | 40.67 ± 6.95 Ca3 | 34.69 ± 5.45 Cd | 31.07 ± 5.38 Bd | |
| SCr (mg/dL) | 8 | 0.45 ± 0.050 | 0.43 ± 0.049 | 0.49 ± 0.089 | 0.45 ± 0.090 | 0.48 ± 0.047 | 0.47 ± 0.079 |
| 13 | 0.49 ± 0.052 | 0.47 ± 0.075 | 0.81 ± 0.074 C | 0.54 ± 0.067 d | 0.53 ± 0.061 d | 0.53 ± 0.065 d | |
| 18 | 0.47 ± 0.064 | 0.45 ± 0.40 | 0.97 ±0.072 C | 0.60 ± 0.074 Ad | 0.57 ± 0.053 d | 0.55 ±0.11 d | |
| TC (mg/dL) | 8 | 121.69 ± 6.27 | 113.52 ± 9.26 | 142.19 ± 5.39 C | 138.49 ± 6.43 C | 137.76 ± 7.74 B | 133.32 ± 7.12 A |
| 13 | 115.92 ± 7.66 | 118.43 ± 10.95 | 194.01 ± 7.93 C | 172.59 ± 5.82 Cd3 | 152.50 ± 5.45 Cd | 139.46 ± 6.95 Cd | |
| 18 | 117.41 ± 10.10 | 122.55 ± 8.45 | 259.83 ± 16.59 C | 213.14 ± 18.33 Cd3 | 176.71 ± 15.18 Cd1 | 148.67 ± 13.82 Ad | |
| TG (mg/dL) | 8 | 79.84 ± 5.58 | 83.12 ± 4.86 | 92.2 ± 7.13 A | 95.03 ± 6.30 B | 88.28 ± 6.96 | 87.60 ± 7.79 |
| 13 | 76.8 ± 6.64 | 82.01 ± 8.44 | 119.39 ± 10.31 C | 108.16 ± 5.49 C2 | 93.75 ± 5.40 Bd | 91.79 ± 8.69 Ad | |
| 18 | 78.22 ± 5.41 | 81.54 ± 6.31 | 154.03 ± 6.07 C | 129.63 ± 11.35 Cb3 | 98.46 ± 10.09 Ad | 92.47 ± 7.84 d | |
| AST (U/L) | 8 | 97.63 ± 8.99 | 103.18 ± 5.21 | 114.66 ± 7.72 A | 117.45 ± 7.87 B | 116.47 ± 9.57 B | 113.13 ± 6.59 A |
| 13 | 103.64 ± 4.55 | 105.93 ± 7.01 | 147 ± 12.85 C | 141 ± 8.29 C3 | 124.78 ± 11.60 Bb | 117.32 ± 5.87 d | |
| 18 | 104.36 ± 7.54 | 107.79 ± 11.38 | 185.53 ± 13.39 C | 160.63 ± 17.56 Ca3 | 138.49 ± 10.13 Bd | 121.17 ± 12.85 d | |
| ALT (U/L) | 8 | 39.81 ± 5.20 | 37.93 ± 4.18 | 53.12 ± 5.96 B | 52.36 ± 3.89 B | 54.56 ± 5.16 B | 53.4 ± 6.25 B |
| 13 | 40.88 ± 4.71 | 39.66 ± 4.24 | 59.71 ± 3.72 C | 55.85 ± 5.66 C | 52.63 ± 6.25 A | 49.51 ± 5.93 a | |
| 18 | 41.23 ± 3.16 | 42.57 ± 4.47 | 70.93 ± 5.11 C | 61.76 ± 6.35 Ca3 | 51.28 ± 5.28 Ad | 45.66 ± 4.36 d |
| Groups | UCr (mg/dL) | CrCl (ml/min) | UALB (mg/L) | Na (mEq/L) | K (mEq/L) |
|---|---|---|---|---|---|
| NC | 38.26 ± 2.09 | 270.46 ± 57.03 | 20.37 ± 8.37 | 104.70 ± 16.75 | 22.50 ± 3.54 |
| NC + SA | 36.12 ± 5.16 | 285.70 ± 77.95 | 18.51 ± 5.73 | 105.28 ± 7.59 | 20.31 ± 4.32 |
| DC | 16.35 ± 2.30 C | 124.30 ± 26.30 C | 127.22 ± 33.13 C | 156.61 ± 13.82 C | 8.26 ± 2.88 C |
| DC + SA 25 | 23.66 ± 2.88 Ca | 217.01 ± 30.17 a | 86.11 ± 17.84 Cb2 | 126.67 ± 12.33 b | 14.32 ± 2.62 Ba |
| DC + SA50 | 29.50 ± 4.85 Bd | 242.83 ± 40.65 b | 56.85 ± 14.54 Bd | 114.90 ± 10.96 d | 17.13 ± 2.48 Ad |
| DC + MET | 30.80 ± 3.15 Ad | 254.05 ± 27.75 d | 45.18 ± 8.65 d | 108.47 ± 11.15 d | 18.02 ± 2.26 d |
| Parameters | NC | NC + SA | DC | DC + SA25 | DC + SA50 | DC + MET |
|---|---|---|---|---|---|---|
| HbA1C (%) | 4.32 ± 0.70 | 4.52 ± 0.81 | 7.881 ± 0.60 C | 6.38 ± 1.02 Ba | 5.89 ± 0.64 Ab | 5.17 ± 0.78 d |
| TG (mg/dL) | 85.32 ± 6.43 | 89.10 ± 7.00 | 259.83 ± 16.59 C | 213.14 ± 18.33 Cd3 | 176.71 ± 15.18 Cd1 | 148.67 ± 13.82 Ad |
| TC (mg/dL) | 117.41 ± 10.10 | 122.55 ± 8.45 | 154.03 ± 6.81 C | 129.68 ± 11.80 Cb3 | 98.46 ± 9.58 Ad | 92.47 ± 7.40 d |
| HDL (mg/dL) | 50.33± 6.03 | 54.16 ± 4.52 | 32.75± 3.46 C | 42.50± 6.16 a | 44.95 ± 4.90 b | 46.16 ± 4.49 b |
| LDL (mg/dL) | 49.99± 12.24 | 50.57± 10.30 | 196.26 ± 16.27 C | 144.72± 8.80 Cd3 | 112.09 ± 16.53 Cd | 83.96± 16.60 Ad |
| AI | 1.36 ± 0.35 | 1.27 ± 0.27 | 6.98 ± 0.77 | 4.10 ± 0.85 Ab | 2.97 ± 0.58 d | 2.26 ± 0.59 d |
| TP (g/dL) | 8.56 ± 0.71 | 9.20 ± 0.64 | 5.55 ± 0.42 C | 6.85 ± 0.43 Cb1 | 7.38 ± 0.43 Ad | 7.89 ± 0.52 d |
| SCr (mg/dL) | 0.47 ± 0.06 | 0.45 ± 0.07 | 0.97 ± 0.19 C | 0.68 ± 0.11 Bd | 0.61 ± 0.08 d | 0.57± 0.07 d |
| BUN (mg/dL) | 19.03 ± 2.19 | 22.34 ± 3.46 | 50.04 ± 5.64 C | 37.67 ± 4.47 Cb1 | 32.67 ± 5.64 Cd | 27.56 ± 4.68 Ad |
| ALB (g/dL) | 3.72 ± 0.41 | 3.84 ± 0.61 | 2.66 ± 0.45 C | 3.17 ± 0.12 Aa1 | 3.51 ± 0.31 d | 3.67 ± 0.12 d |
| CRP (mg/l) | 0.60 ± 0.65 | 0.8 ± 0.61 | 3.60 ± 1.31 C | 2.20 ± 0.48 Aa | 1.80 ± 0.65 d | 1.6 ± 0.61 d |
| LDH (U/l) | 339.53 ± 37.69 | 345.89 ± 45.70 | 698.02 ± 56.87 C | 575.84 ± 26.97 Cd3 | 439.68 ± 43.95 Bd | 415.34 ± 36.86 d |
| CK-MB (U/l) | 294.41 ± 25.11 | 301.58 ± 24.73 | 507.21 ± 62.19 C | 438.56 ± 23.76 Ca3 | 346.76 ± 25.72 d | 325.56 ± 37.82 d |
| AST (U/L) | 104.36 ± 7.54 | 107.79 ± 11.38 | 185.53 ± 13.39 C | 160.63 ± 17.22 Ca3 | 138.49 ± 10.13 Cd | 121.17 ± 12.85 d |
| ALT (U/L) | 41.23 ± 3.16 | 42.57 ± 4.47 | 70.93 ± 5.11 C | 61.76 ± 6.35 Ca3 | 51.28 ± 5.28 Ad | 45.66± 4.36 d |
| BR (mg/dL) | 0.25 ± 0.04 | 0.24 ± 0.07 | 0.46 ± 0.04 C | 0.37 ± 0.04 Ca2 | 0.32 ± 0.06 d | 0.26 ± 0.043 d |
| Group | % GE | % SIT |
|---|---|---|
| NC | 53.54 + 8.89 | 63.12 + 5.37 |
| NC+SA | 49.10 + 4.10 | 64.78 + 4.38 |
| DC | 25.89 + 8.98 C | 36.89 + 2.66 C |
| DC + SA25 | 41.47 + 7.16 a | 48.04 + 5.17 Cb1 |
| DC + SA50 | 45.38 + 4.22 d | 53.77 + 4.87 Ad |
| DC + MET | 48.38 + 6.10 d | 57.78 + 4.77 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirza, A.C.; Panchal, S.S.; Allam, A.A.; Othman, S.I.; Satia, M.; Mandhane, S.N. Syringic Acid Ameliorates Cardiac, Hepatic, Renal and Neuronal Damage Induced by Chronic Hyperglycaemia in Wistar Rats: A Behavioural, Biochemical and Histological Analysis. Molecules 2022, 27, 6722. https://doi.org/10.3390/molecules27196722
Mirza AC, Panchal SS, Allam AA, Othman SI, Satia M, Mandhane SN. Syringic Acid Ameliorates Cardiac, Hepatic, Renal and Neuronal Damage Induced by Chronic Hyperglycaemia in Wistar Rats: A Behavioural, Biochemical and Histological Analysis. Molecules. 2022; 27(19):6722. https://doi.org/10.3390/molecules27196722
Chicago/Turabian StyleMirza, Anwarbaig C., Shital S. Panchal, Ahmed A. Allam, Sarah I. Othman, Milan Satia, and Sanjay N. Mandhane. 2022. "Syringic Acid Ameliorates Cardiac, Hepatic, Renal and Neuronal Damage Induced by Chronic Hyperglycaemia in Wistar Rats: A Behavioural, Biochemical and Histological Analysis" Molecules 27, no. 19: 6722. https://doi.org/10.3390/molecules27196722
APA StyleMirza, A. C., Panchal, S. S., Allam, A. A., Othman, S. I., Satia, M., & Mandhane, S. N. (2022). Syringic Acid Ameliorates Cardiac, Hepatic, Renal and Neuronal Damage Induced by Chronic Hyperglycaemia in Wistar Rats: A Behavioural, Biochemical and Histological Analysis. Molecules, 27(19), 6722. https://doi.org/10.3390/molecules27196722